Przyczyny róŜnorodności świata organicznego
Tym razem spróbujemy zrozumieć, dlaczego tak róŜnorodne jest Ŝycie. To problem,
który nurtował ludzi od dawna. Jaka jest tego przyczyna? Czy jest jedna? Dziś wiemy, Ŝe
róŜnorodność przyczyn jest najwaŜniejszym powodem tak wielu form, gatunków, kształtów,
kolorów... Te zajęcia poświęcimy omówieniu tych przyczyn i ich skutków.
Dwa podstawowe pytania to:
1. Skąd się wzięło Ŝycie?
2. Skąd wzięła się taka róŜnorodność gatunków?
Na oba pytania próbowano odpowiedzieć przez setki lat teorią kreacjonizmu. W
największym skrócie mówi ona, Ŝe całe Ŝycie (wszystkie gatunki) powstały na drodze
stworzenia (kreacji) przez Siłę WyŜszą (Boga). Wspierano się przy tym Biblią twierdząc, Ŝe
dokładny opis tej kreacji jest opisany w Księdze Rodzaju. Niektórzy na tej podstawie
obliczali wiek Ziemi na ok. 6-10 tysięcy lat. Do kreacjonizmu zalicza się tzw. teorię
inteligentnego projektu – pogląd zakładający u początku ewolucji akt sprawczy, podjęty przez
Boga. Kreacjonizm jednak rzadko przenika do dziedzin biologicznych i zazwyczaj jego
zwolennicy poruszają się w przestrzeni filozoficznej lub religioznawczej.
Dominacja kreacjonizmu była niczym nie zachwiana jeszcze w XVIII wieku. Karol
Linneusz, twórca systemu klasyfikacji organizmów (który stał się podstawą współczesnej
taksonomii) był równieŜ gorącym zwolennikiem tego nurtu, choć pod koniec Ŝycia zaczął
dopuszczać istnienie zmienności pokreacyjnej w postaci np. mieszańców. Nie była to spójna
teoria, nie zalicza się go więc do ewolucjonistów, ale to waŜna przesłanka do tezy, Ŝe juŜ
wówczas zaczęły pojawiać się próby wytłumaczenia odstępstw do teorii kreacjonistycznej,
które coraz częściej obserwowano. W XVIII wieku obalono teorię samorództwa w stosunku
do zwierząt. Wcześniej twierdzono np., Ŝe w mięsie larwy much powstają samoistnie.
Dopiero Redi udowodnił, Ŝe jeśli nie dopuści się do składania jaj przez muchy, to w mięsie
nie będzie ich larw. Nadal jednak uwaŜano, Ŝe ma to miejsce w przypadku mikroorganizmów.
Dopiero Ludwik Pasteur w 2 połowie XIX w. udowodnił, Ŝe wysterylizowana (przez wrzenie)
poŜywka będzie wolna od bakterii, jeśli będzie odpowiednio izolowana od powietrza.
Późniejszą konsekwencją tego odkrycia był rozwój higieny m.in. w chirurgii (mycie rąk i
narzędzi chirurgicznych przed operacją), co wybitnie podwyŜszyło skuteczność tej metody
medycznej.
Na początku XIX wieku Jan Chrzciciel Lamarck stworzył pierwszą teorię ewolucyjną.
ZałoŜył, Ŝe nie wszystkie gatunki powstały od razu, a jedne (prymitywne) zmieniały się z
biegiem czasu w inne (wyŜsze, bardziej zaawansowane). Dla niego czynnikiem sprawczym
takiej ewolucji były czynniki środowiska. To one powodowały, Ŝe organy uŜywane się
rozwijały, wzmacniały, a nieuŜywane uwsteczniały i zanikały. śyrafa np. ma tak długą szyję,
poniewaŜ musiała wysoko sięgać po poŜywienie. Dziś wiemy, Ŝe cechy nabyte przez
osobnika nie są dziedziczone, jak pamiętamy jednak początki genetyki to koniec XIX wieku,
Lamarck nie mógł więc skonfrontować swojej teorii z tą dziedziną. Jednak teza, Ŝe jedne
gatunki mogą przechodzić w inne stała się fundamentem darwinowskiej teorii ewolucji.
