Aminokwasy
Wiadomości ogólne
Aminokwasy są związkami organicznymi zawierającymi dwie grupy funkcyjne,
karboksylową -COOH i aminową -NH2. Aminokwasy są podstawowymi jednostkami
strukturalnymi białek. Każdy aminokwas prócz grup funkcyjnych zawiera też atom wodoru i
charakterystyczny dla siebie podstawnik oznaczany ogólnie jako R. Jest on różny dla
poszczególnych aminokwasów. Inaczej jest to łańcuch boczny aminokwasu. Ten składnik
aminokwasów ma różne kształty, wielkości, ładunki elektryczne i reaktywność. Łańcuch
boczny może mieć właściwości kwasowe, zasadowe, obojętne, może mieć też charakter
alifatyczny, aromatyczny lub heterocykliczny.
Budowa aminokwasu
Grupa aminowa i karboksylowa aminokwasów jest przyłączona do tego samego węgla a i stąd
nazwa a-aminokwasy. Organizmy żywe budują wyłącznie a-aminokwasy.
Aminokwasy występujące w przyrodzie
W przyrodzie występuje ponad 150 różnych aminokwasów, częstotliwość wykrywania
wzrosła wraz z rozwojem technik chromatograficznych, które pozwalają wykryć nawet
niewielkie ilości aminokwasów w badanej próbce. Systematycznym badaniom poddawane są
próbki pochodzenia roślinnego jak i zwierzęcego, które mogą być źródłami nowych
aminokwasów. Dużą część nowoodkrytych aminokwasów powstaje w roślinach na skutek
przemiany materii podczas kiełkowania. Bardzo rzadko spotykane aminokwasy wykryto
podczas hydrolizy antybiotyków i innych produktów wydzielanych przez mikroorganizmy.
Pierwszym odkrytym aminokwasem występującym w przyrodzie była aspargina,
wyodrębniona z soku Asparagusa w 1906 r. przez Vauquelina i Robiqueta.
Stereoizometria aminokwasów
W 1935 roku W. Kuhn określił absolutną konfigurację 1-metylopropanolu oraz kwasu
mlekowego, na podstawie obliczeń dyspersji skręcalności optycznej. To osiągnięcie
umożliwiło określenie absolutnej konfiguracji aminokwasów pod warunkiem, że można je
było przekształcić w kwas mlekowy. W 1933 roku W. Kuhn i K. Freundberg zaobserwowali
korelację między skręcalnością optyczną i sposobem podstawiania analogicznych
pochodnych alaniny i kwasu mlekowego. Wartość skręcalności molowej wykazuje ten sam
kierunek zmian od dodatnich do ujemnych dla L-(+)-alaniny i kwasu L-(+)-mlekowego
sugerując, że związki te należą do tego samego szeregu. Ostatecznym dowodem, że
aminokwasy występujące w białkach należą do serii L, była praca K. Ingolda z 1950 roku.
Powiązał on bezpośrednio kwas L-(+)-mlekowy z L-(+)-alaniną poprzez kwasy D-(+)bromopropionowy i L-azydopropionowy jako produkty pośrednie.
Treonina, izoleucyna i hydroksyprolina maja drugie centrum chiralności i dlatego mogą
istnieć w czterech formach stereoizometrycznych. Względna konfiguracja atomów Cb w
treoninie jest analogiczna do kwasu L-winowego.
Podział aminokwasów
Tradycyjny podział
Najczęściej stosowanym podziałem aminokwasów jest podział na podstawie charakterystyki
właściwości podstawnika aminokwasów. Wyodrębniamy siedem grup:
Aminokwasy alifatyczne
Ich podstawnikiem jest łańcuch węglowodorowy. Najprostszym przedstawicielem tej grupy
jest glicyna, która za cały łańcuch boczny ma atom wodoru. Alanina ma łańcuch boczny
utworzony z grupy metylowej. Walina, leucyna i izoleucyna mają rozgałęzione łańcuchy
węglowodorowe. Nietypowym aminokwasem jest prolina zawierająca układ heterocykliczny
z drugorzędową resztą aminową. Jej pochodna hydroksyprolina jest przykładem aminokwasu
rzadkiego.
Aminokwasy zawierające grupę hydroksylową
Do grupy tej należy seryna i treonina. Aminokwasy te mają podstawioną grupę hydroksylową
w miejsce wodoru przyłączonego do pierwszego atomu węgla w podstawniku.
Aminokwasy zawierające siarkę w podstawniku
Są to metionina i cysteina. Oba posiadają pewne unikalne cechy, które są wykorzystywane
przez wszystkie organizmy żywe.
Metionina jest aminokwasem startowym w procesie translacji białka.
