2016-03-10
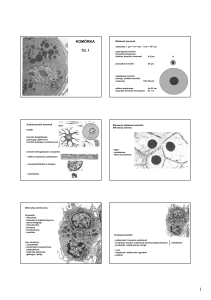
Wielkość komórek
Jednostki: 1 μm = 10-3 mm, 1 nm = 10-3 μm
KOMÓRKA
najmniejsze komórki
(komórki przytarczyc,
niektóre komórki nerwowe)
4-5 μm
przeciętne komórki
20 μm
największe komórki
(oocyty, niektóre komórki
nerwowe)
100-150 μm
włókna mięśniowe
wypustki komórek nerwowych
Zróżnicowanie komórek
kształt
•
do 30 cm
do 1 m
Elementy składowe komórki:
Mikroskop świetlny:
komórki bezjądrzaste:
erytrocyty, płytki krwi,
komórki budujące soczewkę oka
komórki wielojądrzaste i zespólnie:
• włókna mięśniowe szkieletowe
• jądro
• cytoplazma
• błona komórkowa
• syncytiotrofoblast w łożysku
• osteoklasty
Mikroskop elektronowy:
Organelle:
• rybosomy
• siateczka śródplazmatyczna
• aparat Golgiego
• mitochondria
• lizosomy
• peroksysomy
• centriole
Inne struktury:
• cytoszkielet
• pęcherzyki (transportowe,
wydzielnicze)
• materiały zapasowe
(glikogen, lipidy)
Błona biologiczna - uniwersalny budulec większości organelli
komórkowych
Błona biologiczna zbudowana jest z lipidów (głównie fosfolipidów) i białek.
Cząsteczka fosfolipidu
część hydrofilna
część hydrofobowa
W środowisku wodnym cząsteczki
fosfolipidów tworzą dwuwarstwę
o uporządkowanym układzie.
Decyduje ona o integralności błony
1
2016-03-10
Lipidy błon biologicznych:
białka
Białka transbłonowe
• fosfolipidy (aminowe i cholinowe)
• cholesterol
• glikolipidy
Białka błon biologicznych:
klasyfikacja
strukturalna:
• powierzchniowe
• transbłonowe (integralne)
Białka swobodnie „pływają” w dwuwarstwie lipidowej
Białka powierzchniowe
czynnościowa:
• strukturalne
• enzymatyczne
• receptorowe
• transportowe
Jedno białko może pełnić kilka funkcji
Błona biologiczna jest barierą dla substancji chemicznych i kontroluje
ich transport pomiędzy środowiskiem a komórką i pomiędzy przedziałami
wewnątrzkomórkowymi.
Mechanizm otwierania kanałów
Transport substancji niskocząsteczkowych (transbłonowy)
- poprzez zawarte w błonie transbłonowe białka transportowe
1. Kanały
zamknięty
otwarty
Transport bierny (dyfuzja): zgodnie z gradientem stężeń,
bez nakładu energii
Substancje transportowane: jony
Przykłady: kanały sodowe, potasowe, wapniowe, chlorkowe
2. Przenośniki
zmiana konformacji
Transport ułatwiony:
zgodnie z gradientem stężeń,
bez nakładu energii
• kanały otwierane zmianą potencjału elektrycznego błony
• kanały otwierane ligandem (przyłączeniem cząsteczki sygnałowej)
• kanały otwierane mechanicznie
Przenośniki i pompy mogą transportować jedną substancję (uniport)
albo równocześnie dwie substancje (kotransport)
Substancje transportowane:
aminokwasy, cukry proste,
nukleotydy, itp.