Lamarck co prawda uwaŜał, Ŝe moŜna to narysować w postaci wieŜy od najprostszych
(najbardziej prymitywnych) gatunków do człowieka – szczytu ewolucji (jak to sobie
udowodnimy – jest to bardziej skomplikowane, a człowiek wcale nie jest takim szczytem
moŜliwości ewolucji), ale juŜ sama interpretacja zmienności w czasie była rewolucyjna.
Z kolei Ŝyjący niemal równolegle z Lamarckiem Georges Cuvier był zdecydowanym
kreacjonistą. JednakŜe jego teoria była odmienna od poprzedników i zwana jest powszechnie
teorią katastrof. Cuvier twierdził, Ŝe w historii ziemi istniały takie zdarzenia, które
powodowały wymieranie gatunków, a ich miejsce Bóg zapełniał innymi. Jedną z katastrof był
biblijny potop, ale Cuvier twierdził, Ŝe było ich więcej. I – jak dowodzi dzisiejsza
paleontologia – w tym nie mylił się. Jak widać był to waŜny argument dla przyszłych prac
Darwina. Tu jednak musimy wspomnieć jeszcze o jednym wybitnym osiągnięciu Cuviera –
jego sztandarowym dziełem było stworzenie systematyki opartej nie tylko na morfologii
(budowie zewnętrznej), ale teŜ na porównaniu budowy anatomicznej (wewnętrznej – układów
i narządów). Na tej podstawie stworzył podwaliny takiego uporządkowania zwierząt, którego
wyraźne ślady moŜemy znaleźć w dzisiejszych jej odmianach (choć wciąŜ jest zmieniana).
Anatomia porównawcza jest wciąŜ jedną z najwaŜniejszych dziedzin wspierającą
systematykę. Dopiero w ostatnich latach znacząco wzrosło znaczenie genetyki molekularnej i
biochemii biopolimerów (głównie białek).
Niewątpliwie jedną z największych rewolucji nie tylko w biologii, ale i w rozwaŜaniach
światopoglądowych wywołało opublikowanie dzieła „O powstawaniu gatunków drogą
doboru naturalnego, czyli o utrzymywaniu się doskonalszych ras w walce o byt”. Darwin
w niej przedstawił podwaliny teorii, która dziś jest wiodącą i dobrze udokumentowaną
hipotezą powstania Ŝycia i bioróŜnorodności. Twierdził on, Ŝe gatunki powstają pod
wpływem działania doboru naturalnego, który powoduje, Ŝe osobniki słabiej przystosowane
do środowiska giną, a silniejsze (lepiej przystosowane) przeŜywają i wydają na świat
potomstwo. W ten sposób zachodzi ewolucja, która jest siłą udoskonalającą istniejące
gatunki. Ewolucja zachodzi w skali populacyjnej (mikroewolucja) i na kaŜdym wyŜszym
poziomie organizacji organizmów (makroewolucja). Jest pierwszą i podstawową przyczyną
zróŜnicowania istot Ŝywych. W przeciwieństwie do Lamarcka Darwin odrzucił wpływ
zmienności środowiskowej jako przyczyny zmienności ewolucyjnej, choć długotrwały wpływ
środowiska będzie oddziaływał na wykształcenie jakiejś cechy. Darwinowską teorię
ewolucjonizmu moŜna sprowadzić do następujących załoŜeń:
• świat istot Ŝywych nie jest niezmienny
• proces zmian jest ciągły i stopniowy
• wszystkie gatunki są ze sobą spokrewnione
• zmiany ewolucyjne są wynikiem doboru naturalnego.
Więcej o tym i Ŝyciu Darwina moŜna znaleźć na licznych stronach internetowych, m.in.:
http://www.ewolucja.org/
http://www.wiw.pl/biblioteka/klasycy_nauki/darwin_spisrzeczy.asp
Równolegle z Darwinem teorię ewolucji opisał Wallace i często mówi się o teorii
Wallaca-Darwina (szczególnie w Anglii). Trzeba jednak pamiętać, Ŝe Wallace wysunął teorię,
podczas gdy Darwin udokumentował swoje twierdzenie – było to więc o wiele dojrzalsze
patrzenie.
