Kora ruchowa
Kora ruchowa stanowi zasadniczy element kontroli ruchowej średniego poziomu.
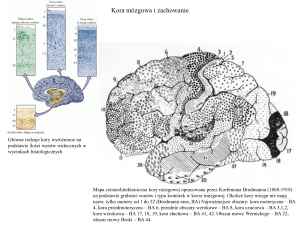
Lokalizacja kory u różnych ssaków, w tym u małp (1870), potwierdziła lokalizacje kory i
sekwencję obszarów (nogi, ręce, twarz) przewidzianą przez Jacksona.
Motoryczny homunkulus otrzymany przez Penfielda i Rasmussena (1950) metodą stymulacji kory mózgowej.
Trzy obszary kory ruchowej
Kora ruchowa znajduje się w płacie czołowym (frontal lobe), tuż przed bruzdą centralną (Central Sulcus) nazywaną też
bruzdą Rolanda (Rolandic fissure) i jest podzielona na trzy wzajemnie połączone obszary, pierwszorzędową korę ruchową
(MI), dodatkową korę ruchową (SMA) i korę przedruchową (PM). SMA i PM stanowią wspólnie drugorzędową korę ruchową
(MII). Funkcjonalnie oznacza to wielokrotną reprezentację mapy ruchowej w korze mózgowej.
Kora ruchowa
Dokładniejsze studia (intracortical microstimulation or ICMS) pokazały, ze neurony w pierwszorzedowej korze ruchowej (MI)
aktywują nie pojedyncze mięśnie, a te same mięśnie są aktywowane przez wiele różnych obszarów kory. Na wykresach widać
miejsca w MI, stymulacja których wywołuje aktywność EMG (wskazując na połączenie monosynaptyczne) w mięśniu ramienia
(deltoid muscle) i nadgarstka (extensor carpi radialis; ECR). Mapy topograficzne (1/próg stymulacji) ukazują zarówno
redundancje jak i przekrywanie się reprezentacji. Umożliwia to aktywacje różnych grup mięśni w różnych zadaniach.
Trzy obszary kory ruchowej - wejścia
Wejścia do kora ruchowej u małpy. A. Wejścia do kory pierwszorzedowej: PMd (dorsal premotor area); PMv (ventral
premotor area), S1 (primary sensory cortex), SMA (supplementary motor area.), CMA (cingulate motor area). B. Wejścia do
kory przedruchowej: pola 5 i 7 Brodmana, kora przedczołowa (prefrontal cortex, pole Brodmana 46). Obszary 5 i 7 Brodmana
zajmują się integracją multisensoryczną w celu planowania ruchu. Obszar 46 zajmuje się pamięcią operacyjną (working
memory) i przetrzymuje w pamięci informacje o lokalizacji przestrzennej obiektów, tylko na czas wykonania ruchu.
Trzy obszary kory ruchowej - wejścia
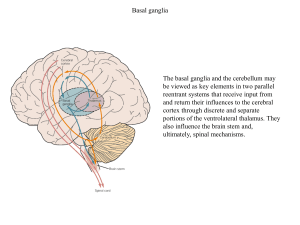
Kora przedruchowa i pierwszorzedowa kora ruchowa dostają również wejścia ze zwojów podstawy i móżdżku poprzez
wzgórze.
Plastyczność somatotopicznej organizaji kory ruchowej
A. Widok normalny somatotopicznej organizacji kory ruchowej szczura z obszarami reprezentującymi mięśnie łap przednich,
wąsów i okołooczne. B. Po przecięciu nerwu twarzowego, obszary mięśni łap przednich i okołooczne są zwiększone kosztem
obszaru wąsów. Reorganizacja może się odbyć nawet w czasie paru godzin.
Plastyczność somatotopicznej organizaji kory ruchowej cd
Ruchy zamierzone ulegają poprawie wraz z praktyką. B. Skany fMRI ukazujące aktywacje kory MI podczas wykonywania
sekwencji ruchowej ćwiczonej przez 3 tygodnie (prawo) oraz nowej sekwencji ruchowej (lewo). Reprezentacja wyćwiczonej
sekwencji jest większa niż sekwencji kontrolnej
Wielosynaptyczne i monosynaptyczne połączenia drogi
piramidowej z motoneuronami ruchowymi
Porównanie zakończeń drogi piramidowj u ssaka niższego (dydelf – rząd torbaczy), wyższego (kot) i u
naczelnych. Tylko u naczelnych neurony kory ruchowej tworzą monosynaptyczne połączenia z motoneuronami
a w rdzeniu kręgowym.