Cysteina posiada wolna grupę sulfhydrolową, która może reagować z taką samą grupą drugiej
cysteiny. Reakcja ta prowadzi do utworzenia mostka siarczkowego pomiędzy atomami siarki.
Wiązanie to odgrywa ważną role w tworzeniu struktury przestrzennej białek.
Aminokwasy aromatyczne
Aminokwasy aromatyczne posiadają w łańcuchu podstawnika pierścienie sześciowęglowe.
Należą do nich: fenyloalanina, tyrozyna i tryptofan. Obecność pierścienia aromatycznego
powoduje, że fenyloalanina i tryptofan nie są syntezowane przez zwierzęta. Ich pochodne
wykazują właściwości oligodynamiczne i regulatorowe np. hormony T3 i T4.
Zalicza się do nich, asparginę i glutaminę. Ich cząsteczki zawierają zmodyfikowaną grupę
karboksylową przez wstawienie za grupę hydroksylową grupy aminowej.
Aminokwasy kwaśne
2
Wykazują odczyn kwaśny, ponieważ mają dodatkową grupę karboksylową. Zalicza się do
nich: kwas asparginowy i kwas glutaminowy.
Aminokwasy zasadowe
Wykazują odczyn zasadowy, ponieważ mają dodatkową grupę aminową. Zalicza się do nich:
lizynę, argininę i histydynę, przy czym ta ostatnia nie zawsze posiada ładunek dodatni (zależy
to od bezpośredniego otoczenia aminokwasu). Histydyna ma bardzo słabe właściwości
zasadowe, często występuje w centrach aktywnych wielu enzymów związanych z
odwodornieniem, gdyż może łatwo odłączać lub przyłączać protony.
U Procaryota spotyka się rzadkie aminokwasy posiadające dwie grupy aminowe i dwie
karboksylowe. Przykładem jest kwas dwuaminopimelinowy - ważny składnik ściany.
Aminokwasy występujące w białkach
Istnieje kilka sposobów klasyfikacji aminokwasów. Jednym ze sposobu jest podział na
aminokwasy wystękujące w białkach. Białka są budowane przez 20 aminokwasów
występujących w nich. Te same aminokwasy budują białka zwierzęce jak i roślinne.
Aminokwasy budujące białka to: Glicyna, Alanina, Walina, Leucyna, Izoleucyna, Prolina,
Hydroksyprolina, Seryna, Treonina, Cysteina, Metionina, Fenyloalanina, Tyrozyna,
Tryptofan, Aspargina, Glutamina, Kwas asparginowy, Kwas glutaminowy, Lizyna, Arginina i
Histydyna.
Aminokwasy egzogenne i endogenne
Aminokwasy egzogenne są to aminokwasy, które nie są syntezowane w organizmie ludzkim,
a ich obecność w białkach spożywanych decyduje o wartości odżywczej. Histydyna jest
niezbędna dla dzieci do 12 roku życia, ale nie jest niezbędna dla dorosłych.
Aminokwasy endogenne są to aminokwasy, które są syntezowane w organizmie ludzkim.
Aminokwasy glikogenne i ketogenne
Aminokwasy glikogenne są to aminokwasy, których metabolizm prowadzi do powstania
glukozy (sacharydów).
Aminokwasy ketogenne są to aminokwasy, których metabolizm prowadzi do powstania
związków ketonowych (kwas b-hydroksymasłowy, kwas acetomasłowy, aceton).
Rys. 3 Izomery treoniny
Izoleucyna ma konfiguracje analogiczną, a różni się tym od treoniny, że zamiast grupy
hydroksylowej jest rodnik etylowy.
Atom hydroksyproliny ma konfigurację L względem standardu, którym jest aldehyd
glicerynowy.
3
Zaobserwowano korelacje między konfiguracją a smakiem niektórych aminokwasów.
L-tryptofan, L-fenyloalanina, L-tyrozyna i L-leucyna mają smak gorzki, w przeciwieństwie
do słodkich D-związków. Glicyna ma słodki smak a jego sodowa sól jest używana w
przemyśle spożywczym jako przyprawa. Kwas asparginowy nie ma smaku, natomiast jego
dipeptyd z fenyloalaniną ma bardzo słodki smak.
Właściwości
Rozpuszczalność
Z kilkoma wyjątkami aminokwasy są dobrze rozpuszczalne w wodzie, amoniaku i innych
rozpuszczalnikach polarnych, ale są źle rozpuszczalne w niepolarnych i mniej polarnych
rozpuszczalnikach takich jak: etanol, metanol, aceton itp. Spowodowane to jest tym, że w
obojętnych roztworze wodnym występują w postaci jonów obojnaczych.