Syntetyczna teoria ewolucji
Wiek XX obfitował w rozwój takich dziedzin jak genetyka, biologia molekularna,
techniki mikroskopowe itp. które odsłaniały nowe „światy”, a w nich dowody na
prawdziwość teorii Darwina. Zaowocowały one syntetyczną, uwzględniającą odkrycia
podarwinowskie teorią ewolucji.
Procesy ewolucyjne wynikają zarówno z doboru naturalnego, jak i zmienności
genetycznej – zarówno mutacyjnej, jak i rekombinacyjnej (związanej z crossing-over). W
syntetycznej teorii ewolucji wymienia się róŜne zjawiska modyfikujące pulę genową
populacji wyjściowej. NaleŜą do niech:
- zjawiska losowe, np. gwałtowne zmiany warunków środowiskowych, dryf genetyczny
- mutacje prowadzące do powstawania nowych genotypów
- dobór naturalny, który faworyzuje lub eliminuje niektóre genotypy
-
rekombinacja, która przyczynia się do utrzymywania róŜnorodności genetycznej
(proszę przy okazji przypomnieć sobie, co to jest crossing-over)
- izolacja, która utrudnia krzyŜowanie się grup osobników (czyli mieszanie się genów)
– w tym przypadku pojawia się bariera (moŜe być geograficzna, ale moŜe wynikać z
róŜnej budowy narządów kopulacyjnych, co powoduje brak moŜliwości wystąpienia
kopulacji między osobnikami).
PowyŜej wymienione zjawiska nie zawsze występują w równym nasileniu, niektóre
mogą w ogólne nie funkcjonować, albo mogą działać następująco po sobie, np. w wyniku
mutacji pojawi się zmiana w budowie narządów kopulacyjnych, co doprowadzi do izolacji; w
miarę upływu czasu osobniki noszące w genotypie ową mutację i niezmutowane będą się
coraz bardziej od siebie róŜniły.
Skoro ewolucja jest teorią istnieje obowiązek udowodnienia jej istnienia. Dowody na
prawdziwość tej teorii moŜemy czerpać z wielu dziedzin. Bezpośredniego potwierdzenia
moŜemy szukać w paleontologii. Jednak ze względu na szczątkowość i przypadkowość
znalezisk (odnajdujemy bardzo nikłą część tego, co zostało po organizmach Ŝyjących dawno
temu) daje ona dość kontrowersyjne, często podwaŜane wyniki. Bezsprzecznie nie jest to
dziedzina, która samodzielnie wystarczyłaby do udowodnienia tej teorii. W ten teŜ sposób
kreacjoniści zazwyczaj podwaŜają jej słuszność. Dodatkowo wpływały na to liczne
fałszerstwa poszukiwaczy, którzy chcieli za wszelką cenę zaistnieć jako odkrywcy.
Przypadkiem najsłynniejszym jest zapewne tzw. „człowiek z Piltdown”1 .
Kopalnymi dowodami ewolucji są więc:
- Skamieniałości – szczątki lub całe organizmy zachowane w postaci .m.in. skamielin.
Przykładem moŜe być Archeopteryks (praptak) odkryty w Bawarii czy amonity i
trylobity, które są tzw. skamieniałościami przewodnimi – charakterystycznymi dla
danej epoki (pierwsze od Dewonu do kredy, drugie – w całej erze paleozoicznej).
- Odciski – to pozostałości między dwoma warstwami skał obrazujące kształty roślin
czy zwierząt.
- Odlewy – powstają gdy miejsce wypłukanej skamieniałości wypełniają krystalizujące
minerały lub osad.
- Formy przejściowe – to wymarłe organizmy o cechach przejściowych między dwoma
grupami systematycznymi. MoŜemy do nich zaliczyć np. następujące kręgowce:
Ichtiostega (ryby/płazy), Seymuria (płazy/gady), Archeopteryx (gady/ptaki).
- śywe skamieniałości – często pojedyncze gatunki bądź niewielkie ich grupy, które
Ŝyją do dziś, choć wszystkie spokrewnione gatunki dawno wymarły. Takimi
przykładami mogą być: stawonóg skrzypłocz (Limulus polyphemus), mięczak łodzik
(Nautilus sp.), okrytozaląŜkowy miłorząb dwuklapowy (Gingko biloba), ryba latimeria
(Latimeria chalumnae) czy gad hatteria (Sphenodon punctatus). Waszym zadaniem
jest sprawdzenie, w jakim okresie geologicznym grupy reprezentowane przez ww.
gatunki występowały licznie na ziemi.