Komórki V warstwy kory motorycznej dają początek drodze
piramidowej
Z lewej: siedem warstw kory motorycznej wg.
klasyfikacji Cajala. Warstwy są wyróżnione na podstawie
względnej ilości dużych ciał neuronów (komórki
piramidalne) i małych ciał neuronów (komórki
piramidalne, gwiaździste i ziarniste). Z prawej: komórka
Betza kory motorycznej obrazowana metodą Golgiego.
Porównanie sześciu warstw kory w różnych obszarach
mózgu. W V warstwie kory motorycznej widoczne ciała
neuronów komórek Betza. Komórki Betza znajdują się
głównie w obszarze ruchowym nogi.
Równoległe drogi ruchowe
Pośrednie i bezpośrednie
połączenia kory ruchowej i pnia
mózgu z boczną i przyśrodkową
częścią rdzenia kręgowego.
Bezpośrenia droga korowordzeniowa (droga piramidowa,
corticospinal tract) omija
wybrzuszenie piramidowe w pniu
mózgu i schodzi do pnia mózgu,
gdzie tworzy połączenia
polisynaptyczne (u wyzszych
ssaków) lub monosynaptyczne (u
naczelnych) z motoneuronami. 85
% włókien przekracza linię
środkową i tworzy boczną drogę
korowo-rdzeniową, pozostałe
aksony tworzą grzbietową drogę
korowo-rdzeniową.
Drogi pośrednie: droga
czerwienno-rdzeniowa i droga
korowo-siatkowo-rdzeniowa.
Równoległe drogi ruchowe
Przecięcie drogi piramidowej
powoduje jednostronną słabość w
mięśniach u małpy. Ustępuje ona
po upływie paru miesięcy i
zwierzęta odzyskują normalne
funkcje ruchowe dzięki przejęciu
funkcji przez drogi pośrednie.
Jednakże indywidualne ruchy
palców są utracone na zawsze.
Kodowanie siły mięśni
Częstość odpalania neuronów w korze ruchowej koduje siłę mięśni. A1,2 - dwa rodzaje zachowania
neuronów w korze ruchowej podczas momentu siły przyłożonego do nadgarstka. B. W obu rodzajach
neuronów częstość odpalania wzrasta (liniowo) z siłą mięśni. (torque – ang. moment obrotowy)
Kodowanie populacyjne kierunku ruchu
Kierunek ruchu jest kodowany w korze motorycznej poprzez populacje
neuronów. (A) Wytrenowana małpa porusza ręką w ośmiu różnych
kierunkach. (B) Aktywność neuronu ruchowego w korze podczas ruchów
w ośmiu kierunkach. C.Krzywa strojenia neuronu. D. Wiele neuronów
korowych jest aktywnych w trakcie ruchu w danym kierunku. Kierunek
każdej linii reprezentuje preferowany kierunek danej komórki, a długość
linii jest proporcjonalna do częstotliwości jej wyładowań. Wypadkowy
‘wektor’ aktywności (czerwona strzałka) koduje wykonywany kierunek
ruchu (czarna strzałka) (Georgopoulos, 1982)
Potencjał przygotowujący
W ruchach generowanych wewnętrznie, na ok. 1 sekundy przed ruchem obserwowany jest ujemny potencjał
w EEG w obszarze dodatkowej kory ruchowej (SMA) zanim nastąpi aktywacja pierwszorzędowej kory
ruchowej. Jest to tzw. potencjał przygotowujący (readiness potential lub Bereitschaft potential), który
sygnalizuje planowanie ruchu. W eksperymentach pokazano, że BP pojawia się ok. 0.35 zanim badany
relacjonował świadom myślenie o wykonaniu ruchu.
Obszary przedruchowe
Aktywność mózgu w trakcie prostego, złożonego i wyobrażonego ruchu. A. Podczas naciskania sprężyny palcem wzrasta aktywność w
obszarze ręki w pierwszorzędowej korze motorycznej i sensorycznej. B. Podczas złożonego ruchu ręką aktywność obejmuje również
przyśrodkową korę przedruchową obejmującą dodatkową korę ruchową i (SMA) przeddodatkową korę ruchową (preSMA). C. Podczas
wyobrażania ruchu wykonywanego w pkt. B, wzrost aktywności występuje jedynie w przyśrodkowej korze przedruchowej. Wniosek:
dodatkowa kora ruchowa zawiera reprezentacje wyuczonych sekwencji ruchów.