Na przykład glicyna i alanina stosunek formy nie naładowanych cząsteczek do formy jonu
obojnaczego wynosi 1: 2600 000. Przekształcenie jonu obojnaczego w cząsteczkę nie
naładowaną aby się rozpuściła w mniej polarnym rozpuszczalniku jest bardzo niekorzystne
energetycznie. Rozpuszczalność aminokwasów zależy również od ich łańcuchów bocznych.
Aminokwasy aromatyczne tyrozyna i fenyloalanina mają szczególnie małą rozpuszczalność w
wodzie. Iminokwasy (prolina i hydroksyprolina) mają rozpuszczalność względnie dużą.
Prolina jest również dobrze rozpuszczalna w alkoholu. Prawie wszystkie aminokwasy cechuje
rozpuszczalność w gorącym kwasie octowym, jest to związane z tworzeniem się wiązań
wodorowych pomiędzy kwasem a aminokwasem.
Widma UV i IR aminokwasów
Alifatyczne aminokwasy nie absorbują w nadfiolecie powyżej 220 nm., natomiast takie
aminokwasy aromatyczne jak tyrozyna, arginina, histydyna, fenyloalanina wykazują
maksimum absorpcji dla fali o długości powyżej 250 nm. W widmach IR aminokwasów nie
znajduje się normalnych drgań rozciągających grupy NH, które powinny powodować
absorpcję w obszarze 3300-3500 cm-1.
Pojawia się pasmo absorpcyjne przy 3700 cm-1, Które przypisuje się grupie -NH3+ Pasmo to
się pojawia się w chlowodarkach aminokwasów, ale nie obserwuje się go w N-podstawionych
pochodnych oraz iminokwasach. Obserwuje się też dwa dodatkowe pasma charakterystyczne
dla -NH3+, w obszarze 1500-1600 cm-1. Wszystkie aminokwasy utworzone przez grupy
karboksylowe wykazują absorpcje typowa dla -COO- w zakresie 1500-1600 cm-1, której nie
obserwuje się w przypadku chlorowodorków aminokwasów. W chlorowodorkach
aminokwasów pasmo absorpcji grupy -COOH jest przesunięte w kierunku wyższych wartości
fal o około 0,05 cm ze względu na obecność grupy -NH3+.
Pierwsze widmo dla aminokwasów białkowych zostało pierwszy raz otrzymane przez Takadę
i Jardezkiego w 1957 r. Badacze ci stwierdzili zależność przesunięć chemicznych a
kwasowością środowisk.
Białka
4
Wiadomości ogólne
Ogólna charakterystyka
Białka są to polipeptydy zbudowane z około 100 aminokwasów. Charakteryzują je dość duże
masy cząsteczkowe powyżej 10 000 Da. Pierwiastkami budującymi białka w większej mierze
są: C, O, H, N, S i P. Informacje o budowie białek jest zawarta w DNA. Ogromne bogactwo
form żywych i złożoność budowy należy zawdzięczać białkom. W organizmie żywym
większość związków organicznych stanowią białka, które stanowią 50% suchej masy
komórki. Cechuje je daleko posunięta swoistość gatunkowa, a nieraz osobnicza.
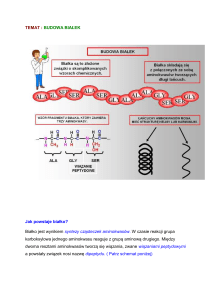
Zasadniczy szkielet części białkowej stanowią powtarzające się w łańcuchu polipeptydowym
elementy strukturalne -CH-CO-NH-, składające się z węgla a oraz wiązania peptydowego CO-NH-. Atom wodoru grupy NH znajduje się prawie zawsze w pozycji trans do atomu tlenu
grupy ketonowej. Łącząc te elementy węglem a rodników aminokwasowych (R) otrzymuje
się łańcuch polipeptydowy o budowie zygzaka.
O takim kształcie łańcucha peptydowego decyduje sposób ułożenia atomów w wiązaniu
peptydowym. Przegrupowanie elektronów (delokalizacja) w obrębie wiązania peptydowego
powoduje, że staje się ono polarne i może występować w dwóch odmianach.
Wiązanie C-N ma około w 40% charakter wiązania podwójnego, a wiązanie C-O w około
60%. Wytworzenie wiązania podwójnego pomiędzy C i N nadaje wiązaniu peptydowemu
znaczna sztywność oraz sprawia, że wszystkie jego cztery atomy (CO i NH) są rozmieszczone
w tej samej płaszczyźnie. Łańcuch ten nie jest do końca sztywny, gdyż pojedyncze wiązanie
przy atomach a węgla umożliwia ruch obrotowy. W tych miejscach łańcuch może się zginać
lub zwijać.
