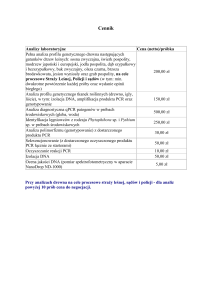
Acta Sci. Pol., Biotechnologia 15 (2) 2016, 15-30
ISSN 1644–065X (print) ISSN 2083–8654 (on-line)
TECHNIKI MUTAGENEZY LOSOWEJ PROWADZĄCEJ
DO ZWIĘKSZENIA POLIMORFIZMU 1
Jakub Kustosz
Zachodniopomorski Uniwersytet Technologiczny w Szczecinie
Streszczenie. Inżynieria białek jest obecnie bardzo ważnym działem we współczesnej biotechnologii. Polega ona między innymi na losowej mutagenezie genów kodujących białka
i pozwala na tworzenie enzymów oraz białek o pożądanych cechach lub o zwiększonej
aktywności. Istnieje wiele technik umożliwiających losową zmianę w sekwencji aminokwasowej białek poprzez zmianę sekwencji nukleotydów w kodujących je genach. Można
to osiągnąć dzięki poddaniu całych bakterii lub DNA działaniu związków chemicznych
(1) bądź wystawić je na promieniowanie UV (2), przy wykorzystaniu zmodyfikowanych
technik PCR lub RCA (Error-prone rolling circle amplification) – z zamierzonymi błędami,
w mutowanych szczepach (3) lub przy użyciu jednej z metod tasowania DNA (4). Dzięki
tym technikom i odpowiednim sposobie selekcji zmutowanych klonów możliwe jest przyspieszenie ewolucji genomów na potrzeby przemysłu. Poniższy artykuł przedstawia opis
tych metod i przykłady ich udanego wykorzystania do modyfikacji białek.
Słowa kluczowe: mutageneza chemiczna i fizyczna, EP-PCR, RCA, zmutowane szczepy,
tasowanie DNA, StEP, RACHITT
WSTĘP
W dzisiejszych czasach duży nacisk kładzie się na wydajność i zmniejszenie kosztów
przy równoczesnym zwiększeniu zysków płynących z produkcji. Dotyczy to również
enzymów i białek wykorzystywanych w przemyśle chemicznym, spożywczym czy farmaceutycznym. Jednak każdy enzym ma swoje ograniczenia związane ze specyficzną
zależnością substratową, optymalną temperaturę i pH. Często białka nie są w stanie spełnić wymagań, jakie przed nimi stawia przemysł oparty na procesach enzymatycznych.
Rozwiązaniem tego problemu jest ewolucja białek. Zachodzi ona w naturalny sposób
na poziomie genów. Jednak dla przemysłu enzymatycznego jest to proces zbyt wolny.
© Copyright by Uniwersytet Przyrodniczy we Wrocławiu
Adres do korespondencji – Corresponding author: Jakub Kustosz, Zakład Cytogenetyki Molekularnej, Wydział Biotechnologii i Hodowli Zwierząt, Zachodniopomorski Uniwersytet Technologiczny, ul. Doktora Judyma 6, 71-466 Szczecin, e-mail: [email protected]
16
J. Kustosz
Dlatego opracowano metody przyspieszające naturalne procesy. Zostały one scharakteryzowane w dalszej części artykułu.
MUTAGENEZA CHEMICZNA I FIZYCZNA
Jednymi z najstarszych metod mutagenezy genów kodujących białka są metody chemiczne i fizyczne. W tych przypadkach mutanty generowane są poprzez traktowanie ich czynnikami chemicznymi lub fizycznymi. Do mutagenezy chemicznej wykorzystuje się taki
związek jak metanosulfonian etylu (EMS). Powoduje on alkilacje reszt guanidyny, co
prowadzi do nieprawidłowego kopiowania podczas replikacji DNA (rys. 1) [Selifonova
i in. 2001].