Obszary przedruchowe
Visual cue: małpa miała dotykać podświetlonych paneli, Prior training: wykonanie wcześniej wyuczonej sekwencji. Neurony
w korze ruchowej MI odpalały przed i w trakcie ruchu, tak samo w obu zadaniach. Dodatkowa kora ruchowa (SMA) była
aktywna tylko przed i w trakcie zadania wyuczonego. Kora przedruchowa była aktywna tylko przed i w trakcie
wykonywanego zadania na podstawie informacji wzrokowej. Wniosek: dodatkowa kora ruchowa programuje wyuczone
sekwencje ruchowe. Kora przedruchowa bierze udział w przygotowaniu i wykonaniu ruchu w odpowiedzi na bodziec
zewnętrzny. Po długotrwałym treningu, aktywność w dodatkowej korze ruchowej zanika – sekwencje wykonywane
automatycznie ‘przenoszą się’ do kory pierwszorzędowej.
Obszary przedruchowe
Neuron w korze przedruchowej
staje się aktywny gdy małpa
przygotowuje się do ruchu w
lewo. Instrukcja (podświetlenie
jednego z czterech paneli)
wskazuje panel do naciśnięcia
po bodźcu wyzwalającym
(zapalenie się diody).
Odpalanie neuronu rozpoczyna
się po pojawieniu się instrukcji
i trwa do wykonania zadania po
bodźcu wyzwalającym.
ruchowe. Wniosek: kora
przedruchowa bierze udział w
przygotowaniu i wykonaniu
ruchu w odpowiedzi na bodziec
zewnętrzny (w tym
doświadczeniu - wzrokowy).
Transformacja sensomotoryczna
Transformacja reprezentacji sensorycznej otoczenia na sygnały kontroli mięśni nazywa się transformacją sensomotoryczną.
Oddzielne, równoległe drogi zapewniają transformację wzrokowo-motoryczną wymaganą do sięgania i chwytania. PMd –
premotor dorsal, PMv – premotor ventral.
Komórki zwierciadlane
Komórki w grzbietowej korze przedruchowej wykazują aktywność gdy małpa wykonuje zadanie lub gdy obserwuje wykonanie
zadania. Komórki te są więc zaangażowane w abstrakcyjną reprezentację ruchu. A. Aktywność neuronu podczas obserwacji
chwytania wykonywanego przez inna małpę. B. Aktywność neuronu podczas obserwacji chwytania wykonywanego przez
eksperymentatora. C. Aktywność neuronu podczas chwytania wykonywanego przez małpę.
Obwody ruchowe - synteza
1.
Ruch zamierzony
rozpoczyna się ‘programem
centralnym’, który aktywuje
w odpowiedniej sekwencji
obszary kory ruchowej.
2.
Włókna korowo-rdzeniowe
pobudzają motoneurony.
3.
Motoneurony wywołują ruch
mięśni.
4.
Poprzez kolaterale, włókna
korowo-rdzeniowe
pobudzają komórki czuciowe
(informacja o wysłanych
komendach).
5 i 6 Wyjścia czuciowe z mięśni,
stawów i skóry dostarczają
informacji zwrotnej do kory
motorycznej o stanie skurczu
mięśni i fazie wykonania
ruchu.
Mowa i komunikacja
Gra świerszcza
Mechanizm generacji dźwięku u świerszcza.
Pocieranie skrzydłami o siebie powoduje przesuwanie
‘smyczka’ po ‘pile’, co wywołuje wibracje skrzydła.
Poprzez wysyłanie dźwięku paczkami (czirpami) świerszcz może sygnalizować rożne stany: nawoływanie,
agresję, partnerstwo.
Pieśń świerszcza – mechanizmy neuronalne i genetyczne
A. Zależność dźwięków świerszcza (czirpów) od aktywności komórek mięsni otwierających skrzydła (środek) i
zamykających skrzydła (dół) . Generator rytmu znajduje się w zwojach w odwłoku B. Elektryczna stymulacja
neuronów komenderujących (sygnał górny) wywołuje czirpowy wzorzec aktywności w motoneuronach. C. Wzorzec
pieśni u dwóch odmian dzikich świerszczy (zapis górny i dolny) oraz ich potomstwa (zapisy środkowe). Wniosek:
każdy genotyp jest związany z innym wzorcem dźwiękowym.
Pieśń świerszcza – mechanizmy kontroli neuronalnej
Hierarchiczna kontrola generacji pieśni u świerszcza. Stymulacja neuronów komenderujących inicjuje i utrzymuje generacje
czirpów. Włókna te znajdują się pod kontrolą wyższych układów. Ciało grzybiaste – najwyższy poziom hierarchii
motorycznej, przechowuje programy pieśni. Stymulacja ciała grzybiastego może wywołać różne pieśni (np. wołającą lub
rywalizacji) w zależności od siły, częstości i lokalizacji stymulacji. Cały układ znajduje się pod kontrolą hormonalną –
śpiewają głównie samce w okresie dojrzałym.
