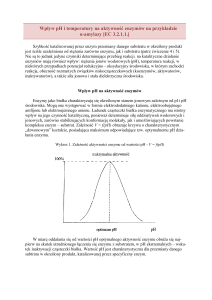
Badanie termostabilności oraz wpływu aktywatorów i inhibitorów na działanie α-amylazy [EC 3.2.1.1.]
Termostabilność enzymów
Dla większości enzymów zmiany denaturacyjne zachodzą bardzo intensywnie powyżej 40 – 50 °C, jednakże istnieją enzymy wykazujące znaczną odporność na inaktywujące
działanie podwyższonej temperatury – cechuje je wysoka termostabilność. Termostabilność
jest jedną z cech charakterystycznych każdego enzymu, może być wyższa bądź niższa. Wyrażamy ja podając najwyższą temperaturę, przy której nie dochodzi w określonym czasie do
termicznej inaktywacji enzymu, poddawanego działaniu tej temperatury w nieobecności substratu. Termostabilność białka enzymatycznego jest zależna od wielu czynników, takich jak
pH i siła jonowa środowiska, a także obecności w nim wielorakich substancji niskocząsteczkowych. Niektóre enzymy muszą wykazywać wysoką tolerancję na wysoką lub niską temperaturę chociażby z powodu środowiska życia niektórych organizmów, szczególnie bakterii.
Aktywatory i inhibitory enzymów
Szybkość reakcji enzymatycznych zależy najczęściej od obecności w układzie substancji innych niż enzym i substrat. Mogą one przyspieszyć (aktywatory) bądź opóźnić (inhibitory) katalityczne działanie enzymu.
Do aktywatorów enzymów zaliczamy:
jony niektórych metali, zwłaszcza dwuwartościowych (najczęściej Mg2+, Zn2+, Mn2+,
Ca2+, Fe2+, Fe3+, rzadziej Cu2+, Co2+, Ni2+, K+, Na+). Jon metalu może być zlokalizowany w katalitycznym centrum enzymu (bierze wówczas bezpośredni udział w reakcji) lub w innym fragmencie molekuły stabilizując jej aktywną konformację;
substancje białkowe, aktywujące grupy czynne enzymów, np. biorące udział w aktywacji proenzymu, kowalencyjnej modyfikacji nieaktywnej formy enzymu;
niskocząsteczkowe związki organiczne, znoszące działanie inhibitorów enzymów.
aniony wywierają niewielki wpływ na aktywność enzymów. Do wyjątków należy aktywacja α-amylazy przez jony Cl-.
Inhibitory enzymów mogą mieć charakter specyficzny lub niespecyficzny. Wśród inhibitorów
specyficznych, a więc obniżających aktywność ściśle określonych enzymów wyróżniamy
związki:
współdziałające z enzymem w tym samym obszarze cząsteczki, co substrat – łączące
się z centrum aktywnym. Są one strukturalnie podobne do substratu, łączą się z enzymem odwracalnie. Nazywamy je inhibitorami kompetencyjnymi (współzawodniczącymi);
wiążące się z enzymem w innym obszarze cząsteczki niż substrat zwanym centrum allosterycznym. Inhibitory tego typu, zwane niekomopetycyjnymi mogą się łączyć albo
z wolnym enzymem, albo z kompleksem enzym – substrat (w tym przypadku inhibicja
ma zwykle charakter nieodwracalny)
Niespecyficznymi inhibitorami enzymów są jony metali ciężkich (Cu, Pb, Hg, As, Ag). Wiążą się one łatwo i w sposób nieodwracalny ze wszystkimi białkami, powodując zmiany ich
konformacji prowadzące do denaturacji, której często towarzyszy wypadanie białka w postaci
osadu. Szczególnie podatne na wiązanie jonów metali ciężkich są grupy sulfhydrylowe (-SH).
Metal może się również wbudować w mostek disiarczkowy. Jon metalu ciężkiego może przy-
łączać się zarówno w centrum katalitycznym enzymu, jak i w innym obszarze cząsteczki.
Prowadzi to do całkowitej i najczęściej nieodwracalnej utraty aktywności przez enzym. Tylko
w niewielu przypadkach można cofnąć inaktywację stosując specjalne substancje konkurujące
z białkiem o jon metalu ciężkiego (np. 2,3-dimerkaptopropanol).