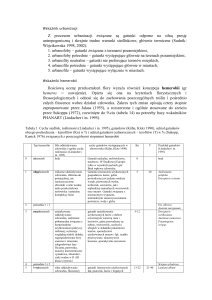
RozwaŜania
paleontologiczne zakończymy schematycznym przeglądem historii
geologicznej (tabela). Wymaga ona analizy – proszę przyjrzeć się jej dokładnie. Warto teŜ
Zwrócić uwagę na zakresy czasowe poszczególnych etapów. Ziemia istnieje ok. 4,6 miliarda
lat. śycie na ziemi (w postaci pierwszych udokumentowanych komórek prokariotycznych)
1
Okazało się, Ŝe "człowiek z Piltdown" (odkryty w 1912 r., fałszerstwo zdemaskowane ostatecznie w 1953 r.)
rzeczywiście był pół-człowiekiem, pół-małpą: fragment czaszki pochodził od człowieka (mieszkańca Ziemi
Ognistej) z okresu średniowiecza, dolna szczęka, mająca około 500 lat, naleŜała do orangutana z Borneo, a ząb
to kopalny ząb szympansa. Znalezisko wyglądało na starsze niŜ było w rzeczywistości dzięki zamoczeniu go w
roztworze Ŝelaza i kwasu chromowego. Ponadto kły zostały spiłowane, co wykazały badania pod mikroskopem,
tak Ŝeby pasowały do pozostałych fragmentów kości.
powstało ok. 3,5 miliarda lat temu. Rysunek z zegarem obrazuje proporcje czasowe
poszczególnych er. Dopełnieniem tej analizy będzie omówienie przyczyn róŜnorodności
geograficznej jako efektu wędrówki kontynentów, do czego wrócimy przy rozwaŜaniach
biogeograficznych pod koniec omawiania tego tematu.
Na szczęście dla teorii ewolucji dysponujemy równieŜ całym zestawem dowodów
pośrednich. PoniŜej omówimy sobie główne z nich.
Dowody z anatomii porównawczej.
Twórcą tej dziedziny jest Cuvier (juŜ to wspomnieliśmy). Okazało się, Ŝe analiza
budowy róŜnych organizmów (istniejących i – w miarę moŜliwości – równieŜ wymarłych)
daje duŜe moŜliwości prześledzenia zmian róŜnych ich fragmentów ciał, co prowadzi do
odpowiedzi na pytanie „co z czego powstało”.
MoŜemy więc obserwować zmiany w budowie i funkcjonowaniu róŜnych narządów i
ich układów. Często taką analizę robi się w odniesieniu do kręgowców, np. w odniesieniu do
budowy serca i układu krwionośnego. Ewolucja przebiega tu od układu jednoobiegowego i
serca zbudowanego z jednego przedsionka i jednej komory (bezŜuchwowce, ryby) poprzez
dwuobiegowy
z
sercem
dwuprzedsionkowym,
ale
jednokomorowym
(płazy),
dwukomorowym z niepełną przegrodą (większość gadów) do serca dwukomorowego
(krokodyle, ptaki i ssaki). Analogicznie moŜna rozwaŜać rozwój szkieletu kończyn (od płetw,
poprzez czworonogi o tułowiu zawieszonym na kończynach do takich, które mają ciało
podparte kończynami), narządów wymiany gazowej, budowy mózgu itd.
WaŜną przesłanką ewolucyjną są narządy homologiczne. Mają one odmienną budowę ,
ale wspólne pochodzenie. Odkrycie tego faktu jest bardzo waŜne, bo czasem takie homologie
trudno potwierdzić ze względu na bardzo róŜną budowę dziś Ŝyjących organizmów.
Przykładami narządów homologicznych są łuski gada i pióra ptaka; skrzydło nietoperza,
przednia łapa kreta i płetwa płetwala błękitnego; wąsy czepne fasoli i ciernie opuncji; korzeń
selera i korzeń epifitów; bulwy ziemniaka, bulwy kalarepy i łodyga czepna chmielu.
