bioch2.doc
(72 KB) Pobierz
1.Aktywatory i inhibitory enzymów allosterycznych.
Enzymy-białka katal. specyf. R.chem. Białka allost.- białko które może istnieć w 2 lub więcej konfor.zależ.od związ.innej cząst.(liganda) w miejscu innym niż miejsce
katal.tego białka. Złożone jest z wielu podjednostek często wykazują kooperatywną odp na związanie liganda. Mogą mieć więcej niż 1 miejsce aktywne. Aktywatoryutrwalają konformacje R, dobrze katalizują reakcje,wyłapują wszyst.konf.R i utrwalają,co zwiększa powinowactwo enzymu do substratu,co przyspiesza reakcje.
Inhibitory- utrwalają konf.T, żle katal.reak. wyłapują konf.T co zmniejsza powinow.enzymu do substratu, zwalnia przebieg reakcji.
Wyróżniamy inhibitory: odwracalne ( kompetatywne, niekompetat, pozakompeta.) i nieodwracalne.
2. Różnice DNA i RNA. DNA- dwuniciowy polinukleotyd , nośnik informacji genetycznej. Cukier- deoksyryboza.Zasady azotowe:A,T,C,G. Dwuniciowa helisa
zwinięta wokół 1 osi,przeważnie prawoskrętna,zasady ułożone do wewnątrz.Przeciwległe zasady łączą się wiązaniem wodorowym wg.zas.komplementarności A=T,
GC , nici biegną antyrównolegle, na pełen obrót przypada 10 zasad, inform.kodowana w nukleotydach. Replikacja może być tylko kontynuaowana. RNA- polimer
utworzony z rybo-nukleotydów. Mamy: rRNA, tRNA, mRNA. Cukier- ryboza, Zasady azotowe: A,U,C,G. Krótsze nici polinukleotydowe , jednoniciowa (ale może
tworzyć spinki”), ma fragm.helikowe i złożoną przestrzenną formę trójwymiarową, replikacja może zajść na od początku.
3. Lipidy występujące w org. żywych. Lipidy- cząst.org.zaw.łańcuchy węglowodorowe ,nierozp.w wodzie,rozp.w niepolarnych rozpusz.organ. Funkcje: * są
mater.budulcowym ( fosfolip) *decyd.o właści.dynam.błony komórk.*prekursory hormonów steroidowych(cholest) i tkankowych(kw.tłusz)*są subst.do syntezy
kw.tłusz.i wit.*biorą udział w zjawisk.immunolog ( eikozanoidy). Podział: proste i złożone. Proste : a)tłuszcze właściwe- estry kw.tłusz.z glicerolem b)woski-estry
kw.tł z wyższymi alkoh.jednowodorotl. Złożone(estry zaw.dodat.grupyfunkc.):1)fosfolipidy-zaw.reszte kw.fosforowegoi zas.azotowej lub aminoalkoholu.
Glicerofosfolipidy-zaw.glicerol. sfingolipidy-zaw.sfingozynę. 2) glikolipidy-zaw.kw.tłu.,alko,sfingozyne, węglowodan. 3)inne lip.złożone-sulfolipidy,
aminolip,lipoproteiny.
4.ATPaza mitochondrialna. ATP jest nukleotydem skł.się z adeniny,rybozy i trifosforanu. Główny nośnik energii w kom. Końcowe grupy fosforanowe są silnie
reaktywne i ich odłącz.na drodze hydrolizy lub przniesienia do innej cząst. zachodzi z wydzieleniem dużej ilości energii swobodnej. Jest stabilny,nie rozpada się
samorzutnie,wymaga enzymu. Syntez odbywa się gł.w mitochondrium.ATP powst. z ADP w wyniku fosforylacji poleg.na przyłączeniu przez ADP grupy fosforowej.
