Podstawy histologii
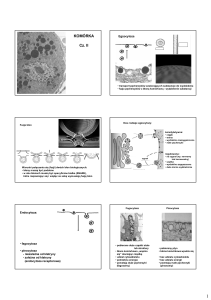
KOMÓRKA
Wielkość komórek: 4-150 mikrometrów (m), przeciętnie ok. 20 m.
Elementy składowe komórki: (1) błona komórkowa, (2) jądro, (3) organelle (4) cytoszkielet,
(5) cytoplazma podstawowa
Błona komórkowa i prawie wszystkie organelle zbudowane są z błon biologicznych
Błony biologiczne
Zbudowane są z
(1) dwuwarstwy fosfolipidowej - cząsteczki fosfolipidów układają się w dwie warstwy
zwrócone do siebie grupami hydrofobowymi, a na zewnątrz grupami hydrofilnymi;
dwuwarstwa jest półpłynna i decyduje o integralności błony; oprócz fosfolipidów w jej skład
wchodzą glikolipidy (tylko w błonie komórkowej, uczestniczą w tworzeniu glikokaliksu, p.
dalej) i cholesterol (zwiększa sztywność błony);
(2) białek błonowych - białka związane są z zewnętrzną lub wewnętrzną powierzchnią
dwuwarstwy (białka powierzchniowe) lub przechodzą przez całą grubość dwuwarstwy (białka
transbłonowe); białka mogą się przemieszczać w płaszczyźnie dwuwarstwy (wyjątek - białka
związane z cytoszkieletem, p. dalej).
Funkcjonalna klasyfikacja białek błonowych:
a) białka strukturalne (wzmacniają strukturę błony, przytwierdzają do niej cytoszkielet,
odpowiadają za przyleganie i łączenie się komórek)
b) białka enzymatyczne (katalizują reakcje biochemiczne związane z błoną)
c) białka transportowe (transportują substancje niskocząsteczkowe przez błonę)
d) białka receptorowe (rozpoznają i wiążą cząsteczki sygnałowe - np. hormony) i inicjują
odpowiedź komórki na te substancje
Niektóre białka mogą jednocześnie pełnić różne funkcje.
Błona biologiczna jest barierą dla substancji chemicznych i kontroluje ich transport pomiędzy
środowiskiem a komórką i pomiędzy poszczególnymi przedziałami wewnątrzkomórkowymi.
Niektóre substancje (np. gazy, cząsteczki hydrofobowe: kwasy tłuszczowe, hormony
steroidowe) mogą swobodnie dyfundować przez dwuwarstwę fosfolipidową. Cząsteczki
hydrofilne transportowane są poprzez białka transportowe.
A. Transport substancji niskocząsteczkowych (jony, woda, substancje proste):
a) kanały jonowe: mogą być otwarte lub zamknięte, w stanie otwartym pozwalają na dyfuzję transport jonów zgodnie z gradientem stężeń, bez nakładu energii. W zależności od
mechanizmu otwierania wyróżniamy: (1) kanały otwierane zmianą potencjału (elektrycznego
błony), (2) kanały otwierane ligandem (czyli przez przyłączenie określonej substancji, taki
kanał ma część będącą receptorem) i (3) kanały otwierane mechanicznie (rzadkie);
b) przenośniki: zmieniając swoja konformację przenoszą substancje proste przez błonę
zgodnie z gradientem stężeń, bez nakładu energii - transport ułatwiony;
c) pompy: zmieniając konformację i wykorzystując energię z rozkładu ATP przenoszą jony i
substancje proste przez błonę wbrew gradientowi stężeń - transport aktywny (np. pompa
sodowo-potasowa błony komórkowej). W innym procesie jedne jony przenoszone są wbrew
gradientowi stężeń wykorzystując energię z przepływu innego jonu zgodnie z gradientem transport aktywny wtórny (np. wymiennik wapniowo sodowy w mięśniówce serca).