3. Pompy
zmiana konformacji
Transport aktywny:
wbrew gradientowi stężeń,
konieczna energia
Substancje transportowane:
wszystkie substancje
niskocząsteczkowe
Przykłady: pompa wapniowa,
pompa protonowa
uniport
symport
antyport
Przykłady:
• Na+-K+ ATPaza (pompa sodowo-potasowa)
• H+-K+ ATPaza (wymiennik protonowo-potasowy)
• symporter Na+-glukoza
2
2016-03-10
Transport substancji wysokocząsteczkowych i dużych struktur:
transport pęcherzykowy
Błona komórkowa
oddziela komórkę od środowiska zewnętrznego
błona
komórkowa
Równocześnie transportowana jest substancja/struktura
oraz fragment błony
Glikokaliks
glikokaliks
• najgrubsza
• trójwarstwowy obraz w mikroskopie elektronowym
• warstwa cukrowców na powierzchni (glikokaliks)
• specyficzne glikoproteidy odpowiedzialne za kontakt z innymi
komórkami i z substancją międzykomórkową (cząsteczki adhezyjne)
• liczne białka transportowe i receptory
• nierównomierne rozmieszczenie ładunków elektrycznych
po obu stronach błony (potencjał spoczynkowy)
Cząsteczki adhezyjne
kadheryny
Glikoproteidy odpowiedzialne za wiązanie się
komórek z otoczeniem (innymi komórkami
i substancją międzykomórkową)
• kadheryny - wiązanie komórka-komórka
• selektyny - wiązanie komórka-komórka
• białka z nadrodziny immunoglobulin - wiązanie komórka-komórka
• integryny - wiązanie komórka-substancja międzykomórkowa
Warstwa cukrowcowa na zewnętrznej powierzchni błony komórkowej
zbudowana z łańcuchów cukrowcowych połączonych z cząsteczkami
białek (glikoproteidów) i lipidów (glikolipidów)
Funkcje:
• ochrona komórki przed czynnikami mechanicznymi i chemicznymi
• udział w regulacji pobierania substancji przez komórkę (endocytozy)
• udział w kontaktowym „rozpoznawaniu się” komórek
• pośredniczenie w tworzeniu połączeń komórek ze sobą i z otoczeniem
Receptory
i sygnalizacja
międzykomórkowa
Sygnalizacja
międzykomórkowa
jest konieczna dla:
• normalnego
funkcjonowania
komórki
• podziałów
• różnicowania
• przeżycia
Substancje sygnałowe:
• hormony
• neuroprzekaźniki
• czynniki wzrostowe
• cytokiny
FUNKCJONOWANIE
integryna
elementy substancji
międzykomórkowej
Komórka reaguje na sygnał
jeżeli posiada specyficzne receptory
dla cząsteczek sygnałowych
błona komórkowa
PODZIAŁ
receptor
RÓŻNICOWANIE
cząsteczka sygnałowa
ŚMIERĆ
Cząsteczkę rozpoznawaną i wiązaną przez receptor nazywamy ogólnie ligandem
3
2016-03-10
Reakcja komórki zależy od jej możliwości
i od rodzaju receptorów (różne receptory wywołują różne reakcje
komórki na tę samą substancję sygnałową)
sygnalizacja endokrynna (dokrewna)
sygnalizacja autokrynna
sygnalizacja parakrynna
przekaźnictwo nerwowe
sygnalizacja
kontaktowa
Jądro komórkowe
Lokalizacja receptorów zależy od tego, czy cząsteczki sygnałowe
mogą dostać się do wnętrza komórki
Funkcje:
• magazynuje informację genetyczną
(DNA)
• powiela informację genetyczną
(replikacja DNA) przed podziałem
komórki
• wytwarza rybosomy (jąderko)
• steruje syntezą białek w komórce,
kierując w ten sposób wszystkimi
procesami życiowymi
DNA
RNA
transkrypcja
Typy chromatyny:
• euchromatyna (jasna, luźna,
aktywna transkrypcyjnie)
• heterochromatyna (ciemna, zwarta,
nieaktywna transkrypcyjnie)
- konstytutywna (stała dla wszystkich
komórek, DNA niekodujący)
- fakultatywna (zależna od typu
komórki, wynik „wyłączania” genów
w trakcie różnicowania)
białko
Główne składniki:
• chromatyna
• jąderko
• otoczka jądrowa
• receptory błonowe (powierzchniowe)
• receptory wewnątrzkomórkowe
(np. jądrowe, mitochondrialne)
Chemiczne składniki chromatyny:
• DNA
• białka
- histony (H1, H2A, H2B, H3, H4)
- białka niehistonowe (enzymatyczne,
regulatorowe, strukturalne)
translacja
Budowa chromatyny
Podjednostka: nukleosom
• rdzeń - oktamer histonów
(2 X H2A, H2B, H3, H4)
• nawinięty fragment DNA
(146 par zasad)
Odcinek DNA łączący
nukleosomy: 80 par zasad
+ histon H1
4
2016-03-10
Budowa chromatyny c.d.