ATP jest szybko wykorzystywany tylko w miejscu jego powstania.Wew.błona mitoch.jest nieprzepuszczalna dla protonów,które mogą powrócić do matrixa jedynie
przez spec.kanały znajd.się w błonie utworz.przez cząst.enzymu syntazy ATP. Syntaza ATP tworzy kompl.zwane zesp.oddech. i wyst.w wew.pow.błony mitoch. W
miarę przesuwania się protonów z grad.energ.uwalnia się energia, którą syntaza ATP wykorz.do tworz.ATP. . Podjednostki syntezy ATP: *kompleks F1-katalizuje
syntezę ATP * kompl.Fo- kanał protonowy.oba umiejscowione w wew.błonie mitoch. Transport elektronów i syneza ATP są sprzężone przez gradient protonowy
utworz. w poprzek wew.błony mitoch. Przepływ e przez łańcuch oddech. Przepompowywanie e z matrixowej strony na przestrzeń między błonową. Wytworzona siła
protonomotoryczna w poprzek błony mitoch.powoduje transport e i napędza syntezę ATP przez kompleks ATPazy. Gdy kanał protonowy zostanie naruszony to jony
H+ nie będą mogły się przemieszczać( wrócić do matrix) => zatrzymanie syntezy ATP.
5.Kod genetyczny. Kodowanie. Kod gen-to współzależność między sekwencją zasad w DNA(lub mRNA) a sekwencją aminokwasów w białkach. Kod gen jest:
jednoznaczny, uniwersalny, zdegradowany, bezprzecinkowy, kolinearny, nie nakładający się. Kodowanie- jeden aminokwas jest kodowany przez grupę 3 zasad (
KODON). W DNA występują 4 rodz.zasad więc możliwości kodowania jest 4x4x4=64.. Sekwencja zasad odczytywana jest kolejno począwszy od kodonu start ( AUG)
do kodonu stop ( UAA,UAG,UGA).
1)Enzymy alloster. I te z r.M-M. Białka allost.- białko które może istnieć w 2 lub więcej konfor.zależ.od związ.innej cząst.(liganda) w miejscu innym niż miejsce
katal.tego białka. Złożone jest z wielu podjednostek często wykazują kooperatywną odp na związanie liganda. Mogą mieć więcej niż 1 miejsce aktywne. Nie spełniają
założ.równ.Mich-Mens. Wyst.w 2 formach R i T. Wiązanie substratu: * model sekwenccyjny TT TR RR i model jednoprzejściowy TT RR RR (+S na
strz.) Aktywatory- utrwalają konformacje R, dobrze katalizują reakcje,wyłapują wszyst.konf.R i utrwalają,co zwiększa powinowactwo enzymu do substratu,co
przyspiesza reakcje. Inhibitory- utrwalają konf.T, żle katal.reak. wyłapują konf.T co zmniejsza powinow.enzymu do substratu, zwalnia przebieg reakcji. Teoria M-M
opis.właśc.kinet.enzym. [E][S] / [ES]= Ks- stała substratowa. Wychodz.z określ.ilości enzymu i zwieksz.stęż.substr. az cały enzym przejdzie w kompleks [ES],szybkość
reak.wzrasta.
2) Podwójna helisa DNA. Helisa zbud.jest z dwóch wzajemnie oplatających się nici,razem tworzą helikalną strukturę w której zasady skierowane sa do wewnątrz a
rdzeń cukrowo-fosforanowy znajduje się na zewn. Nici ułożone są antyrównolegle względem siebie (jedna 3’->5’,druga 5’-> 3’) Cząst.polarna przy: C5’-gr.fosforanowa
C3’-gr.hydroksylowa. Nici utrzymują się razem dzięki słabym wiąz.wodorowym między zasadami purynowymi a pirymidowymi. Łączenie w pary na
zas.komplementarności A=T GC . Pojed.nić zbud.z nukleotydów.
3) Błona Biologiczna. kompleks białkowo-lipidowy będący jedną z podstawowych struktur budowy komórki organizmów prokariotycznych i eukariotycznych.Błonę
biologiczną charakteryzują:asymetria; płynność; półprzepuszczalność. Jest ona złożona z dwóch warstw lipidów, do których należą fosfolipidy, glikolipidy i steroidy,
oraz białek. Charakterystyczną budowę błony zapewnia amfipatyczność cząsteczek - zbudowane są z apolarnego ogona węglowodorowego oraz polarnej głowy, dzięki
czemu lipidy układają się w sferyczne pęcherzyki bądź właśnie dwuwarstwę. Dwuwarstwa lipidowa jest asymetryczna. Płynność dwuwarstwy lipidowej zależy od
charakteru łańcuchów lipidów, ich długości oraz obecności cholesterolu. Błona komórkowa jest strukturą półprzepuszczalną.