Przenośniki i pompy mogą transportować tylko jedną substancję (uniport), lub równocześnie
dwie różne substancje (kotransport) – jeżeli przenoszą je w tym samym kierunku nazywamy to
symportem, a jeżeli w przeciwnych kierunkach – antyportem.
B. Transport substancji wysokocząsteczkowych i niekiedy dużych struktur – tzw. transport
pęcherzykowy: błona wytwarza pęcherzyki zawierające transportowaną substancję/strukturę,
transportowane są zatem zarówno fragmenty błony (błona pęcherzyka), jak i zawartość
pęcherzyka. W trakcie fuzji (połączenia) błony pęcherzyka z błoną komórkową (lub inną
błoną) nie dochodzi do przerwania jej ciągłości.
Transport pęcherzykowy przez błonę komórkową:
- endocytoza (do wnętrza komórki): fagocytoza, pinocytoza, endocytoza receptorowa
(swoista)
- egzocytoza (z komórki na zewnątrz) – w ten sposób odbywa się wydzielanie
Transport pęcherzykowy pomiędzy różnymi organellami i błoną komórkową nosi nazwę
przepływu błon w komórce.
Główne szlaki przepływu błon w komórce:
szlak wydzielniczy: siateczka śródplazmatyczna → aparat Golgiego → błona komórkowa
szlak endocytozy: błona komórkowa → pęcherzyki endocytotyczne (endosomy) →
lizosomy
Błona komórkowa oddzielająca komórkę od środowiska zewnętrznego, ma szczególne cechy:
- jest najgrubsza spośród wszystkich błon biologicznych (7,5 nm)
- w obrazie z mikroskopu elektronowego ma wyraźna trójwarstwową strukturę (dwie
zewnętrzne warstwy ciemne odpowiadają hydrofilnym odcinkom fosfolipidów, środkowa
warstwa jasna odcinkom hydrofobowym)
- posiada szczególnie liczne białka receptorowe i transportowe
- zawiera cząsteczki adhezyjne – specyficzne białka odpowiedzialne za łączenie komórek
między sobą (selektyny, kadheryny i cząsteczki z nadrodziny immunoglobulin) i z otoczeniem
czyli substancją międzykomórkową (integryny);
- wykazuje tzw. potencjał spoczynkowy – różnicę potencjału elektrycznego po jej obu
stronach wynikająca z nierównomiernego rozmieszczenia jonów; za wartość potencjału
spoczynkowego w głównej mierze odpowiadają kanały sodowe, potasowe i pompa sodowopotasowa;
- na zewnętrznej powierzchni pokryta jest glikokaliksem - warstewką cukrowców
wchodzących w skład glikoproteidów i glikolipidów błony. Glikokaliks pełni funkcje
ochronne, a także uczestniczy w procesach endocytozy oraz w kontaktowym rozpoznawaniu
się i łączeniu komórek.
Jądro komórkowe
Funkcje:
a) magazynuje informację genetyczną (DNA)
b) powiela informację genetyczną przed podziałem (replikacja DNA)
c) wytwarza podjednostki rybosomów (jąderko)
d) kieruje wszystkimi procesami życiowymi komórki poprzez sterowanie syntezą białek
DNA → (transkrypcja) → RNA → (translacja) → białko
Elementy składowe jądra:
1. Chromatyna jądrowa: zawiera DNA i białka (histony i białka niehistonowe), koduje
informację genetyczną, reguluje wszystkie procesy komórkowe poprzez sterowanie syntezą
białek. Chromatyna występuje w dwóch formach:
euchromatyna - luźna (jasna), aktywna transkrypcyjnie,
heterochromatyna - zbita (ciemna), nieaktywna transkrypcyjnie.
Najmniejszą strukturalną jednostką chromatyny jest nukleosom, zbudowany z rdzenia
utworzonego przez histony, wokół którego owija się fragment nici DNA. Połączone nicią DNA
nukleosomy tworzą nukleofilament – jest to najluźniejsza forma chromatyny, umożliwiająca
zachodzenie procesów replikacji i transkrypcji. Kolejne etapy zagęszczania chromatyny to
agregacja nukleosomów w włókno chromatynowe, tworzenie bocznych pętli, wtórna
spiralizacja i wreszcie (tylko przed podziałem komórki) wytworzenie chromatydy i
chromosomu.