Narządy analogiczne z kolei mają podobną budowę, często spełniają podobną funkcję,
ale mają róŜne pochodzenie. Najczęściej w taki sposób porównujemy organizmy
niespokrewnione ze sobą. Przykładami mogą tu być ryzoid mszaka i korzeń marchwi,
pancerze pancernika i Ŝółwia, oko głowonoga i kręgowca, skrzydło nietoperza i skrzydło
waŜki.
Narządy szczątkowe to takie, które u przodków badanego organizmu pełniły waŜną rolę,
obecnie zaś są silnie zredukowane (w szczątkowej formie). Tu moŜemy wymienić
szczątkowo wykształcone miednice i tylne kończyny u pytona czy płetwala, szczątki skrzydeł
kazuara czy kiwi, uchyłek jelita ślepego u człowieka, trzecia powieka w kącie oka u
człowieka.
Atawizmy z kolei to cechy, które mogą się pojawić jako kuriozum u niektórych
osobników. Są one stosunkowo rzadkie i nawiązują do przodków danego ratunku.
Ograniczając się do człowieka moŜemy wymienić: większą liczbę kręgów ogonowych, kły
wystające poza linię zgryzu czy nadmierne owłosienie całego ciała (z twarzą włącznie).
Dowody z embriologii
Ernest Haeckel w połowie XIX wieku stwierdził, Ŝe „ontogeneza (rozwój osobnika) jest
powtórzeniem filogenezy (historii gatunku)”. Oznacza to, Ŝe podczas procesów rozwoju
(zarówno embrionalnego, jak i larwalnego) pojawiają się formy, które przypominają
przodków danego gatunku. Jest to zjawisko dość szerokie – często uŜywano go zestawiając
embriony róŜnych kręgowców (ryby, płaza, gada, ptaka i ssaka) pokazując, Ŝe m stadium
wcześniejsze, tym są one do siebie bardziej podobne. Ale dotyczy to jeszcze wcześniejszych
etapów – na etapie gastruli np. pozostały jamochłony, tzw. zwierzęta dwuwarstwowe. Z wielu
badań wynika, Ŝe nasi przodkowie teŜ musieli wyglądać podobnie.
Ten sam Ernest Haeckel stworzył tzw. teorię listków zarodkowych. Mówi ona, Ŝe kaŜdy
narząd czy układ narządów pochodzi z dokładnie określonej, jednej z 3 warstw komórek:
ektodermy (zewnętrznej), entodermy (wewnętrznej) i mezodermy (środkowej). Przykładowo
skóra i układ nerwowy pochodzi z ektodermy, przewód pokarmowy z entodermy, a narządy
rozrodcze z mezodermy. Inny wybitny embriolog przełomu XIX i XX wieku, August
Weissmann odkrył tzw., szlak komórek płciowych (proszę sobie to przypomnieć z
poprzedniego semestru – z lekcji o rozmnaŜaniu).
NajwaŜniejsze etapy embriogenezy rozmnaŜania płciowego to:
1. Zapłodnienie – połączenie się haploidalnych komórek rozrodczych i powstanie
diploialnej komórki zwanej zygotą.
2. Pierwsze podziały komórkowe zygoty zwane bruzdkowaniem prowadzi najpierw do
moruli (bryłki komórek), a następnie do pęcherzyka zwanego blastulą (jedna warstwa
komórek).
3. Blastula z jednej strony wpukla się i tworzy się dwuwarstwowy twór zwany gastrulą;
posiada otwór gębowy, nie posiada odbytu. Proces ten nazywa się gastrulacją. Na tym
etapie rozwojowym do dziś Ŝyją jamochłony. Jama wewnętrzna zwana jelitem jest
pierwotną jamą ciała zwaną teŜ jamą gastralną.
4. Jelito gastruli wpukla się i wyodrębniają się pęcherzyki zlokalizowane pomiędzy
ektodermą i entodermą. Komórki je otaczające są mezodermą (trzeci listek
zarodkowy) a otaczają one wtórną jamę ciała zwaną celomą.
5. Następnym etapem jest organogeneza czyli stopniowe tworzenie i dojrzewanie
poszczególnych narządów i grupowanie ich w układy.