DNA
Jąderko
nukleosom
produkuje podjednostki rybosomów
nukleofilament
euchromatyna
włókno chromatynowe
1
2
--------------------------------------------------------tworzenie bocznych pętli
heterochromatyna
superspirala
--------------------------------------------------------podział
chromosom
Otoczka jądrowa
kontroluje wymianę substancji pomiędzy jądrem
a cytoplazmą przez znajdujące się w niej pory
Obszary jąderka:
1 - jasne centra włókienkowe - nieaktywny rDNA
2 - gęste obszary włókienkowe - pre-rRNA
3 - obszary ziarniste - podjednostki rybosomów
Etapy produkcji podjednostek rybosomów:
• transkrypcja rDNA pre-rRNA
• cięcie pre-rRNA na mniejsze fragmenty rRNA
• przyłączanie białek importowanych z cytoplazmy podjednostki rybosomów
Kompleks poru jądrowego
Białka kompleksu:
• receptory
• białka transportowe
cytoplazma
białka
transportowe
Zbudowana z dwóch błon:
• błona wewnętrzna z blaszką jądrową (laminy A,B,C)
• błona zewnętrzna - kontynuacja błon siateczki
śródplazmatycznej
Pomiędzy błonami przestrzeń okołojądrowa (do 100 nm)
Otoczka jądrowa uważana jest za szczególny obszar
siateczki śródplazmatycznej
3
jądro
Transport
jądrowo-cytoplazmatyczny:
• z jądra do cytoplazmy:
mRNA, tRNA, podjednostki
rybosomów
• z cytoplazmy do jądra:
wszystkie białka jądrowe
Jak działa kompleks poru:
• małe cząsteczki przechodzą swobodnie
• duże cząsteczki są rozpoznawane przez receptory
• a następnie przenoszone przez białka transportowe
Rybosomy - syntetyzują białka
- rybosomy związane z błonami siateczki szorstkiej produkują:
białka błon biologicznych, białka wydzielnicze, białka lizosomowe
- rybosomy wolne (cytoplazmatyczne) produkują:
białka jądrowe, mitochondriów, peroksysomów, cytoszkieletu, cytoplazmy
49 białek
33 białka
Podjednostki rybosomów łączą się
w cytoplazmie dopiero po przyłączeniu
mRNA
Polirybosomy (polisomy):
rybosomy połączone nicią mRNA
(w trakcie translacji)
Docelowa lokalizacja białka zależy od tzw. odcinka sygnałowego: krótkiego
fragmentu łańcucha polipeptydowego zlokalizowanego zazwyczaj na jednym
z końców cząsteczki białkowej
5
2016-03-10
rybosom
odcinek
sygnałowy
Biosynteza białka (na przykładzie białka wbudowywanego
do siateczki śródplazmatycznej)
Białka mają odmienne odcinki sygnałowe
umożliwiające wbudowanie ich do:
• siateczki śródplazmatycznej
• jądra
• mitochondriów
• peroksysomów
• cytoszkieletu
Odcinki sygnałowe są rozpoznawane przez odpowiednie receptory
znajdujące się w docelowych organellach. Po związaniu odcinka sygnałowego
z receptorem, białko zostaje wbudowane do organelli:
• do jądra - przechodzi przez por w otoczce jądrowej
• do siateczki, mitochondriów i peroksysomów przechodzi przez specjalne „kanały białkowe”,
translokony, obecne w ich błonach
Białko może mieć także odcinek „stop” - zazwyczaj
wewnątrz cząsteczki - powoduje on zatrzymanie białka
w błonie (w ten sposób wbudowywane są białka błon).