Właściwości aktywujące lub inhibitujące względem biokatalizatorów wykazuje również szereg związków organicznych. Niektóre z nich działają wybiórczo na wybrany enzym lub grupę
enzymów, dzięki czemu są stosowane do badań molekularnych struktury białek enzymatycznych.
Przebieg enzymatycznej amylolizy
α-Amylazy to szeroko rozpowszechnione w przyrodzie enzymy należące do klasy
hydrolaz i podklasy hydrolaz glikozydowych. Wytwarzają je liczne drobnoustroje oraz organizmy roślinne i zwierzęce. Ze względu na wartość produkcji i zapotrzebowanie, najważniejszymi przemysłowymi preparatami tego enzymu są α-amylazy bakteryjne, wytwarzane przez
szczepy z rodzaju Bacillus oraz roślinna α-amylazy ze słodu.
Skrobia jest wielkocząsteczkowym polimerem α-D-glukopiranozy. Znane są dwie
frakcje skrobi: nierozgałęziona amyloza, w której reszty glukozy połączone są wiązaniami
α-1,4-glikozydowymi w długie łańcuchy oraz amylopektyna, w której oprócz głównych wiązań α-1,4-glikozydowych występują także wiązania rozgałęziające α-1,6-glikozydowe.
α-Amylaza rozkłada tylko wewnętrzne wiązania α-1,4-glikozydowe, powodując stopniowe
rozszczepienie łańcuchów skrobi na coraz krótsze fragmenty.
Najpierw pod działaniem skrobi powstają nieco krótsze fragmenty – amylodekstryny nastepnie krótsze produkty hydrolizy – erytrodekstryny. α-Amylaza degraduje
powstające erytrodekstryny do achrodekstryn. W drugim etapie hydrolizy, achrodekstryny są
rozkładane przez enzym do glukozy, maltozy i mieszaniny oligocukrów redukujących.
Zasada oznaczania aktywności α-amylazy
W celu zaobserwowania przebiegu hydrolizy skrobi pod działaniem α-amylazy wykorzystuje się metody pozwalające na pomiar przyrostu stężenia cukrów redukujących uwalnianych z substratu przez enzym. Są to między innymi metody: Fehlinga, Somogyi - Nelsona,
oraz Hagedorna – Jensena, w których stężenia cukrów redukujących oznacza się na podstawie
ilości Cu2O powstającego na skutek redukcji jonów miedziowych przez wolne grupy redukujące cukrów, oraz metoda Millera z kwasem 3’,5’-dinitrosalicylowym (DNS) stosowana w
niniejszym ćwiczeniu.
W metodzie Millera aktywność enzymu oznacza się mierząc przyrost cukrów redukujących uwolnionych w czasie reakcji hydrolizy skrobi katalizowanej przez α-amylazę przebiegającej w przyjętych warunkach temperatury i pH. Cukry te oznacza się ilościowo stosując
alkaliczny roztwór kwasu 3’,5’-dinitrosalicylowego (DNS). W analogiczny sposób można
oznaczyć aktywność dowolnej hydrolazy glikozydowej, stosując odpowiedni dla niej substrat.
DNS nie tylko umożliwia oznaczenie stężenia cukrów redukujących, ale także pełni rolę inaktywatora enzymu, hamującego katalizowaną przez niego reakcję.
W środowisku alkalicznym i w temperaturze 100 °C DNS ulega redukcji pod działaniem cukrów redukujących, dając pochodną aminową, zabarwioną w środowisku alkalicznym na kolor pomarańczowy. Natężenie barwy, o maksimum absorbancji przy długości fali λ = 540 nm,
jest proporcjonalne do stężenia cukrów redukujących w badanym roztworze.
Stężenie cukrów redukujących w próbie odczytuje się z krzywej wzorcowej A540 = f(cmaltozy),
przygotowanej w warunkach oznaczenia.