Przegrupowanie atomów (delokalizacja)
Budowę białek bada się metodami chemicznymi, fizykochemicznymi i fizycznymi.
Początkowo białko jest wyizolowywane i oczyszczane. Sekwencje aminokwasów wyznacza
się chemicznie przez znakowanie końcowej grupy amidowej dinitrofluorobenzenem i
następnie przez hydrolizę wg. Sangera, można też oznaczać sekwencje aminokwasów metodą
Edmana, w trakcie, której aminokwas o wolnej grupie aminowej uwalnia się z peptydu w
postaci pochodnej hydantoinowej. Biochemicznie sekwencje aminokwasów wyznacza się
przez hydrolizę enzymatyczną, głównie przy udziale karboksy- i aminopeptydaz.
Dane na temat struktury przestrzennej białek dostarczają badania kryształów białek metodami
dyfrakcji promieni rentgenowskich.
2.1.2. Znaczenie białek
Białka pełnią różne ważne funkcje biologiczne. Stanowią element budulcowy i wzmacniający
(podporowy, osłaniający, spajający), np. keratyna naskórka, kolagen tkanki łącznej, białka
błon komórkowych. Występują też białka kurczliwe np. aktyny i miozyny z miofibryli tkanki
mięśniowej, które łącząc się powodują skurcz mięśnia realizując jego ruch. We wszystkich
komórkach występują białka katalityczne tzw. enzymy, które przyspieszają reakcje
chemiczne. W płynach ustrojowych występuje wiele białek transportujących, które wiążą i
przenoszą różne cząsteczki np. hemoglobina transportuje tlen, a inne transportują jony metali
hormony, składniki pokarmowe. Do białek regulujących należą hormony. Do białek
obronnych należą immunoglobuliny, tworzące różne przeciwciała. Białka sensoryczne
(nerwowe) z narządów zmysłowych np. rodopsyna mają zdolność odbierania i przesyłania
5
sygnałów z środowiska. Włosy i pióra okrywające ciała zwierząt są zbudowane z keratyny. W
tym przypadku białko pełni funkcje izolacyjną od wpływów środowiska zewnętrznego. U
roślin białka pełnią też funkcję magazynującą substancje zapasowe i energetyczne odkładane
w nasionach. Jady i toksyny zwierzęce w dużej mierze oparte są na substancjach białkowych.
W tym wypadku białka pełnia funkcje toksyczne. Funkcje pełnione w organizmie przez białka
są opisane w rozdziale 2.6.
Aspekty historyczne
W 1784 r. De Fourcroy jako pierwszy doszedł do wniosku, że białka stanowią szczególną
klasę substancji. Angielska nazwa białek protein, pochodzi od greckiego słowa proteuo
(zajmuje pierwsze miejsce) i została wprowadzona przez Berzeliusa. Pod koniec XIX w.
Kossel wyodrębnił pewna ilość białek. W tym samym czasie Hofmeister, i Fischer stwierdzili,
że białka są łańcuchami złożonymi z jednostek aminokwasowych, co udowodniono później,
dokonując syntezy tych związków. Hofmeister pracując z albuminą jajka kurzego, jako
pierwszy wyodrębnił białko w postaci krystalicznej. Abel w 1925 r. wykrystalizował insulinę,
a dziesięć lat później Summer przeprowadził krystalizację ureazy. Stanley w 1935 r. otrzymał
w krystalicznej postaci wirusa mozaiki tytoniowej. Svebderg określił masę cząsteczkową
wielu białek metodą ultra wirowania. Rozwój nowych technik analitycznych np. elektroforazy
(Tiselius w 1937 r.) oraz ulepszenie technik chromatograficznych doprowadziło do wielu
odkryć w tej dziedzinie.
W 1956 r. Sanger określił sekwencje aminokwasów w insulinie. Metody wykorzystane w tej
pracy, były podstawa do ustanowienia zasad systematycznego badania pierwszorzędowej
struktury wielu innych białek. Po 1945 r. rozpoczęto systematyczne badania nad
drugorzędowa strukturą białek oraz ich struktura przestrzenna. Pierwsze badania nad tym
zagadnieniem podjęli Pauling i Corey. Następnie Kendrew i Purutz określili strukturę
mioglobiny i hemoglobiny, korzystając z zjawiska dyfrakcji promieni rentgena. Na podstawie
danych strukturalnych, można uzyskać wskazówki na temat funkcji biologicznych białek.