Do siateczki białka wbudowywane są podczas translacji,
do innych organelli po zakończeniu translacji
SRP
receptor
i translokon
1. Przyłączenie mRNA do małej podjednostki rybosomu.
2. Przyłączenie dużej podjednostki do małej - tworzy się kompletny rybosom.
3. Rozpoczęcie translacji - jako pierwszy powstaje odcinek sygnałowy
4. Przyłączenie „cząsteczki rozpoznającej sygnał” - SRP - do odcinka sygnałowego
5. Przyłączenie kompleksu SRP-odcinek sygnałowy do receptora SRP w błonie
siateczki śródplazmatycznej
Siateczka śródplazmatyczna
6. Skierowanie odcinka sygnałowego do translokonu w błonie siateczki - białko
zaczyna przechodzić przez błonę.
7. Przyłączenie dużej podjednostki rybosomu do błony siateczki przy udziale
białek mocujących (ryboforyn) obecnych w błonie.
8. Dalsze przechodzenie białka przez błonę, odcięcie odcinka sygnałowego.
9. Jeżeli białko nie ma odcinka „stop”, przechodzi w całości przez błonę
i wewnątrz siateczki ulega sfałdowaniu (przyjmuje strukturę trzeciorzędową).
10. Jeżeli białko ma odcinek „stop”, zostaje wbudowane w błonę siateczki
i tam się fałduje.
11. Po ukończeniu translacji rybosom rozpada się na oddzielne podjednostki.
Aparat Golgiego
szorstka
• spłaszczone cysterny
• rybosomy
• receptory dla SRP, translokony, ryboforyny
• funkcje:
- synteza białek
- wstępna glikozylacja białek
Bieguny diktiosomu
CIS
- wypukły
- błona (5 nm) podobna
do błon siateczki
Diktiosom - podjednostka
aparatu Golgiego
gładka
• kanaliki
• brak rybosomów
• funkcje:
- produkcja lipidów
- neutralizacja leków i trucizn
- gromadzenie jonów Ca2+
(kalciosomy)
TRANS
- wklęsły
- błona (7,5 nm) podobna
do błony komórkowej
- wewnętrzna powierzchnia
błony bogata w cukry
Przez diktiosom „przepływają” (transport pęcherzykowy) fragmenty błony
i białka z siateczki śródplazmatycznej. W trakcie przepływu od bieguna cis
do trans błona i transportowane białka ulegają modyfikacji chemicznej.
Błona stopniowo zmienia charakter na podobny do błony komórkowej.