2. Jąderko: w mikroskopie świetlnym widoczne jako ciemna grudka/grudki (w jądrze może
być kilka jąderek), w mikroskopie elektronowym można w nim wyodrębnić 3 rodzaje
obszarów:
- jasne centra włókienkowe, zawierające nieaktywny rDNA (kodujący rRNA),
- gęste obszary włókienkowe, gdzie odbywa się transkrypcja i powstaje pre-rRNA,
- obszary ziarniste, zawierające świeżo wytworzone podjednostki rybosomów.
„Produkcja” podjednostek rybosomów rozpoczyna się od transkrypcji pre-rRNA, który
następnie jest cięty na fragmenty odpowiadające poszczególnym rodzajom rRNA, a te z kolei
są łączone z białkami rybosomowymi importowanymi do jąderka z cytoplazmy. Powstałe w ten
sposób podjednostki rybosomów (zwłaszcza duże) przez pewien dojrzewają na terenie
obszarów ziarnistych jąderka, a następnie przechodzą z jadra do cytoplazmy. Na terenie jądra
podjednostki nigdy nie łączą się w kompletne rybosomy – dochodzi do tego w cytoplazmie, na
początku procesu translacji.
3. Otoczka jądrowa: zbudowana z dwóch błon biologicznych – zewnętrzna ma połączenie z
błonami siateczki śródplazmatycznej, a do wewnętrznej przylega od strony jądra tzw. blaszka
jądrowa – pokład filamentów pośrednich (p. cytoszkielet) zbudowanych z białek lamin.
Pomiędzy błonami znajduje się wąska przestrzeń okołojądrowa.
W otoczce znajdują się pory jądrowe, umożliwiające wymianę substancji między jądrem a
cytoplazmą. Pojedynczy por (tzw. kompleks poru jądrowego) składa się z trzech białkowych
pierścieni zawierających receptory rozpoznające przenoszone substancje i białka
odpowiedzialne za ich przenoszenie przez por. Substancje niskocząsteczkowe przechodzą przez
por swobodnie, a selektywne przenoszenie dotyczy substancji wysokocząsteczkowych:
z jądra do cytoplazmy przechodzą: mRNA, tRNA, podjednostki rybosomów,
z cytoplazmy do jądra przechodzą: białka jądrowe (histony, białka niehistonowe,
enzymy) i białka rybosomowe (na terenie jądra nie odbywa się synteza białek).
Rybosomy
Rybosomy to kompleksy rybonukleoproteinowe w formie ziarenek o wielkości 30 nm,
odpowiadają za syntezę białek w komórce (translację). Rybosom zbudowany jest z dwóch
podjednostek: dużej (3 rodzaje rRNA i 49 białek) i małej (1 rodzaj rRNA i 33 białka).
Podjednostki łączą się tworząc rybosom dopiero po zainicjowaniu translacji, a po ukończeniu
tego procesu ponownie oddzielają się od siebie. Podczas translacji z jedną nicią mRNA łączy
się wiele rybosomów, tworząc polisom (polirybosom). W komórce rybosomy mogą być:
związane z błonami szorstkiej siateczki śródplazmatycznej - produkują białka błonowe,
wydzielnicze i enzymów lizosomalnych,
wolne w cytoplazmie - produkują białka jądrowe, mitochondriów, peroksysomów,
cytoszkieletu, cytoplazmy.
Siateczka śródplazmatyczna
Jest to labirynt wewnątrzkomórkowych błon (cysterny, kanaliki) łączących się ze sobą.
Rodzaje:
szorstka – z rybosomami, ma formę cystern, główna funkcja – produkcja białek i ich
wstępna glikozylacja (przyłączanie łańcuchów cukrowcowych);
gładka – bez rybosomów, ma formę kanalików, funkcje: produkcja lipidów, neutralizacja
leków i trucizn, gromadzenie jonów wapnia.