PowyŜszy plan jest charakterystyczny dla większości zwierząt (z wyjątkiem
najprostszych form – u nich jest tylko nieco odmiennie).Jeśli przyjąć, Ŝe ewolucja ma
charakter progresywny, to w rozwoju zarodkowym mamy szansę odnajdywać formy
przodków badanych organizmów. MoŜna to prześledzić np. u zwierząt osiadłych, których
larwy są zazwyczaj wolno Ŝyjące co dowodzi, Ŝe organizmy te utraciły zdolność poruszania
się wtórnie, a ich przodkowie charakteryzowali się zdolnością do ruchu. To z kolei wyjaśnia
pozorną sprzeczność – z jednej strony mówimy, Ŝe jedną z cech zwierzęcych jest zdolność do
poruszania się, a z drugiej znamy wiele form osiadłych.
Dowody z fizjologii, biochemii i genetyki
Wiele zjawisk fizjologicznych czy biochemicznych przebiega podobnie – zdarza się to
nawet u organizmów daleko spokrewnionych ze sobą. Przykładami mogą być:
• bodźce elektryczne u wszystkich zwierząt przewodzone są w taki sam sposób
• procesy metaboliczne funkcjonują na zasadzie przeciwstawienia reakcji utleniania i
redukcji
• niemal wszystkie organizmy mają taki sam skład kodu genetycznego
• cytochrom c (por. lekcja o oddychaniu) ma podobną budowę u większości
organizmów
• replikacja DNA występuje u wszystkich organizmów
• tylko 6 białek histonowych występuje w przyrodzie
• podziały komórkowe zachodzą wg kilku podstawowych planów: mitotyczny,
mejotyczny i amitoza
Dowody z biogeografii
Rozmieszczenie roślin i zwierząt w róŜnych regionach na świecie nie jest jednorodne.
Wpływa na to szereg czynników: klimat i historia kontynentu (w tym czas izolacji od innych
obszarów).
Musimy zdawać sobie sprawę z tego, Ŝe lądy wędrują. Znajdują się one na tzw. płytach
kontynentalnych, które się przesuwają. Jeśli jedna płyta przesuwa się w przeciwnym kierunku
do drugiej, to na ich styku będziemy obserwowali duŜą aktywność sejsmiczną (trzęsienia
ziemi) i wulkaniczną. W dłuŜszym czasie tam „wyrosną” góry. Na schemacie obrazującym
historię Ŝycia na ziemi są zaznaczone główne etapy wędrówki kontynentów.
Jak widać poniŜsza historia ukazuje sytuację od końca ery paleozoicznej – wcześniej
istniało więcej niŜ 1 ląd, ale dla naszych rozwaŜań nie jest to waŜne. Z powyŜszego widać, Ŝe
w Permie nie istniały Ŝadne większe bariery wodne, powstrzymującą swobodną migrację
roślin i zwierząt. MoŜna by wręcz przyjąć, Ŝe wówczas głównym czynnikiem determinującym
rozmieszczenie były warunki klimatyczne.
Pangea czyli prakontynent rozerwała się na Laurazję (część północna) i Gondwanę
(część południową), które rozdzielone były morzem Tetydy. Stopniowo, przez dalsze
rozczłnokowanie powstawały kolejne kontynenty – z Laurazji powstała Ameryka Północna i
Eurazja, a z Gondwany Ameryka Południowa, Afryka (z Madagaskarem), Australia,
Antarktyda i... Dekan czyli Półwysep Indyjski. Ten ostatni uderzając w Azję spowodował
wypiętrzenie pasma Himalajów!
Jakie są tego konsekwencje? Proszę prześledzić jeszcze raz tabelę historii Ŝycia na
Ziemi w kontekście tego kiedy ostatecznie kontynenty traciły ze sobą kontakt. Najlepszym
przykładem takiej długotrwałej izolacji lądu i ewolucji biologicznej przebiegającej odrębnie
od innych terenów jest Australia – niemal wyłącznymi przedstawicielami ssaków na tych
terenach są torbacze i stekowce, a brak ssaków łoŜyskowych, które poza tym kontynentem są
wiodąc i niemal jedyne. Więcej – obserwujemy tam zajmowanie nisz ekologicznych przez
podobne gatunki – np. drapieŜnikiem jest wilk workowaty – torbacz, który nie tylko
zachowuje się podobnie do naszego wilka, ale i pokrój jego ciała przypomina naszego
drapieŜnika. PowyŜsze fakty powinny ułatwić Wam przyswojenie podziału na państwa
roślinne i krainy zoogeograficzne. Wcześniej jednak musimy wytłumaczyć sobie 2 terminy:
Endemit to organizm mający niewielki zasięg geograficzny występowania. Czasem jest
to jedno miejsce (np. archipelag wysp Galapagos to jedyne miejsce występowania Ŝółwi
olbrzymich).