Wykres 1. Krzywa wzorcowa. Zależność absorbancji przy λ = 540 nm od stężenia maltozy - A540 =
f(cmaltozy)
Literatura:
Stryer L., Biochemia, PWN, 2003
Sivak M.N, Preiss J;. Starch: Basic Science to Biotechnology. Advances in Food and Nutrition Research, vol. 41. New York: Academic
Press, 1998
Miller G.L.; Use of dinitriosalicylic acid reagent for determination of reducing sugar, Anal. Chem., 31, 426–428, 1959
ODCZYNNIKI-MATERIAŁY-SPRZĘT
1% koloidalny roztwór skrobi
1% alkaliczny roztwór kwasu 3’,5’-dinitrosalicylowego (DNS)
1% roztwór NaCl
1% roztwór CuSO4
woda destylowana
pipety automatyczne
termobloki: 37, 50 oraz 100 °C
spektrofotometr
stoper
PROCEDURY
Przygotowanie roztworu śliny
1. Do probówki 50 ml wprowadzić 5 ml śliny.
2. Następnie rozcieńczyć ślinę wodą destylowaną 1:1.
3. Tak przygotowany roztwór enzymu użyć do wykonania kolejnych punktów ćwiczenia.
Badanie termostabilności α-amylazy
1. Do 5 probówek odmierzyć po 0,25 ml roztworu śliny i po 0,25 ml buforu o pH 7. Zawartość probówek wymieszać. Równocześnie przygotować próby odczynnikowe dla
każdej temperatury zawierające (0,25 ml wody oraz 0,25 ml buforu o pH 7 , UWAGA: do prób odczynnikowych nie dodajemy śliny)
2. Probówki inkubować przez 10 minut w następujących temperaturach: -20 °C zamrażarka, pokojowa (stół laboratoryjny), 37, 60 oraz 95°C.
3. Po 10 minutach roztwór enzymu doprowadzić do temperatury pokojowej i do każdej
probówki dodać 0,5 ml koloidalnego roztworu skrobi (po uprzednim dokładnym wymieszaniu) i dokładnie wymieszać.
4. Probówki inkubować przez 20 minut w temperaturze 37°C , po czym pobrać 0,2 ml
mieszaniny reakcyjnej, wprowadzić do czystych probówek, do których dodać także po
0,2 ml roztworu DNS. Zawartość wymieszać i umieścić na 5 minut w termobloku 100
°C
5. Po 20 minutach probówki wyjąć, schłodzić do temperatury pokojowej pod bieżącą
wodą, dodać 0,6 ml wody destylowanej i dokładnie wymieszać.
6. Zmierzyć absorbancję roztworu przy λ=540 nm (A540) wobec próby odczynnikowej.
7. Z krzywej wzorcowej odczytać stężenie cukrów redukujących (mgmaltozy/ml). Wyniki
umieścić w tabeli.
8. Na podstawie tabeli sporządzić wykres zależności stężenia cukrów redukujących
uwolnionych przez α-amylazę ze skrobi od temperatury preinkubacji enzymu.
9. Z wykresu odczytać , w jakim zakresie temperatur 10-minutowa preinkubacja nie
spowodowała spadku aktywności α-amylazy.
Wpływ aktywatorów i inhibitorów na α-amylazę.
1. Do 3 probówek odmierzyć po 0,25 ml roztworu śliny, 0,25 ml buforu o pH 7. Następnie dodać do próbówek po 0,1 ml aktywatorów/inihibitorów (CuSO4, NaCl oraz wodę). Zawartość probówek wymieszać. Równocześnie przygotować próby odczynnikowe dla każdego ze związków zawierające (0,25 ml wody, 0,25 ml buforu o pH 7
oraz po 0,1 ml każdego inhibitora/aktywatora) , UWAGA: do prób odczynnikowych
nie dodajemy śliny)
2. Do każdej probówki dodać 0,5 ml koloidalnego roztworu skrobi (po uprzednim jej
wymieszaniu) i dokładnie wymieszać. Probówki inkubować przez 20 minut w temperaturze 37°C.
3. Po inkubacji pobrać 0,2 ml mieszaniny reakcyjnej, wprowadzić do czystych probówek, do których dodać także po 0,2 ml roztworu DNS. Zawartość wymieszać i umieścić na 5 minut w termobloku 100 °C
4. Po 5 minutach probówki wyjąć, schłodzić do temperatury pokojowej pod bieżącą wodą, dodać 0,6 ml wody destylowanej i dokładnie wymieszać.
5. Zmierzyć absorbancję roztworu przy λ=540 nm (A540) wobec próby odczynnikowej.
6. Z krzywej wzorcowej odczytać stężenie cukrów redukujących (mgmaltozy/ml). Wyniki
umieścić w tabeli.
7. Na podstawie tabeli określić wpływ badanych związków na aktywność enzymu.