5) Ekspresja genów u prokariota i eukariota. Eukariota *rozdzielenie transkrypcji i translacji w czasie i przestrzeni. * najpierw transkypcja w jądrze,później w
cytoplazmie *geny nie ciągłe( introny i eksony) konieczna obróbka potranskryp. * AUG start metionina Prokariota * translacja i transkrypcja zachodzą jednocześnie
w tym samym miejscu * na powstałej nici mRNA od razu zaczyna się transkrypcja * brak intronów-nie wymaga obróbki potr. * AUG start – formylometionina.
1Inhibitory odwracalne: kompetatywne-zykle przypom.substr. łączy się z miejsc.akty.tworząc kompl.E1. Uniemożl.subst. dostęp do tego centr. Niekompetat.- wiąże
się z enzym w miejscu innym niż miejs.wiąza.substr. Wiąz.inhib. powod.zmniejsz.Vmax,, nie wpływa na Km. Pozakompet. Łączy się z enzymem w miejscu różnym
od miejsc.wiąz.subst, może łącz.się tylko z kompl.ES. Wpływa na Vmax i Km. Nieodwarcalny- nieszczą lub łaczą się z z gr.funkcyjną enzymu istotną dla jego
aktywności.częste połączenie.
3.Teoria chemiosmotyczna.Transport elektronów i fosforylacja oksydacyjna (synteza ATP) są powiązane z sobą przez gradient protonów w poprzek wewnętrznej
błony mitochondrium. Zgodnie z modelem chemiosmotycznym stopniowy przepływ elektronów z NADH lub FADH2 przez układ przenośników elektronów na tlen
powoduje uwalnianie energii. Jest ona wykorzystywana do przepompowania protonów przez wewnętrzną błonę mitochondrium do przestrzeni międzybłonowej.Protony
są przenoszone przez wewnętrzną błonę mitochondrialną przy udziale trzech kompleksów przenośników elektronów, które uczestniczą w poszczególnych etapach
transportu elektronów. Różnica w stężeniu protonów (H+) między matriks mitochondrium a przestrzenią miedzybłonową stanowi energię potencjalną (podobnie jak
woda spiętrzona w zaporze).Wewnętrzna błona mitochondrium jest nieprzepuszczalna dla protonów, które mogą powrócić do matriks jedynie przez specjalne kanały
znajdujące się w błonie wewnętrznej. Kanały te utworzone są przez cząsteczki enzymu – syntetazy ATP. Syntaza ATP tworzy kompleksy zwane zespołami
oddechowymi, występujące na wewnętrznej powierzchni błony mitochondrium. W miarę przesuwania się protonów zgodnie z gradientem energetycznym (tzn. poprzez
kanały syntetazy ATP na drugą stronę wewnętrznej błony mitochondrioum) uwalnia się energia, którą syntaza ATP wykorzystuje do tworzenia ATP.
5.Sacharoza – związek organiczny z grupy węglowodanów,Sacharoza, jest disacharydem, czyli cukrem złożonym z dwóch reszt monosacharydowych połączonych ze
sobą wiązaniem glikozydowym. Monosacharydami budującymi cząsteczkę sacharozy są : fruktoza (β-D-fruktofuranoza) oraz glukoza (α-D-glukopiranoza). Grupy te
są połączone za pomocą wiązania α,β-1,2-glikozydowego.Skrobia – węglowodan, polisacharyd roślinny, składający się wyłącznie z merów glukozy, pełniący w
roślinach rolę magazynu energii. Skrobia ma budowę ziarnistą.(C6H10O5)n n>300 Skrobia hydrolizuje wyłącznie na α-D-glukozę, składa się z dwóch różnych
polisacharydów:*nierozgałęzionej amylozy łatwiej rozpuszczalnej w wodzie (ok. 20% naturalnej skrobi); jest wielocukrem – jej cząsteczki składają się z wielu reszt
glukozowych połączonych ze sobą atomami tlenu (wiązania α -1,4 glikozydowe). *rozgałęzionej amylopektyny, nierozpuszczalnej w wodzie (ok. 80% naturalnej
skrobi); rozgałęzienia powstają dzięki wiązaniom α-1,6-glikozydowym. Glukoza -cukier skrobiowy, C6H12O6) – węglowodan należący do cukrów prostych. Glukoza
jest izomerem fruktozy - mają identyczny wzór sumaryczny, ale różne wzory strukturalne.Glukoza jest podstawowym związkiem energetycznym dla większości
organizmów, jest rozkładana w procesie glikolizy na kwas pirogronowy.