6
2016-03-10
Jak działa diktiosom
• małe pęcherzyki oddzielają się od siateczki i łączą z błoną diktiosomu
po stronie cis
• podczas przechodzenia przez kolejne cysterny diktiosomu, do białek błony
i białek transportowanych wewnątrz pęcherzyków zostają przyłączane grupy
cukrowcowe (glikozylacja), niekiedy również fosforanowe i siarczanowe
• w ostatniej cysternie po stronie trans białka są sortowane i „pakowane”
do różnych pęcherzyków, które oddzielają się od tej cysterny
Proces oddzielania pęcherzyków wymaga współdziałania
tzw. białek okrywających (COP I, COP II, klatryna)
Z diktiosomu „wypączkowują” trzy rodzaje pęcherzyków:
• małe pęcherzyki z materiałem do wydzielenia na drodze egzocytozy
konstytutywnej
• duże pęcherzyki z materiałem do wydzielenia na drodze egzocytozy
regulowanej
• pęcherzyki hydrolazowe zawierające enzymy trawienne prekursory lizosomów
Egzocytoza
Funkcje aparatu Golgiego:
• przebudowa błon i odnowa błony komórkowej
• glikozylacja przepływających białek, ich sortowanie i kierowanie
do różnych pęcherzyków
• tworzenie pęcherzyków i ziarn wydzielniczych - udział w procesie wydzielania
• tworzenie pęcherzyków hydrolazowych
• transport pęcherzyków zawierających substancje do wydzielenia
• fuzja pęcherzyków z błoną komórkową - wydzielenie substancji
Endocytoza:
Dwa rodzaje egzocytozy:
• fagocytoza
• konstytutywna
(ciągła, wolna,
małe pęcherzyki)
sygnał
• pinocytoza
• endocytoza receptorowa
• regulowana
(na sygnał, np.
nerwowy lub
hormonalny),
szybka,
duże ziarna
wydzielnicze
7
2016-03-10
Fagocytoza
Pinocytoza
Endocytoza
receptorowa:
etapy procesu
wczesny endosom
późny endosom
• pobierane duże cząstki stałe
• błona komórkowa „wspina
się” otaczając cząstkę
• udział cytoszkieletu
• potrzebna energia
• powstają duże pęcherzyki
(fagosomy)
• pobierany płyn
• błona komórkowa wpukla się
• bez udziału cytoszkieletu
• bez udziału energii
• powstają małe pęcherzyki
(pinosomy)
1. Przyłączenie ligandów do receptorów na powierzchni błony komórkowej
2. Podbłonowa agregacja klatryny, skupienie receptorów i utworzenie
dołeczka okrytego
3. Endocytoza - utworzenie pęcherzyka okrytego
4. Oddzielenie klatryny od pęcherzyka
5. Połączenie pęcherzyka z wczesnym endosomem
Endocytoza
receptorowa:
etapy procesu c.d.
wczesny endosom
późny endosom
6. We wczesnym endosomie: niskie pH (pompa protonowa), oddzielenie
ligandów od receptorów, powrót receptorów transportem pęcherzykowym
do błony komórkowej (recyrkulacja receptorów).
7. Przemieszczenie endosomu w głąb komórki - staje się późnym endosomem
8. W późnym endosomie: przyłączenie pęcherzyków hydrolazowych
zawierających enzymy trawienne - późny endosom przekształca się
w lizosom
9. Trawienie ligandów
Lizosomy
pęcherzyki, w których zachodzi trawienie wewnątrzkomórkowe;
zawierają enzymy trawienne (hydrolazy) i trawione substancje
lizosom
Lizosomy powstają przez połączenie (fuzję) pęcherzyków hydrolazowych
zawierających enzymy trawienne z pęcherzykami zawierającymi substancje,
które mają zostać strawione
Powstawanie pęcherzyków
hydrolazowych
1. Synteza enzymów trawiennych
(hydrolaz) w siateczce szorstkiej
2. Transport pęcherzykowy
enzymów do diktiosomu a. G.
3. W diktiosomie do enzymów
przyłączane są grupy
mannozo-6-fosforanu (znacznik)
4. Znacznik rozpoznawany jest
i wiązany przez receptory
w błonie ostatniej cysterny
diktiosomu po stronie trans
5. Od tej cysterny odrywa się
pęcherzyk okryty zawierający
hydrolazy związane z receptorami.
Hydrolazy (esterazy, glikozydazy, peptydazy) mogą trawić wszystkie rodzaje
substancji wysokocząsteczkowych.
Hydrolazy w pęcherzykach hydrolazowych są nieaktywne, gdyż związane
są z receptorami.