Podział białek
Białka są substancjami o skomplikowanej budowie, dlatego ciężko było by przeprowadzić ich
klasyfikacje na podstawie ich struktury. Z tego powodu białka są klasyfikowane na różne
sposoby, na przykład: na ich pochodzenie (białka roślinne, zwierzęce, wirusowe, bakteryjne)
lub na występowanie w różnych organach (białko plazmy, mięśni wątroby itp.), albo
organellach komórkowych (białko cytoplazmy białko rybosonalne, jądrowe, błon itp.). Jako
inne kryterium można przyjąć ich funkcje biochemiczne np.(białka enzymatyczne, ochronne,
strukturalne, strukturalne, zapasowe, transportowe). Funkcjami biologicznymi białka zajmie
się w dalszej części swej pracy.
Najwcześniej stosowany podział uwzględnia różnice w rozpuszczalności i kształcie
cząsteczek.
Białka globularne: rozpuszczalne w wodzie i rozcieńczonych roztworach soli, przybierają
kształt kulisty.
Białka fibrylarne: nie rozpuszczalne w roztworach wodnych i roztworach soli, struktury
włókniste występują na poziomie makroskopowym. Są odporne na działanie zasad i kwasów.
Obecnie stosuje się podział białek pod względem składu chemicznego i dzieli białka na dwie
grupy:
Białka proste - w wyniku całkowitej hydrolizy tych białek powstają tylko same aminokwasy.
Białka złożone - w wyniku całkowitej hydrolizy tych białek powstają aminokwasy i składniki
nie białkowe.
6
Podział ten jest stosowany do dnia dzisiejszego, chociaż badania najnowocześniejszą
aparaturą wykazały, że niektóre białka proste błędnie zostały zaliczone do tej grupy.
Białka proste
a. Albuminy
Albuminy są białkami o małej masie cząsteczkowej. Dobrze rozpuszczają się w wodzie i
roztworach soli w zakresie pH od 4 do 8,5, a także łatwo krystalizują. Można je wytrącić
bardzo stężonym siarczanem VI amonu. Kwasowy charakter albumin wynika z dużej
zawartości (20-25%) kwasu glutaminowego i asparaginowego.
Do tej grupy zalicza się: albuminy surowicy, a-laktoalbuminy, owalbuminę jaja kurzego oraz
albuminy roślinne.
Albuminy surowicy- (masa cząsteczkowa 67 000 u) jest głównym składnikiem białek plazmy,
występującym w ilości około 60%. Najważniejszą funkcją tego białka jest regulacja ciśnienia
osmotycznego krwi. Wykazuje ono dużą zdolność wiązania jonów K+, Na+, Ca2+, jak
również hormonów, kwasów tłuszczowych i leków. Cząsteczki albuminy surowicy ludzkiej
są łańcuchami polipeptydowymi złożonymi z 579 aminokwasów, powiązanymi poprzecznie
17 mostkami disulfidowymi.
a-laktoalbumina- występuje w mleku ssaków. Jej cechą charakterystyczna jest duża
odporność na wysoka temperaturę. Odmiennie od albumin surowicy, a-laktoalbumina zawiera
oligosacharyd, który jest przyłączony do łańcucha plipeptydowego przez grupę bkarboksylową reszty kwasu asparaginowego. Ludzka a-laktoalbumina (masa cząsteczkowa 14
176 u) składa się z 123 reszt i zawiera cztery mostki disiarczkowe. Pierwszorzędowa struktura
tego białka jest zgodna ze strukturami a-laktoalbuminy u innych gatunków ssaków. Analiza
struktur wykazała, że a-laktoalbumina zawiera 30% aminokwasów tworzących strukturę a i
14% aminokwasów tworzących strukturę b. Wykazano, że łańcuch a-laktoalbumina jest
pofałdowana w podobny sposób jak cząsteczka lizozymu.
Owalbumina- (masa cząsteczkowa 44 000 u) zawiera 3,2% węglowodanów oraz resztę kwasu
fosforowego, która przyłączona jest do łańcucha bocznego seryny.
Albuminy roślinne występują w małych ilościach w nasionach, np. rycyna w nasionach
rącznika, leukazyna w nasionach różnych zbóż i nasionach roślin strączkowych. Rycyna
wykazuje właściwości toksyczne.