Białka wytworzone na rybosomach siateczki wbudowywane są do jej błony (białka
błonowe) lub dostają się do jej wnętrza. Na drodze transportu pęcherzykowego (przepływu
błon) białka te mogą się przemieszczać do aparatu Golgiego, błony komórkowej, pęcherzyków
hydrolazowych i lizosomów.
Aparat Golgiego
Jest zbudowany z podjednostek (diktiosomów) utworzonych przez kilka wygiętych,
błonowych cystern. Każdy diktiosom ma dwa bieguny:
cis (wypukły), skierowany w stronę siateczki śródplazmatycznej; jego błona jest
podobna do błony siateczki śródplazmatycznej;
trans (wklęsły), skierowany w stronę powierzchni komórki; jego błona jest podobna
do błony komórkowej.
Przez diktiosom „przepływają” (transportem pęcherzykowym) fragmenty błony i białka z
siateczki śródplazmatycznej. Małe pęcherzyki oddzielają się od siateczki i łączą z błoną
diktiosomu po stronie cis (tzw. sieć cis). Podczas przechodzenia przez kolejne cysterny
diktiosomu, do białek błony i białek transportowanych wewnątrz pęcherzyków zostają
przyłączane grupy cukrowcowe (glikozylacja), niekiedy również fosforanowe i siarczanowe, a
błona stopniowo zmienia charakter na podobny do błony komórkowej. W ostatniej cysternie po
stronie trans (tzw. sieć trans) białka są sortowane i „pakowane” do różnych pęcherzyków, które
oddzielają się od tej cysterny:
- małe pęcherzyki z materiałem do wydzielenia na drodze egzocytozy konstytutywnej,
- duże pęcherzyki (ziarna wydzielnicze) z materiałem do wydzielenia na drodze egzocytozy
regulowanej,
- pęcherzyki hydrolazowe zawierające enzymy trawienne (prekursory lizosomów).
Funkcje aparatu Golgiego:
przebudowa błon (typ siateczki → typ błony komórkowej), odnowa błony komórkowej
glikozylacja przepływających białek, ich sortowanie i kierowanie do różnych pęcherzyków
tworzenie pęcherzyków i ziarn wydzielniczych oraz pęcherzyków hydrolazowych.
Egzocytoza
Egzocytoza jest mechanizmem wydzielania komórkowego. Wyróżnia się 2 typy tego procesu:
egzocytoza konstytutywna: ciągła, wolna, za pośrednictwem małych pęcherzyków;
egzocytoza regulowana: wywołana sygnałem (np. nerwowym lub hormonalnym),
szybka, uczestniczą w niej ziarna wydzielnicze – duże pęcherzyki z zagęszczoną
zawartością (w ten sposób wydzielają prawie wszystkie komórki gruczołowe).
Endocytoza
Istnieją trzy odmiany tego procesu.
1. Fagocytoza: na tej drodze pobierane są duże cząstki stałe lub struktury (np. bakterie).
Błona komórkowa wysuwa wypustki otaczajace pobieraną strukturę – jest to proces
wymagający nakładu energii i udziału cytoszkieletu, a w jego wyniku tworzą się duże
pęcherzyki (fagosomy). Fagocytoza często wymaga wstępnego rozpoznania
pobieranego materiału przez receptory błonowe. Za proces ten odpowiadają
wyspecjalizowane komórki (np. makrofagi, granulocyty).
2. Pinocytoza: pobierany jest płyn wraz z zawartymi w nim substancjami, błona
komórkowa wpukla się bez udziału energii i cytoszkieletu, powstają małe pęcherzyki
(pinosomy). Zachodzi we wszystkich komórkach.
Endocytoza receptorowa: warunkiem koniecznym jest wstępne związanie pobieranych
substancji z receptorami błonowymi, błona komórkowa zachowuje się tak jak w
pinocytozie, ale niezbędny jest udział specjalnego białka (klatryny) otaczającego
powstający pęcherzyk..
Lizosomy
Lizosomy to pęcherzyki (0,1-2,0 m), w których zachodzi trawienie wewnątrzkomórkowe.
