Biologia dla ambitnych
ŁAŃCUCH ODDECHOWY I JEGO BLOKERY
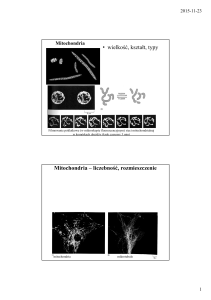
Mitochondria są odpowiedzialne za wytwarzanie energii w postaci
adenozynotrifosforanu (ATP). Te otoczone błonami struktury występują we wszystkich
komórkach eukariotycznych, poza czerwonymi krwinkami(erytrocytami) i ostatecznymi
keratynocytami. Występują licznie w komórkach zużywających duże ilości energii, np. w
komórkach mięśni szkieletowych i mięśnia sercowego, w komórkach egzokrynowych
trzustki, w plemnikach.
Wiele enzymów mitochondrialnych jest kodowana przez DNA jądrowe, ale
Mitochondria mają własne DNA, obdarzone zdolnością kodowania.
Zarówno w błonach, jak i macierzy mitochondrialnej znajduje się wiele enzymów
katalizujących przebieg dalszych (po glikolizie) procesów oddychania tlenowego.
W zewnętrznej, gładkiej błonie występują monoaminooksydazy, syntazy acetylokoenzymu A
(CoA) i fosfolipazy. Druga, grubsza błona wewnętrzna, jest wyspecjalizowaną błoną bogatą
w fosfolipid – kardiolipinę, mający cztery łańcuchy kwasów tłuszczowych związane z
glicerolem. Czyni ona błonę nieprzepuszczalną dla jonów. Ponadto błona ta zawiera białka
działające w reakcjach oksydacyjnych w czasie transportu elektronów przez łańcuch
oddechowy, kompleks enzymów syntazy ATP oraz białka transportowe, które regulują
transport metaboliczny do i z macierzy. Enzymy łańcucha oddechowego są zagłębione w
błonie i odpowiadają za fosforylację oksydacyjną, w wyniku której wytwarzany jest ATP.
Według teorii chemioosmozy Mitchell’a (Nagroda Nobla w 1975r.) ATP powstaje w wyniku
magazynowania energii podczas transportu protonów.
Pompa protonowa przenosi jony wodorowe (H+) w poprzek błony wewnętrznej do przestrzeni
śródbłonowej. Taki transfer protonów kończy się ustaleniem elektrochemicznego gradientu
protonowego w poprzek błony wewnętrznej. Gradient ten kieruje reakcjami, w których
wytwarzana jest energia w czasie, gdy protony płyną z powrotem „z góry na dół” przez
integralne białka błonowe osadzone w mitochondrialnej błonie wewnętrznej (np. białkiem
tym jest syntaza ATP).
Pompy protonowe są napędzane przez energię swobodną, która jest uwalniana podczas
przenoszenia kolejno elektronów przez łańcuch transportujący, nazywany systemem
transportu elektronów.
System transportu elektronów składa się z trzech dużych kompleksów białkowych
(I, II, III) i ubichinonu, odpowiedzialnych za kolejne dostarczenie wysokoenergetycznych
elektronów przenoszonych przez NADH i FADH2 na tlen cząsteczkowy. W każdym
kompleksie wyróżniamy:
- złożony z wielu podjednostek główny kompleks enzymatyczny, który katalizuje
przenoszenie elektronów
- grupy prostetyczne, które wiążąc się z enzymem przyjmują i uwalniają elektrony
- białka, które ułatwiają ruch protonów
Między trzema dużymi kompleksami oscylują dwa małe przenośniki energii: cytochrom c i
ubichinon. (rys. 1)
Transport elektronów zaczyna się, gdy dwa elektrony uwolnione z NADH są
przeniesione na pierwszy przenośnik elektronów w łańcuchu oddechowym. Mają one
najwyższą energię. Podczas przenoszenia elektronów wzdłuż łańcucha ( w praktyce elektrony
są przekazywane z atomów metalu występujących w grupach prostetycznych) energia jest
stopniowo rozpraszana. Elektrony wędrują wzdłuż łańcucha dopóki nie zostaną przeniesione
na tlen cząsteczkowy, który wykazuje największe powinowactwo do elektronów.
Grupy prostetyczne przechodzą cyklicznie między stanem utlenienia i redukcji,
gdy elektrony są przekazywane wzdłuż szlaku.