Rys. 1. Alkilacja reszt guanidyny (modyfikacja: URL1)
Fig. 1. Alkylation guanidine (modification: URL1)
Mutagenezę z zastosowaniem EMS można prowadzić in vivo przy użyciu całych komórek lub in vitro z zastosowaniem DNA (rys. 2). Kwas azotowy jest kolejnym chemicznym mutagenem. Powoduje on powstanie mutacji punktowych przez deaminację. Może
powodować transwersje punktowe (A/T do G/C i vice-versa) [Tessman i in. 1964]. Inne
mutageny chemiczne to między innymi: mitomycyna C, N-metylo-N-nitrozomocznik
(MNH), diepoksybutan (DEB) oraz 1,2,7,8-diepoksyoctan (DEO), metanosulfonian metylenu (MMS) i metanosulfonian etylenu (EMS), N-metylo-N’-nitro-N-nitrozoguanidyna
(MNNG), 4-nitrochinolino-1-tlenku (4-NQO), 2-aminopuryna (2AP), hydroksyloaminy
(HA), wodorosiarczan (BS) oraz metoksyloaminy (MA) [Labrou 2010].
Posiew komórek
zmutowanych na
podłoże selekcyjne
Culture mutant cells
Dodanie do pożywki
czynnika mutagennego
Adding a mutagen
Hodowla komórkowa
Cell culture
Inkubacja komórek
czynnikiem mutagennym
Incubation of cells
with a mutagen
Selekcja mutantów
Mutant selection
Rys. 2. Schematyczne przedstawienie mutagenezy chemicznej (modyfikacja: URL2)
Fig. 2. Schematic representation of chemical mutagenesis (modification: URL2)
Acta Sci. Pol.
17
Techniki mutagenezy losowej...
Do mutagenów fizycznych możemy zaliczyć: promieniowanie jonizujące, w którego
skład wchodzą promienie X, promienie gamma, alfa, beta oraz protony i neutrony; promieniowanie elektromagnetyczne i ultrafioletowe. Promieniowanie jonizujące może powodować zmiany w sekwencji DNA i aberracje chromosomowe. Promienie X sprawiają,
że sekwencja DNA ulega zmianie na skutek pęknięć w łańcuchu DNA – pęknięcia jedno
lub dwuniciowe. Natomiast emisja promieniowania UV powoduje powstanie dimerów
pirymidynowych (rys. 3) [Tanooka i Munakata 1978, Brandenburger i in. 1981, Rosenbluth i in. 1985, Little i in. 1997].
Promieniowanie UV
UV radiation
powstanie dimerów
pirymidynowych
formation of pyrimidina
dimers
Rys. 3. Schemat powstawania dimerów pirymidynowych (modyfikacja: URL3)
Fig. 3. Schematic formation of pyrimidine dimers (modification: URL3)
Metody mutagenezy chemicznej i fizycznej były używane głównie w poprzednim
wieku. Jednym z przykładów wykorzystania takich technik była próba mutagenezy genu
psbDI z Synechocystis sp. PCC 6803. Do wywołania zmian wykorzystano wodorosiarczan sodu. Mutageneza była skierowana do konkretnego fragmentu DNA kodującego
określony region białka. Ostatecznie uzyskano 15 różnych zmian aminokwasowych.
Większość ze zmian nie wpływała na aktywność enzymu. Jednak 3 z nich spowodowały
zakłócenie aktywności, a wpływ dwóch był nieoczekiwany dla naukowców. Komórki posiadające podwójną mutację nie miały aktywnego enzymu, co sugeruje ważną rolę tych
aminokwasów w działaniu centrum aktywnego [Ermakova-Gerdes i in. 1996]. W taki
sposób można określić funkcję niektórych aminokwasów, których wpływ na aktywność
całego białka nie jest znany.
Obecnie jednak metody mutagenezy chemicznej i fizycznej nie mają większego zastosowania, ponieważ mimo niewielkich kosztów indukowania mutantów mają one kilka
wad. Gdy dany mutant zostanie wyselekcjonowany, to nie mamy pewności, że zmiana
zaszła w interesującym nas genie. Rozwój inżynierii genetycznej przyczynił się do opracowania nowych, bardziej wydajnych technik [Selifonova i in. 2001].