Relikt (zwany teŜ Ŝywą skamieliną) to gatunek występujący na jakimś niewielkim
obszarze, który w przeszłości występował powszechnie. Przykładem moŜe być dębik
ośmiopłatkowy – relikt polodowcowy, który kiedyś występował na terenie całej dzisiejszej
Polski, a takŜe świstak.
Państwa roślinne
Holarktyczne (Holarctis) - zajmuje największy obszar (cała Europa, Azja bez
półwyspów Indyjskiego i Malajskiego, północna Afryka i prawie cała Ameryka Północna)
róŜnorodne formacje roślinne: m.in. tundra, tajga, lasy liściaste, stepy, półpustynie i pustynie;
flora: kilkadziesiąt endemicznych rodzin, łącznie 50 tysięcy gatunków, stąd wywodzą się
m.in. wierzby, klony, cały wielki rząd róŜowców, liczne trawy i motylkowe;
charakterystyczne nagonosienne: sosny, modrzewie i miłorzęby.
Paleotropikalne (Palaeotropis) - centralna i południowa Afryka oraz południowowschodnia Azja łącznie z międzyzwrotnikowymi wyspami pacyficznymi; formacje roślinne:
przewaga puszcz tropikalnych, lasów monsunowych, a na obszarach suchych sawann i pustyń
gorących; flora: 40 rodzin endemicznych, bogactwo gatunkowe 65 tysięcy gatunków samych
roślin nasiennych; charakterystyczne rośliny okrytonasienne: figowce, bananowce, palmy,
nagonosienne - sagowce i gniotowce; stąd wywodzą się rośliny uprawne: m.in. kawa, herbata,
bawełna.
Neotropikalne (Neotropis) - Ameryka Środkowa i Południowa (bez skrawka
najbardziej wysuniętego na południe); typowe formacje roślinne: lasy równikowe (selwa
amazońska) i stepy (pampa); flora: ok. 40 rodzin endemicznych; bardzo duŜa róŜnorodność
gatunkowa (90 tys. gatunków nasiennych, nie licząc paprotników i mszaków); charakterystyczne rośliny: liczne kaktusy i storczyki, palmy i sagowce; stąd wywodzi się wiele roślin
uŜytkowych, np. papryka, kukurydza.
Australijskie (Australis) - Australia łącznie z Tasmanią; charakterystyczne formacje
roślinne: lasy twardolistne, sawanny, półpustynie i pustynie, rośnie tu ok. 13 tysięcy
gatunków roślin okrytonasiennych, z czego 75% to endemity skupione w 12 rodzinach;
charakterystyczne są eukaliptusy i akacje.
Przylądkowe (Capensis) - najmniejsze państwo roślinne (zajmuje niecałe 100 tys. km2 );
leŜy na najbardziej na południe wysuniętym skrawku Afryki; formacja roślinna: zbliŜona do
śródziemnomorskiej - gł. wiecznie zielone zarośla twardolistne; spośród rosnących tu 6
tysięcy gatunków 80% stanowią endemity; występują m.in. przypołudniki (tzw. Ŝywe
kamienie), pelargonie, zielistki itp.
Holantarktyczne (Holantarctis) - południowe krańce Ameryki Południowej i Nowej
Zelandii, wybrzeŜa Antarktydy i wiele pomniejszych wysp wokół tego kontynentu; formacje
roślinne: w większości tundra; flora uboga - ok. 10 tysięcy gatunków roślin nasiennych,
większość to endemity; charakterystyczny jest buk południowy (Nothofagus), spokrewniony z
naszym rodzimym bukiem, i trawa śmiałek antarktyczny.
Wszystkie rośliny wodne Ŝyjące w morzach i oceanach zalicza się niekiedy do
odrębnego państwa morskiego.