Rycyna (masa cząsteczkowa 65 000 u) zawiera 493 reszty aminokwasowe. Składa się z
dwóch łańcuchów, powiązanymi ze sobą mostkami disiarczkowymi. Chociaż toksyczny jest
łańcuch A, nie mniej ważny jest łańcuch B, ponieważ zawiera on miejsce łączące się z
powierzchnia komórki. Rycyna działa jako inhibitor biosyntezy białek i wykazuje działanie
antynowotworowe.
b. Globuliny
Globuliny mają większą masę cząsteczkowa niż albuminy i występują praktycznie w
wszystkich komórkach zwierzęcych i roślinnych. Do tej jednolitej i szerokiej klasy białek
należy wiele enzymów, białek plazmy, przeciwciał, białek mleka, glikoproteidów i białek
zapasowych roślin. Ponieważ wiele globulinów zawiera słabo przyłączone węglowodany nie
ma zgodności, co do klasyfikacji tych związków. Globuliny są nie rozpuszczalne lub słabo
rozpuszczalne w wodzie pozbawionej soli. Globuliny można frakcjonować, wytrącając
stopniowo siarczanem VI amonu.
a-laktoglobuliny- (masa cząsteczkowa 36 500 u) jest najważniejszym składnikiem białkowym
mleka krowiego. Składa się z dwóch identycznych łańcuchów polipeptydowych
zawierających po 162 reszty aminokwasowe. a-laktoglobulina świńska zawiera tylko jeden
łąńcuch. Tylko 10 - 17% aminokwasów tworzy strukturę a natomiast 30- 45% tworzy
strukturę b.
7
Globuliny plazmy- są klasyfikowane na podstawie ich zachowania się w czasie elektroferazy
na a1, a2, b i c-globuliny. c-globuliny są przeciw ciałami. Przeciwciała są
immunoglobulinami produkowanymi przez komórki limfatyczne w odpowiedzi na obecność
obcego antygenu.
Haptaglobulina- jest tetramerem, składającym się z dwóch różnych par łańcuchów (a2 i b2),
które są powiązane mostkami disiarczkowymi. Heptoglobina tworzy kompleks z
oksyhemoglobiną, w wyniku, czego zachodzi zmiana konformacji pierścienia porfirynowego,
który może być otwarty przez działanie enzymu.
Globuliny warzyw- należą do nich arachina, zeina oraz tuberyna. Edestyna jest przykładem
białek nasion konopi. Jej masa cząsteczkowa wynosi 300 000 u. Edestyna składa się z sześciu
podjednostek, z których każda skonstruowana jest z dwóch identycznych łańcuchów
polipeptydowych, połączonych wiązaniami disiarczkowymi. Arachina jest białkiem
orzeszków ziemnych, ma strukturę heksameru. Każda z podjednostek stanowi dwa łańcuchy
powiązane ze sobą kowalencyjnie.
c. Histony
Histony są białkami zasadowymi (zawierają dużo argininy i lizyny) o małej masie
cząsteczkowej, rozpuszczalnymi w kwasach i wodzie. Białka te nie są specyficzne dla
określonej tkanki i występują we wszystkich komórkach eukariotycznych, gdzie tworzą
kompleksy z DNA, a zatem odgrywają ważną rolę w procesie represji genów.
Histony składają się z 101 lub 212 aminokwasów, a ich masa cząsteczkowa zawiera między
11 200 a 21 000 u. Prawie we wszystkich histonach znajdują się reszty metylowanych lub
acetylowanych aminokwasów, w tym z 20 reszt lizynowych i 8 arginowych, a N-końcowa
prolina nie jest podstawiona.
Histony w zasadzie nie zmieniły swej budowy ewolucyjnej na przestrzeni milionów lat. Rolą
histonów jest kondensowanie chromatyny w czasie tworzenia się chromosomów. Chromatyna
nawija się na oktamer histonowy. Oktamer jest zbudowany z 6 cząsteczek histonów i tworzy
rdzeń na, który nawijany jest DNA powstaje w wtedy nukleosom. Podstawową funkcją
histonów jest blokowanie DNA, powoduje to korzystanie tylko z tej części materiału
genetycznego, która jest potrzebna komórce do pełnienia swych funkcji biologicznych.
d. Prolaminy
Prolaminy charakteryzują się znaczną zawartością kwasu glutaminowego (30-45%) i proliny
(15%). Prolaminy są nie rozpuszczalne w wodzie i roztworach soli, ale rozpuszczają się w 5090% roztworze etanolu. Gliadyna (pszenica i żyto) oraz hordenina (jęczmień) są ważnymi
białkami zbóż i typowymi przykładami związków tej klasy.
e. Gluteliny
Gluteliny zawierają w swej budowie dużo kwasu glutaminowego do 45%. Gluteliny są
rozpuszczalne w wodzie, roztworach soli i rozcieńczonych alkoholach, a także w roztworach
zasadowych i kwaśnych. Do tej grupy związków należą: glutenina (pszenica), orzynina (ryż),
hordenina (jęczmień). Glutenina i gliadyna składa się na lepkie białko (gluten) mąki pszenej.
Przydatność tej mąki jest związana od zawartości glutenu. Masy tych białek wynoszą od 50
000 do kilku milionów Da. Glutenina składa się z podjednostek o masie 150 000 Da.