4Inicjacja transkrypcji u prokariontów polega na związaniu się polimerazy RNA z odpowiednim odcinkiem pasma matrycowego DNA - tzw. promotorem.
Polimeraza rozpoznaje sekwencje -35 i -10 promotora (a transkrypcja zaczyna się od nukleotydu +1). Specyficzność wiązania zapewnia czynnik σ (sigma). Rozsunięcie
nici DNA na odcinku kilkunastu nukleotydów (czyli powstanie tzw. kompleksu otwartego) umożliwia wstawianie (włączenie) kolejnych, odpowiednich nukleotydów.
Substratami są trifosforany rybonukleozydów (ATP, GTP, CTP i UTP). Transkrypcja zaczyna się od produkcji kilku krótkich (kilka nukleotydów) transkryptów. Dopiero
po oddysocjowaniu czynnika σ może rozpocząć się kolejny etap - elongacja transkrypcji (wydłużanie RNA). Polimeraza RNA przesuwa się systematycznie wzdłuż
helisy DNA, rozplatając ją (na odcinku kilkunastu par zasad) i wydłużając łańcuch RNA, przy czym nukleotydy włączane są zgodnie z zasadą komplementarności.
Powyżej aktualnego miejsca syntezy powstający hybrydowy kompleks DNA - RNA ulega rozpadowi, DNA powraca do swojej pierwotnej dwuniciowej struktury, a
łańcuch powstającego mRNA oddziela się. Etap elongacji kończy się, gdy polimeraza RNA dotrze do terminatora - sekwencji kończącej, wyznaczającej miejsce
terminacji (zakończenia) transkrypcji. Sekwencja taka tworzy strukturę szpilki do włosów (hairpin), która zatrzymuje polimerazę RNA, co powoduje rozpad kompleksu
enzym - DNA - RNA
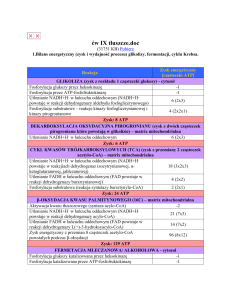
Kluczowe etapy utleniania (oddychania) komórkowego
•
•
•
Tworzenie acetylo – koenzymu A (np. glikoliza)
Obróbka acetylo – koenzymu A (cykl Krebsa)
Utlenianie produktów obróbki acetylo – koenzymu A (mitochondrialny łańcuch oddechowy)
•
Synteza ATP
Acetylo – koenzym A to produkt częściowego utleniania różnych monomerów
Glikoliza
•W wyniku glikolizy glukoza zostaje częściowo utleniona; powstają: ATP, pirogronian i NADH
•NADH musi być utleniany (regenerowany) do NAD + aby glikoliza mogla zachodzić. Temu służy fermentacja oraz pośrednie
utlenianie przez mitochondria (zad. domowe!)
•Pirogronian po opuszczeniu cytoplazmy (gdzie powstaje) i wejściu do mitochondrium jest przekształcany do acetylo – koenzymu A.
Cykl krebsa
W cyklu Krebsa dochodzi do dalszego utleniania fragmentów cząsteczki glukozy wprowadzonych w postaci grupy acetylowej acetylo
– koenzymu A. Skutkiem tego powstają: NADH, FADH2 i GTP.
•NADH i FADH2 to substraty mitochondrialnego łańcucha oddechowego
Plik z chomika:
tospg2012
Inne pliki z tego folderu:
Bichem 1 i 2.docx (401 KB)
bioch1.docx (18 KB)
bioch2.doc (72 KB)
cd do koła 1 moje.pdf (721 KB)
koło 1 MOJE.doc (2880 KB)
Inne foldery tego chomika:
Biochemia
Laboratorium
Wykład
Zgłoś jeśli naruszono regulamin
Strona główna
Aktualności
Kontakt
Dział Pomocy
Opinie
Regulamin serwisu
Polityka prywatności
Copyright © 2012 Chomikuj.pl