Krainy zoogeograficzne
Palearktyczna (Palearktyka) - obszarowo odpowiada euroazjatyckiej części państwa
holarktycznego; występuje tu wiele gatunków Ŝyjących równieŜ w krainie nearktycznej (m.in.
niedźwiedź polarny, renifer, sowa śnieŜna) bądź blisko ze sobą spokrewnionych gatunków
naleŜących do jednego rodzaju (ryś europejski i kanadyjski, Ŝubr i bizon, niedźwiedź brunatny i grizzly, jeleń europejski i wapiti, róŜne gatunki lemingów itp.); bardziej róŜni się fauna
południowych rejonów tych krain - w Palearktyce brak nearktycznych przedstawicieli
szopowatych (np. szop pracz), a w Nearktyce świniowatych (dziki) i łaszowatych (wiwery,
cywety).
Etiopska - odpowiada afrykańskiej części roślinnego państwa paleotropikalnego; kraina
szczególnie bogata w ssaki i ptaki; kilka rodzin endemicznych: m.in. strusie, sekretarze,
mrówniki, góralki, Ŝyrafy; duŜa róŜnorodność ssaków kopytnych (antylopy i zebry), obecność
małp człekokształtnych (szympansy, goryle) i zwierzokształtnych (pawiany, koczkodany).
Orientalna - południowo-wschodnia Azja łącznie z wyspami Borneo, Jawą, Sumatrą i
Filipinami; świat zwierzęcy tej krainy podobny do świata zwierzęcego krainy etiopskiej - w
obu występują m.in. słonie, Iwy, lamparty, bawoły, hieny, małpy (zwierzokształtne makaki),
kobry i krokodyle; podobnie jak w krainie palearktycznej występują krety, niedźwiedzie,
jelenie i wkraczające na północ tygrysy, hieny i gepardy; orientalne endemity, m.in. gibony i
orangutany; we wschodniej części Archipelagu Malajskiego fauna pośrednia między faunami
krain orientalnej i australijskiej (ten rejon nosi nazwę Wallacea).
Neotropikalna - to Ameryka Południowa i Środkowa - w Ameryce Południowej
rozwinęły się m.in. specyficzne rzędy torbaczy (np. owadoŜerne oposy), cały rząd
szczerbaków (leniwce, mrówkojady), nielotne ptaki nandu, małpy szerokonose (wyjce,
czepiaki) oraz specyficzne grupy gryzoni, w tym największy gryzoń świata - kapibara;
połączenie z Ameryką Północną w pliocenie spowodowało, Ŝe zaczęły tu napływać z północy
m.in. koty, szopy i lamy; jednocześnie część zwierząt południowoamerykańskich rozpoczęła
wędrówkę na północ (m.in. pancerniki i oposy).
Australijska - od dawna izolowana od innych kontynentów, dlatego fauna składa się
właściwie z samych endemitów; typowi mieszkańcy: róŜne gatunki torbaczy - drapieŜne
(diabeł tasmański, niegdyś takŜe wilk workowaty), roślinoŜerne (kangury, koale), workowate
krety; występują tu endemiczne stekowce - dziobaki i kolczatki, oraz 12 rodzin endemicznych
ptaków (w tym ptaki rajskie oraz nielotne kazuary, emu i kiwi); rozprzestrzeniły się tu gatunki
sprowadzone przez człowieka - psy dingo, koty, lisy i inne.
Mam nadzieję, Ŝe udało nam się podczas tego wykładu jasno omówić przyczyny
zmienności i róŜnorodności organizmów Ŝywych. Pozostają do omówienia mechanizmy
ewolucji, specjacja, prawidłowości ewolucji, biogeneza i antropogenezę przełoŜymy na
następne zajęcia. Wówczas teŜ podam źródła, na których warto się oprzeć.
Zadanie domowe:
stawonóg skrzypłocz (Limulus polyphemus)
mięczak łodzik (Nautilus sp.)
okrytozaląŜkowy miłorząb dwuklapowy (Gingko biloba)
ryba latimeria (Latimeria chalumnae)
gad hatteria (Sphenodon punctatus).
W jakim okresie geologicznym grupy reprezentowane przez ww. gatunki występowały
licznie na ziemi?