Biotechnologia 15 (2) 2016
18
J. Kustosz
Jedna z nowszych prac wykorzystująca te techniki miała na celu zwiększenie zdolności bakterii Bacillus sp. do biodegradacji barwników. Degradacja ich jest dużym problemem w przemyśle tekstylnym. Do przeprowadzenia doświadczenia wykorzystano
promieniowanie UV i bromek etydyny. Mutanty zostały poddane selekcji na podstawie
ich zdolności do odbarwienia czerwieni Konga. Zmutowane szczepy były w stanie odbarwiać barwnik w wyższym stężeniu i w krótszym czasie. Enzymem odpowiedzialnym
za degradację czerwieni Konga okazała się azoreduktaza. W zmutowanych szczepach
widoczny był jej wzrost o około 53% w rozdziale SDS-PAGE [Gopinath i in. 2009].
Kolejnym badaniem wykorzystującym związki chemiczne do mutagenezy była próba
modyfikacji Ashbya gossypii, który jest wykorzystywany w przemyśle do produkowania witaminy B2 (ryboflawiny). Grzyb ten powodował ekspresję endonukleazy I (EGI)
z Trichoderma reesei, a celem badania było poprawienie ogólnej zdolności grzyba do
pozakomórkowego wydzielania tego enzymu. Z tego powodu zastosowano mutagenezę
losową z zastosowaniem EMS. Technika chemicznej mutagenezy w tym przypadku przypominała tę z rysunku 1. Jeden z mutantów S436 posiadał 1,4–2-krotnie wyższą aktywność wydzielania zewnątrzkomórkowego wszystkich enzymów mierzonych w doświadczeniu (EGI, β-glukozydazy, α-amylazy, lipazy) niż w przypadku szczepu kontrolnego.
Jeden z mikroorganizmów miał większą aktywność w przypadku EGI i α-amylazy. Dwa
inne szczepy charakteryzowały się 2–3-krotną poprawą tylko do pojedynczych białek.
Jeden ze szczepów miał większą aktywność w przypadku β-glukozydazy, natomiast drugi
EGI [Ribeiro i in. 2013].
In vivo
zastosowanie mutagena
chemicznego np. EMS
chemical mutagen
pET
pET
inkubacja w 37°C
incubation 37°C
izolacja plazmidu
plasmid izolation
komórki DH5α
DH5α cells
In vitro
pET
pET
Ndel + Xhol
Ndel
Xhol
zmutowane komórki
DH5α
treated DH5α cells
pET
tranformacja
w komórkach BL-21
transformation cells BL-21
niezmutowany
wektor pET
untreated pET vector
pET
komórki BL-21
BL-21 cells
Rys. 4. Schematyczne przedstawienie zastosowania techniki DuARCheM
(modyfikacja: Mohan i Banerjee 2008)
Fig. 4. Schematic representation of the use of technology DuARCheM
(modification: Mohan i Banerjee 2008)
W 2008 roku została wprowadzona nowa metoda mutagenezy losowej wykorzystująca związki chemiczne. Technika ta została nazwana dwutorowym podejściem do lo-
Acta Sci. Pol.
Techniki mutagenezy losowej...
19
sowej mutagenezy chemicznej (ang. Dual Approach to Random Chemical Mutagenesis
(DuARCheM)). W technice tej wykonuje się zarówno mutagenezę chemiczną in vivo,
jak i manipulacje genetyczne in vitro. Pozwala ona na wprowadzenie zmiany wyłącznie
w interesującym nas genie. Komórki z plazmidem zawierającym sekwencję DNA, którą
chcemy zmienić, hoduje się ze związkiem mutagennym. W drugim etapie izoluje się
plazmid i trawi enzymami restrykcyjnymi w celu izolacji genu. Tak uzyskana sekwencja
nukleotydowa jest wklejana do wektora ekspresyjnego (rys. 4). Metoda ta naśladuje podejście na podstawie techniki PCR [Mohan i Banerjee 2008].