Podjednostki te są łańcuchami polipeptydowymi i można je rozdzielić za pomocą
elektroforezy.
f. Protoaminy
Białka należące do tej grupy są bardziej zasadowe niż histony, co wynika z większej
zawartości argininy (80-85%). Są rozpuszczalne w wodzie i kwasach. Znajdują się w jądrach
komórkowych w połączeniach z DNA. Protoaminy mają małe masy cząsteczkowe 4000-4500
u. Struktura pierwszorzędowa protoamin często charakteryzuje się powtarzalnością tej samej
sekwencji.
8
Właściwości chemiczne
Amfoteryczność
Amfoteryczny charakter białek jest wynikiem obecności i rozmieszczenia kwasowych i
zasadowych grup łańcuchów bocznych, ponieważ w długim łańcuchu polipeptydowym
końcowe grupy aminowa i karboksylowa, odwrotnie niż w przypadku aminokwasów, mają
mały udział w tworzeniu dipolarnego charakteru cząsteczki.
W roztworze kwaśnym białka mają ładunek dodatni w roztworze zasadowym ujemny, a ich
stopień hydratacji i rozpuszczalność są największe w roztworach o skrajnych wartościach pH.
Ruchliwość elektroforetyczną określa ładunek wypadkowy cząsteczki. W punkcie
izoelektrycznym ładunki dodatnie i ujemne równoważą się. W punkcie izoelektrycznym
białko wykazuje minimalną rozpuszczalność i minimalny stopień hydratacji. Punkt
izoelektryczny białka można określić przez: określenie minimalnej rozpuszczalności w
roztworach buforowych, elektroforetycznie przy różnym pH lub metodą ogniskowania
izoelektrycznego. Białka zawierające stosunkowo dużo aminokwasów zasadowych maja
wartość pI w zakresie dużych wartości pH (protoamina pI 11,8 pH), natomiast białka a z dużą
ilością aminokwasów kwasowych mają pI w zakresie niskich wartości pH (pepsyna pI 1 pH).
Ważny fizjologicznie efekt buforowania białek zależy od równowagi:
W stabilizacji pH krwi dużą rolę odgrywa hemoglobina. Normalna wartość pH krwi wynosi
7,35 - 7,40. Obniżenie go o 0,5 pH zagraża życiu.
2.4.2. Wysalanie
Niewielkie stężenie soli w nieorganicznych zwiększa rozpuszczalność białek. Jeżeli jednak
stężenie soli będzie wzrastać, korzystny wpływ jonów soli na rozpuszczalność białka ustaje, a
przy dużym stężeniu soli białka rozpuszczalne w wodzie będą się wytrącać z roztworów
wodnych. Proces ten nazywa się wysalaniem.
Działanie wysalające dużych stężeń jonów mało cząsteczkowych polega na ich znacznie
większym oddziaływaniu na cząsteczki wody w porównaniu z siłami wywołującymi
uwodnienie cząsteczki białka. Sole mające możność wiązania wody rywalizują z
cząsteczkami białka i odbierają im "płaszcz wodny", co zmniejsza ich rozpuszczalność i
prowadzi do wytracenia z roztworu. Stężenie soli potrzebne do wysolenia zależy od
właściwości białka i od pH środowiska. Białka najłatwiej wysolić w ich punkcie
izoelektrycznym, gdyż wtedy ich rozpuszczalność jest najmniejsza. Solą najczęściej
stosowaną do wysalania jest siarczan VI amonu. Wysalanie białek jest procesem
odwracalnym, ponieważ przez obniżenie stężenia soli, np. dodanie rozpuszczalnika wytrącone
białko można ponownie rozpuścić. Białko takie zachowuje wszystkie swoje właściwości,
ponieważ wysalanie nie powoduje denaturacji białek.
Hydroliza
Hydroliza białek polega na depolimeryzacji białek na poszczególne aminokwasy. Hydrolizę
białek można przeprowadzać na trzy sposoby: pod wpływem kwasu, zasady lub enzymu.
9
Hydroliza kwasowa polega na ogrzewaniu białka w roztworze HCl o stężeniu 2 mol lub
roztworze H2SO4 o stężeniu 4 mol przez 18-24 h. Jednak w warunkach tych całkowitemu
rozkładowi ulega tryptofan.
Podczas hydrolizy zasadowej najczęściej ogrzewa się białko z NaOH o stężeniu 2 mol.
Metoda ta jest praktycznie nie stosowana, ponieważ zupełnie niszczy argininę, cysteinę i
treoninę.