Powstają przez połączenie się pęcherzyków hydrolazowych (zawierających nieaktywne
enzymy trawienne – kwaśne hydrolazy), z:
pęcherzykami powstałymi w wyniku endocytozy: fagosomami, pinosomami, późnymi
endosomami - tworzą się heterolizosomy
pęcherzykami zawierającymi własne struktury komórki przeznaczone do likwidacji tworzą się autolizosomy.
W lizosomach hydrolazy ulegają aktywacji (niskie pH), następuje trawienie zawartości,
produkty trawienia (substancje niskocząsteczkowe) przechodzą do cytoplazmy, gdzie mogą być
użyte do syntezy nowych związków i produkcji energii.
Mitochondria
Są to pałeczkowate lub kuliste organelle o wielkości 2-5 m, zbudowane z podwójnej błony
biologicznej. Błona wewnętrzna tworzy fałdy (grzebienie).
Przedziały mitochondrialne:
błona zewnętrzna: przepuszczalna dla substancji niskocząsteczkowych (< 5kDa);
przestrzeń międzybłonowa
błona wewnętrzna: bardzo bogata w białka, selektywnie przepuszczalna (liczne
transportery), zawiera układ przenośników elektronów (łańcuch oddechowy) i kompleksy
syntazy ATP w formie „grzybków mitochondrialnych”;
macierz (przestrzeń wewnętrzna), zawiera enzymy cyklu Krebsa i beta-oksydacji kwasów
tłuszczowych i aparat genetyczny mitochondrium (p. dalej).
Synteza ATP (fosforylacja oksydacyjna) jest główną funkcją mitochondriów. Wymaga
współdziałania enzymów cyklu Krebsa, łańcucha oddechowego i syntazy ATP. W trakcie
przenoszenia elektronów trzy składniki łańcucha oddechowego wykorzystują uwalnianą
energię do „pompowania” protonów z macierzy do przestrzeni międzybłonowej. Powstały w
ten sposób gradient protonowy służy jako źródło energii dla syntezy ATP
Inne funkcje mitochondriów:
β-oksydacja kwasów tłuszczowych
ostatnie etapy syntezy hormonów steroidowych
udział w regulacji poziomu Ca2+ w komórce
produkcja ciepła (specjalne mitochondria w komórkach tkanki tłuszczowej brunatnej)
udział w procesie apoptozy
Peroksysomy
Są to pęcherzyki (0,1-1,0 m) otoczone pojedynczą błoną zawierającą unikatowe dla niej
białka, w tym transportery dla substratów enzymów peroksysomowych. Wewnątrz
peroksysomów znajdują się enzymy: oksydazy peroksysomowe, które utleniając różne
substraty produkują nadtlenek wodoru, katalaza, która ten nadtlenek rozkłada, a także enzymy
ß-oksydacji kwasów tłuszczowych, enzymy biosyntezy lipidów oraz aminotransferazy.
Funkcje peroksysomów:
utlenianie różnych substratów, w tym detoksyfikacja,
rozkład nadtlenku wodoru,
ß-oksydacja długołańcuchowych kw. tłuszczowych,
synteza cholesterolu, kwasów żółciowych i eterolipidów (plazmalogenów),
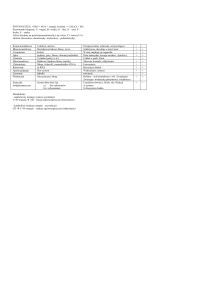
Cytoszkielet
Cytoszkielet to układy cienkich włókienek białkowych w cytoplazmie odpowiadające za
własności mechaniczne i ruch w komórce. Klasyfikacja:
Typ włókienek
mikrotubule
mikrofilamenty
filamenty pośrednie
Średnica
25 nm
6 nm
10 nm
Białko budujące
tubulina
aktyna
różne białka
Funkcja
ruch, podporowa
ruch, podporowa
podporowa
Mikrotubule są cienkimi rurkami. Wyróżnia się w nich koniec „+”, dynamiczny, na którym
może zachodzić polimeryzacja tubuliny (wydłużanie mikrotubuli) lub jej depolimeryzacja
(skracanie) oraz stabilny koniec „–” , zlokalizowany w pobliżu centrioli (p. dalej).