8
2016-03-10
(2) z pęcherzykami powstałymi w komórce, zawierającymi jej własne struktury
(autofagosomami) - powstają autolizosomy
fagocytoza
LIZOSOM
pinocytoza,
endocytoza receptorowa
LIZOSOM
Pęcherzyki hydrolazowe mogą się łączyć:
(1) z pęcherzykami powstałymi w wyniku endocytozy:
• z fagosomami
• z pinosomami
• z późnymi endosomami
- powstają heterolizosomy
Wewnątrz lizosomów panuje niskie pH, w wyniku czego hydrolazy
oddzielają się od receptorów, stają się aktywne i trawią zawarte
w lizosomach substancje.
Produkty trawienia (substancje niskocząsteczkowe) przechodzą przez
błonę lizosomu do cytoplazmy, gdzie mogą służyć do syntezy nowych
substancji użytecznych dla komórki, lub do produkcji energii.
Receptory dla mannozo-6-fosforanu powracają transportem
pęcherzykowym do diktiosomu.
lizosomy
Proteasomy
Białka mogą być również trawione poza lizosomami, przez znajdujące się
w cytoplazmie kompleksy enzymatyczne - proteasomy
Błona
komórkowa
endosomy
siateczka
śródplazmatyczna
pęcherzyki
hydrolazowe
fagosomy,
pinosomy,
pęcherzyki
okryte
pęcherzyki wydzielnicze
Rozpoznawane i trawione przez proteasomy są tylko te białka, które
zostały „oznakowane” przez przyłączenie do nich innego białka ubikwityny
aparat Golgiego
Transport pęcherzykowy pomiędzy organellami i błoną komórkową
określamy jako przepływ błon w komórce
Mitochondria i peroksysomy nie uczestniczą w przepływie błon, zatem
(1) namnażają się przez podział i nie mogą powstać de novo, (2) ich
błony mają unikatowy charakter i (3) ich białka są syntetyzowane na
wolnych rybosomach i posttranslacyjnie wbudowywane do organelli
MITOCHONDRIA
Przedziały
mitochondrialne
•
•
•
•
• błona zewnętrzna
• błona wewnętrzna (tworzy grzebienie lamelarne lub tubularne)
• przestrzeń międzybłonowa
• macierz
9
2016-03-10
Błona wewnętrzna:
Błona zewnętrzna:
• białka/lipidy 1:1
• poryny - nieselektywna przepuszczalność < 5 kDa
• translokony dla importu białek (TOM)
• białka/lipidy 4:1
• specyficzny lipid (kardiolipina)
• liczne białka transportowe (w pełni kontrolowany transport)
• translokony dla importu białek (TIM)
• łańcuch przenośników elektronów
• „grzybki mitochondrialne” - kompleks syntazy ATP
Przestrzeń międzybłonowa:
Macierz:
• kinazy nukleotydów
• miejsca kontaktowe (styk obu błon i translokonów)
• aparat genetyczny (mtDNA, mtRNA, mt-rybosomy)
• enzymy cyklu Krebsa
• enzymy β-oksydacji kwasów tłuszczowych
• ciałka gęste (złogi fosforanów wapnia)
Główna funkcja mitochondriów, produkcja ATP wymaga
współdziałania (1) enzymów cyklu Krebsa, (2) łańcucha
przenośników elektronów i (3) syntazy ATP
Trzy składniki łańcucha oddechowego „pompują” protony
z macierzy do przestrzeni międzybłonowej. Powstały gradient
protonowy służy jako źródło energii dla syntezy ATP w „grzybkach”
(a także dla aktywnego transportu przez błonę wewnętrzną oraz dla
importu białek)
3
2
przestrzeń
międzybłonowa
błona
wewnętrzna
1
macierz
dehydrogenaza
NADH
cytochromy
b – c1
oksydaza
cytochromowa
syntaza ATP
Mitchell, Nagroda Nobla 1978
Grzybek mitochondrialny (F0-F1 ATPaza) składa się z „nóżki” (F0)
zawierającej transporter protonowy i „główki” (F1) - syntazy ATP.