Pierwszy typ grupy prostetycznej (w II kompleksie) zawiera FAD, który w cyklu Krebsa
przyłącza elektrony z utleniania bursztynianu:
COO |
H–C–H
|
H–C–H
|
COO bursztynian
FAD
FADH2
COO –
|
H -C
||
C–H
|
COO fumaran
Rys. 1. Sprzężenie chemioosmotyczne i synteza ATP. Tworzenie ATP przez syntazę ATP jest
związane z przejściem protony przez wewnętrzną błonę ( u góry). Utlenianie w łańcuchu
oddechowym polega na przeniesieniu protonu z macierzy do przestrzeni śródbłonowej, co
chroni gradient protonowy (po stronie prawej). F1 i F0 są podjednostkami białkowymi, które
uczestniczą w fosforylacji.
Utlenione lub zredukowane części cząsteczek w FAD i mononukleotydzie
lawinowym, (FMN), które występują w kompleksie I wywodzą się z ryboflawiny (witamina
B2). Przenośniki te transportują pary elektronów i w czasie redukcji wiążą dwa atomy wodoru
tworząc FMNH2.
Drugą grupą przenośników są cytochromy, które posiadają grupę hemową z
centralnie położonym atomem żelaza zmieniającym stopień utlenienia między Fe3+ a Fe2+.
Trzecią grupę przenośników elektronów stanowią białka żelazo- siarkowe,
których dwa lub trzy atomy żelaza są kowalencyjnie związane z równą ilością atomów siarki,
w wyniku czego formuje się centrum żelazo- siarkowe. Podobnie, jak w cytochromach, atomy
żelaza zmieniają na przemian stan między Fe2+ a Fe3+ ,kiedy kompleks zyskuje i oddaje
elektrony.
Grupy prostetyczne czwartego kompleksu przenośników działają, gdy
przenośnikiem elektronów jest centrum z miedzią. Sądzi się, że stan atomu miedzi waha się
między Cu+ a Cu2+, kiedy kompleks jest zredukowany i utleniony. Miedź może uczestniczyć
w tworzeniu centrum bimetalicznego z atomem żelaza związanym z hemem w końcowym
stadium szlaku transportu elektronów.
Ubichinon nie jest związany z białkiem, a jego hydrofobowa cząsteczka jest rozpuszczalna w
warstwie lipidowej. Ubichinon może przyłączać jeden H+ dla każdego elektronu, który
przyłączył i może przenieść jeden albo dwa elektrony. Kiedy elektrony są transportowane do
następnego przenośnika w łańcuchu, protony są uwalniane.
W łańcuchu oddechowym bardzo ważną rolę odgrywają powiązania
chemioosmotyczne pomiędzy przenośnikami. Głównym jego celem jest uzyskiwanie i
akumulowanie energii w postaci ATP. Dlatego należy zwrócić uwagę na związki chemiczne,
które naruszają powiązania chemioosmotyczne i prowadzą do blokady syntezy ATP, gdyż ma
to skutki zdrowotne dla organizmu. Do tzw. „blokerów” należą między innymi:
- dinitrofenol – czynnik rozprzęgający, powodujący permeabilizację błony wewnętrznej dla
protonów
- oligomycyna – działa na syntazę ATP, blokując przejście protonów do macierzy
- malonian – blokujący przejście wodoru na FAD
- karboksyna, TTFA - blokujący oksydoreduktazy NADH, ubichinon; TTFA jest czynnikiem
helatującym żelazo
- amobarbital, rotenon – blokujący przejście wodoru i elektronów na ubichinon
- antymycyna A – blokująca przejście elektronu z kompleksu III (cytochrom b, FeS,
cytochrom c1) na dalsze cytochromy
- nieorganiczne związki chemiczne: H2S, CO, CN- - blokujące kompleks IV i przejście
elektronu na tlen.
Rys. 2. Hamowanie łańcucha oddechowego przez różne związki chemiczne. Wykres pokazuje
cztery kompleksy łańcucha oddechowego i możliwe miejsca działania różnych inhibitorów
chemicznych.
mgr Beata Urbańczyk
Literatura:
Solomon, Berg, Martin, Villee: „Biologia”
P. Hoser: „Fizjologia organizmów z elementami anatomii człowieka”
B.Filipowicz, W.Więckowski: „Zarys biochemii”
G.M.Fuller, D.Shields: „Podstawy molekularne biologii komórki”
Czasopismo „Świat nauki” na 10 (74) z X. 1997, artykuł pt. „Mitochondrialny DNA w
starzeniu się i chorobie” D.C.Wallace