Aparat głosowy ptaka
Wydawanie głosu przez ptaka. A. mięśnie wzdłuż przełyku są napięte, u podstawy – luźne. Ściany membrany są sztywne
i przepływ powietrza odbywa się na dużym przekroju nie powodując dźwięku. B. mięśnie wzdłuż przełyku są luźne, u
podstawy – napięte. Ściany membrany są rozluźnione i wgłębiają się do wewnątrz. Przepływ powietrza odbywa się na
małym przekroju i powoduje oscylacje membrany o wysokiej częstości.
Wpływ genów i stymulacji sensorycznej
Wpływ genów i otoczenia na pieśń wróbla. A. Na wolności pieśni trzech odmian wróbla są różne. B. Dorastanie w izolacji ma
wpływ na pieśń lecz zachowuje jej strukturę. C. Utrata słuchu u młodych powoduje zanik struktury i różnic pomiędzy
odmianami. Wniosek: istnieje okres krytyczny w rozwoju ptaków podczas którego słuchanie pieśni jest niezbędna do jej
wykształcenia. Tworzy się wzorzec dźwiękowy w obwodach słuchowych (pamięć) służący do rozpoznania i generacji pieśni
danego gatunku w okresie dorosłym.
Rola hormonów
A. Pieśń partnerska jest śpiewana przez samce, a przez samice nie (kastracja samców powoduje zanik pieśni, męskie hormony
androgeny (A) – jej przywrócenie). Samice pod wpływem samych androgenów nie tworzą pieśni lecz w wyniku
wcześniejszego podania estradiolu (E) (metabolizującego testosteron), tworzą pieśni, tak jak samce. Szare koła symbolizują
wielkość obszaru nucleus hyperstriatum ventrale (HVc), który, podobnie jak obszar RA i X, jest większy u samców (B). C.
Histologiczne porównanie HVc u samców, u samic z testosteronem i samic z grupy kontrolnej.
Pieśni ssaków
A.
Mruczenie kota. Zapis EMG
mięsni gardła i przepony
oraz ciśnienia powietrza w
gardle ukazuje dokładne
zależności (synchronizację)
w czasie pomiędzy
aktywnością różnych
elementów emisji głosu.
B.
Sonogram wieloryba.
Pojedyncza pieśń (cztery
paski – ok 10 min) potrafi
trwać do 30min i jest
wiernie odtwarzana przez
danego osobnika. Jest to
być może najbardziej
rozwinięte pojedyncze
zachowanie u zwierząt.
Aparat wokalny u człowieka
Emisja głosu opiera się na
zasadzie tłoczonego powietrza.
Wymaga: źródła ciśnienia
(przepona, mięśnie oddechu i
brzucha), elementów
wibrujących (struny głosowe) i
rezonatorów i artykulatorów
(gardło, usta, język, policzki,
zatoki).
W grze na trąbce źródłem ciśnienia są
płuca, elementem wibrującym –
wargi, a rezonatorem – trąbka.
Aparat wokalny u człowieka – źródło ciśnienia
Aktywność mięśni oddechowych podczas wokalizacji. Dobrze widoczna precyzyjna
sekwencja aktywności różnych grup mięśni w trakcie wydawania głosu.
Aparat wokalny u człowieka – struny głosowe
A. Przekrój przez gardło ukazujący struny głosowe (vocal folds). A. Epiglottis (nagłośnia), vocal folds (struny głosowe),
trachea (tchawica). B. Ułożenie strun w różnych warunkach. Wysokość dźwięku zależy od długości, napięcia i odległości
pomiędzy strunami. Naturalna częstość krtani wynosi u dorosłego mężczyzny ok.. 125 Hz, u kobiety 210 Hz, u dzieci
powyżej 300 Hz.
Aparat wokalny u człowieka – tworzenie zgłosek
Ułożenie języka przy
generacji samogłosek
/i/, /a/, /u/. B.Pole
przekroju
poprzecznego obszaru
mowy od gardła (0) do
ust (16). C. Symulacja
komputerowa.
Odcinki gardłowe i
ustne mogą być
niezależnie regulowane
językiem, co jest
podstawą tworzenia
samogłosek i ludzkiej
mowy. Część gardłowa
obszaru mowy jest
zmniejszona lub
nieobecna u
szympansów,
Neandertala i u
noworodków
Wokalizacja u człowieka – obwody neuronalne
Kora motoryczna steruje
mięśniami mowy poprzez nerwy
czaszkowe i trakt rdzeniowo –
korowy. Wyższy poziom kontroli
obejmuje (prawdopodobnie) zwoje
podstawy i móżdżek. Najwyższy
poziom kontroli znajduje się w
polu Brocka. Linią przerywaną
zaznaczono drogi odpowiedzialne
za emocjonalną zawartość
wokalizacji.