Grzybek działa jak „turbina molekularna”
transporter
protonowy
Inne funkcje mitochondriów:
F0
• β-oksydacja kwasów tłuszczowych
• ostatnie etapy syntezy hormonów sterydowych
• udział w regulacji poziomu Ca2+ w komórce
• produkcja ciepła
• udział w procesie apoptozy
2000 obr/min Boyer & Walker, Nagroda Nobla 1997
10
2016-03-10
Mitochondria zawierają własny aparat genetyczny:
• pętlowy mt DNA
• wszystkie rodzaje mtRNA
• rybosomy
• enzymy niezbędne do procesów replikacji,
transkrypcji i translacji
... Jednak jest on zbyt ubogi, aby zapewnić organelli pełną autonomię
podjednostki
syntazy ATP
podjednostki oksydazy
cytochromowej
podjednostki
dehydrogenazy NADH
długość genomu: 16 569 par zasad
podjednostki
dehydrogenazy NADH
mtDNA koduje:
• 12S i 16S rRNA do rybosomów mitochondrialnych
• 22 cząsteczki tRNA
• 13 białek błony wewnętrznej
pozostałe białka mitochondrialne są kodowane w DNA jądrowym,
syntetyzowane w cytoplazmie na wolnych rybosomach
i po translacji wbudowywane do mitochondriów
Mitochondria ewolucyjnie wywodzą się z prymitywnych bakterii, które
posiadały zdolność do produkcji ATP (teoria endosymbiotyczna)
pierwotna
bakteria
Peroksysomy mają przeważnie formę pęcherzyków, ale
w niektórych komórkach mogą być różnokształtne
pierwotna komórka
eukariotyczna
FAGOCYTOZA (?)
SYMBIOZA, TRANSFER GENÓW
mitochondrium
jądro
Cytoszkielet
Enzymy peroksysomowe:
• oksydazy peroksysomowe
• katalaza
• enzymy ß-oksydacji kwasów tłuszczowych
• enzymy biosyntezy lipidów
• aminotransferazy
Typ włókien
Średnica
Białko
Funkcja
mikrotubule
25 nm
tubulina
ruch,
podporowa
mikrofilamenty
6 nm
aktyna
ruch,
podporowa
10 nm
różne
białka
podporowa
Główne funkcje peroksysomów:
• utlenianie różnych substratów, w tym detoksyfikacja
• rozkład nadtlenku wodoru
• ß-oksydacja długołańcuchowych kw. tłuszczowych
• synteza cholesterolu, kwasów żółciowych
i eterolipidów (plazmalogenów)
• degradacja puryn
filamenty
pośrednie
11
2016-03-10
Mikrotubule nietrwałe (cytoplazmatyczne)
Mikrotubule trwałe (po zakończeniu wzrostu nie rozpadają się):
• neurotubule w wypustkach komórek nerwowych
• mikrotubule budujące złożone struktury: rzęski, witki i centriole
Mikrotubule
tubulina
protofilament
rureczki
zbudowane z tubuliny
-
+
W strukturach mikrotubule łączą się
w dublety lub triplety
Dynamiczne (wydłużają się
i skracają)
koniec „+” - wydłużanie
koniec „-” stabilny, zazwyczaj
zakotwiczony w pobliżu centrioli
Aksonema rzęsek i witek:
9 obwodowych dubletów
i 2 mikrotubule centralne
Centriola: 9 obwodowych tripletów, para centrioli = centrosom
Mikrofilamenty (filamenty aktynowe)
zbudowane z aktyny
--
Centriole i otaczający je materiał
zawierający tubulinę G
indukują i regulują wzrost mikrotubul
cytoplazmatycznych
(centrum organizacji mikrotubul)
Za zjawiska ruchu komórkowego
odpowiedzialne są tzw. mechanoenzymy
(białka motoryczne), które wykorzystując
energię z ATP „kroczą” po powierzchni
mikrotubul i mikrofilamentów
E
+
• nietrwałe
• twałe (w połączeniach międzykomórkowych, w niektórych komórkach
nabłonkowych - sieć krańcowa, w kom. mięśniowych)
dyneina
Po powierzchni mikrotubul mogą
„kroczyć” dwa mechanoenzymy:
• dyneina - w kierunku końca „-”
• kinezyna - w kierunku końca „+”
kinezyna
+
mikrotubula
mikrotubula
Do mechanoenzymów mogą się
przyczepiać różne struktury, które są
w ten sposób transportowane wzdłuż
mikrotubul, „jak po szynach”:
• pęcherzyki
• organelle (ruch organelli)
• chromosomy
• duże białka
E
12
2016-03-10
Po powierzchni filamentów aktynowych może kroczyć tylko jeden
rodzaj mechanoenzymu: miozyna, wyłącznie w kierunku końca „+”.