Najlepszą metodą hydrolizy białek jest hydroliza enzymatyczna, gdyż zniszczeniu nie ulegają
żadne aminokwasy. Wadą tej metody jest powolność procesu, a co za tym idzie długi czas
depolimeryzacji białka.
Metoda hydrolizy enzymatycznej znalazła zastosowanie w określaniu sekwencji białek.
Denaturacja
Denaturacją białka nazywamy takie przeprowadzenie metodami fizycznymi lub chemicznymi,
zmian w strukturze białka, które prowadza do mniejszej lub większej utraty aktywności
biologicznej białka, przy czym jego struktura pierwszorzędowa nie zostaje zmieniona.
Podczas denaturacji w dużej mierze zniszczeniu ulegają wiązania wodorowe, a w obecności
odczynników redukcyjnych rozszczepieniu ulegają wiązania disiarkowego. Często
denaturacja jest odwracalna i po usunięciu czynnika denaturującego białko wraca do
pierwotnej postaci. Proces ten nazywa się renaturacją. Renaturacja jest też możliwa w
przypadku denaturacji redukcyjnej, ale wiele przypadków denaturacji jest nieodwracalnych.
Podczas denaturacji zachodzą takie zmiany jak: zmniejszenie rozpuszczalności, może się w
wyniku wyeksponowania nowych zjonizowanych grup przesunąć pH punktu
izoelektrycznego. Rozwinięcie łańcucha może prowadzić do zwiększenia się lepkości, a także
absorpcji w nadfiolecie. Obserwuje się też procesy agregacji i wytrącania, co jest związane z
ze zmianą stopnia hydratacji i rozpuszczalności białek, a także zmianami w mostkach
disiarkowych.
Fizycznymi metodami denaturacji białek są:
Silne mieszanie, ogrzewanie, naświetlanie promieniami UV, rentgenowskimi i c lub działanie
ultradźwiękami.
Denaturacja chemiczna zachodzi pod wpływem takich związków jak:
Roztwór mocznika (6-8 mol/dm3) lub guanidyny (4 mol/dm3), na skutek działania kwasów
lub zasad (pH poniżej 3 lub powyżej 9), na skutek działania detergentów, roztworów soli
ciężkich lub niektórych substancji organicznych takich jak: aceton, benzen, formaldehyd,
benzyna. Poszczególne białka różnią się zdolnością do denaturacji.
Reakcje charakterystyczne
Białka ulegają różnym reakcjom charakterystycznym determinowanym przez właściwości
chemiczne reszt aminokwasowych. Reakcja biuretowa wykrywa obecność wiązania
peptydowego w białku. Reakcja ksantoproteinowa służy wykryciu obecności związków
aromatycznych w podstawniku tyrozyny i fenyloalaniny. Reakcja Millona też służy wykryciu
związków aromatycznych w podstawniku tyrozyny, a reakcja Pauly`ego ma takie samo
zadanie tylko w tyrozynie i tryptofanie. Wszystkie te reakcje są reakcjami barwnymi.
Reakcja biuretowa
10
Reakcja ta polega na tworzeni się związku kompleksowego pomiędzy jonem miedzi z
rozcieńczonego roztworu siarczanu VI miedzi, a wiązaniem peptydowym w białku w
środowisku zasadowym. Powstały kompleks przybiera barwę fioletową.
Reakcja ksantoproteinowa
Jest to reakcja stężonego kwasu azotowego V na białko. W wyniku tej reakcji związki
aromatyczne będące podstawnikami aminokwasów ulegają reakcji nitrowania. Białko
przyjmuje żółty kolor związków nitrowych. Produkt reakcji przeprowadzanej w środowisku
alkalicznym ma kolor pomarańczowy.
Bibliografia
Hans Dietrich Jakube - Aminokwasy, peptydy, białka
Tadeusz Gałamon - Chemia Repetytorium dla kandydatów na akademie medyczne
Waldemar Lewiński - Molekularne podłoże biologii
Waldemar Lewiński - Anatomia i fizjologia człowieka (z elementami fizjologii zwierząt)
Waldemar Lewiński - Genetyka
Pod redakcją Witolda Seńczuka - Toksykologia
Pod redakcją Piotra Laidlera, Teresy Zgliczyńskiej - Ćwiczenia z chemii i Biochemi dla
studentów Medycyny i stomatologii
Pod redakcją Lidii Włodek - Biochemia ćwiczenia praktyczne dla studentów wydziału
farmacji
B. D. Hans, N. M. Hooper - Biochemia krótkie wykłady
Leokadia Stefanowicz - Cytobiochemia
Pod redakcją Wacława Minakowskiego - Biochemia Kręgowców
Hary Matthews, Richard Fredlana, Roger Miesfeld - Biochemia I biologia molekularna w
zarysie
11