Mikrotubule tworzą struktury wyższego rzędu: aksonemy rzęsek (migawek) i witek (p. tkanka
nabłonkowa) oraz centriole. W tych strukturach mikrotubule łączą się bocznie w pary (dublety)
lub trójki (triplety).
Centriola to walec o długości 0,5 m, którego ścianę tworzy 9 tripletów (trójek) mikrotubul.
Centriole inicjują i kontrolują wzrost nowych mikrotubul, a także uczestniczą w podziale
komórki (tworzą bieguny wrzeciona podziałowego). Dwie centriole ułożone pod kątem
prostym, w większości komórek zlokalizowane w pobliżu jądra, to centrosom.
Mikrofilamenty (filamenty aktynowe) – podobnie jak mikrotubule mają koniec „+” i „–”.
Mikrotubule i mikrofilamenty uczestniczą w zjawiskach ruchowych komórki poprzez
współpracę z mechanoenzymami (białkami motorycznymi), które "kroczą" po ich
powierzchni zużywając ATP i do których mogą przyczepiać się inne struktury;
mikrotubule współpracują z dwoma mechanoenzymami: dyneiną (kroczy w kierunku
końca „–”) i kinezyną (kroczy w kierunku końca „+” ) – w ten sposób wzdłuż mikrotubul
są przesuwane w obu kierunkach struktury wewnątrzkomórkowe - pęcherzyki, organelle,
chromosomy podczas podziału.
mikrofilamenty współpracują z miozyną, która kroczy w kierunku końca „+” zazwyczaj
zakotwiczonego w błonie komórkowej; ten mechanizm odpowiada za zmiany konfiguracji
błony (tworzenie wypustek, fagocytoza), ruch pełzakowaty i skurcz komórki.
Filamenty pośrednie są zbudowane z łańcuchów białkowych skręconych w formę liny
(wytrzymałe, elastyczne). Nie współpracują z mechanoenzymami, pełnią wyłącznie funkcje
podporowe (wewnątrz komórki i w połączeniach międzykomórkowych). Są zbudowane z
różnych białek, zależnie od miejsca występowania (specyficzność tkankowa).
Materiały zapasowe i wtręty cytoplazmatyczne
Komórki mogą gromadzić glikogen (w formie drobnych ziarenek, widocznych tylko w
mikroskopie elektronowym) i lipidy (w formie kropelek nieotoczonych błoną). W niektórych
komórkach wstępują tzw. wtręty: drobne kryształki lub ziarna barwnika (np. melaniny).
Informacje zbiorcze
Typowe wyposażenie komórki wydzielniczej produkującej białka:
szorstka siateczka śródplazmatyczna
aparat Golgiego
ziarna wydzielnicze
Typowe wyposażenie komórki wydzielającej związki cukrowcowo-białkowe (np. śluz):
aparat Golgiego
szorstka siateczka śródplazmatyczna
ziarna wydzielnicze
Typowe wyposażenie komórki produkującej hormony steroidowe:
gładka siateczka śródplazmatyczna
mitochondria z rurkowatymi (tubularnymi) grzebieniami
kropelki lipidowe w cytoplazmie
Typowe wyposażenie komórki transportujacej jony:
sfałdowana błona komórkowa na powierzchni transportującej
liczne mitochondria w tym rejonie komórki
Typowe wyposażenie komórki wchłaniającej:
sfałdowana błona komórkowa na powierzchni szczytowej (brzeżek szczoteczkowy)
ścisłe połączenia z sąsiednimi komórkami
liczne mitochondria
Typowe wyposażenie komórki fagocytującej:
liczne pęcherzyki endocytotyczne (endosomy)
liczne lizosomy i pęcherzyki hydrolazowe
mocno pofałdowana błona komórkowa (odzwierciedlenie endocytozy)