Filamenty aktynowe zakotwiczają się tym końcem w błonie komórkowej,
a układ ten odpowiada za zjawiska ruchowe, w których uczestniczy błona:
tworzenie wpukleń i fałdów
błony, wysuwanie i wciąganie
wypustek (fagocytoza,
ruch pełzakowaty): miozyna I
Filamenty pośrednie
• zbudowane z łańcuchów białkowych
skręconych w formę liny (wytrzymałe
elastyczne)
• nie współpracują z mechanoenzymami,
pełnią wyłącznie funkcje podporowe
(wewnątrz komórki i w połączeniach
międzykomórkowych)
• są zbudowane z różnych białek,
zależnie od miejsca występowania
skurcz komórki: (np. komórki
mięśniowe): miozyna II
(agreguje w filamenty
miozynowe)
Materiały zapasowe i wtręty cytoplazmatyczne
Nazwa
Białka budujące
Występowanie
laminy jądrowe
laminy A i B
jądra wszystkich komórek
filamenty keratynowe
cytokeratyny
komórki nabłonkowe
filamenty wimentynowe
wimentyna
komórki tk. łącznej
filamenty desminowe
desmina
komórki mięśniowe
filamenty glejowe
GFAP (kwaśne
włókienkowe
białko glejowe)
komórki neurogleju
neurofilamenty
białka
neurofilamentów
komórki nerwowe
glikogen
wtręty krystaliczne
i parakrystaliczne
ziarna barwnika
krople lipidowe
nekroza
apoptoza
Śmierć komórki:
• martwica (nekroza)
• apoptoza (zaprogramowana śmierć)
Martwica:
- przerwanie błony komórkowej
- zahamowanie procesów życiowych
- rozpad komórki
- odczyn zapalny
Faza indukcji:
• aktywacja receptorów śmierci lub produkcja
specyficznych białek indukujących apoptozę
• uwalnianie cytochromu c z mitochondriów
fragmentacja DNA
Faza egzekucji:
• aktywacja kaspaz (enzymów proteolitycznych)
• trawienie białek wewnątrzkomórkowych
• zaburzenie procesów metabolicznych
• śmierć komórki
Apoptoza:
- aktywacja kolejnych genów (program)
- produkcja i aktywacja szczególnych białek
Czynniki wywołujące apoptozę:
• uszkodzenie DNA
• określone cząsteczki sygnałowe
działające na tzw. „receptory śmierci”
• brak składników odżywczych lub
czynników wzrostowych
„pączkowanie” błony
Morfologiczne cechy apoptozy:
• fragmentacja DNA
• rozpad jądra na kilka fragmentów
• zagęszczenie cytoplazmy
• rozpad komórki na małe fragmenty (ciałka
apoptotyczne) otoczone błoną
tworzenie ciałek apoptotycznych
13