PCR Z ZAMIERZONYMI BŁĘDAMI (ANG. ERROR PRONE PCR – EP-PCR)
Jedną z podstawowych technik używanych w losowej mutagenezie jest EP-PCR, czyli
PCR z zamierzonymi błędami. W przeciwieństwie do tradycyjnej reakcji w metodzie tej
dochodzi do syntezy nowej nici DNA w sposób niekomplementarny przez polimerazę
charakteryzującą się brakiem zdolności naprawczych [Cadwell i Joyce 1992]. Można
zwiększyć niedokładność powielania DNA w EP-PCR także poprzez zastosowanie warunków reakcji zmniejszających dokładność polimerazy Taq DNA podczas syntezy. Najprostszym sposobem obniżenia dokładności polimerazy o niskiej wierności kopiowania
jest wprowadzenie do buforu reakcyjnego jonów Mn2+ wyższego stężenia jonów Mg2+
i dodanie różnego stężenia dNTP niż w przypadku tradycyjnej techniki PCR [Cadwell
i Joyce 1992, 1994, Cirino i in. 2003]. Obecność jonów Mn2+ i nadwyżka dGTP i dTTP
w reakcji amplifikacji prowadzi do błędu z częstością około 1 nukleotydu na tysiąc par
zasad w końcowej bibliotece mutantów [Caldwell i Joyce 1994, Cirino i in. 2003, Bessler
i in. 2003, Balci i in. 2014]. W celu uzyskania wyższego poziom zmian w konkretnym
odcinku DNA można zastosować analogi nukleotydów trójfosforanowych [Zaccolo i in.
1996].
Budowa idealnej biblioteki polega na uzyskaniu wszystkich możliwych mutacji występujących z równych częstością. Niestety, ten typ reakcji PCR nie jest jednak do końca losowy. Powstałe biblioteki klonów w zależności od zastosowanej polimerazy DNA,
natury kodu genetycznego i procesu amplifikacji są preferencyjne [Neylon 2004]. Wykorzystanie do reakcji EP-PCR polimerazy Taq może spowodować, że niektóre rodzaje
błędów będą częstsze niż inne [Zaccolo i in. 1996, Cirino i in. 2003]. Rozwiązanie tego
problemu można uzyskać przez zastosowanie specjalnej polimerazy przeznaczonej właśnie do reakcji EP-PCR. Polimeraza Mutazyme II DNA pozwala na ominięcie problemu preferencji spowodowanej tempem mutacji nukleotydów A i T w stosunku do G i C
[Cadwell i Joyce 1992, Rasila i in. 2009].
Kolejna przeszkoda wynika z charakteru kodu genetycznego. Pojedyncza zmiana nukleotydu nie jest w stanie doprowadzić do zmiany kodowanego aminokwasu we wszystkich możliwych przypadkach. Na przykład pojedyncza mutacja w kodzie dla waliny
prowadzi do kodowania fenyloalaniny, leucyny, izoleucyny, alaniny, asparaginy lub glicyny. W celu uzyskania pozostałych aminokwasów potrzebne są dwie mutacje punktowe
(K, S, P, H, R, N, T, F, E, Y) lub nawet trzy (Q, W, K). W wyniku tego otrzymamy bibliotekę klonów, w której niektóre aminokwasy będą rzadziej kodowane niż inne. Uważa się,
że może to być ewolucyjną próbą ochrony aminokwasów ważnych w funkcjonowaniu
białka przed utratą aktywności [Miyazaki i Arnold 1999].
Biotechnologia 15 (2) 2016
20
J. Kustosz
Następnym problemem przy wykorzystaniu techniki EP-PCR w tworzeniu biblioteki
zmutowanego genu jest fakt, że zmiany, które podczas amplifikacji nastąpią w pierwszych
etapach reakcji powielania, będą występowały częściej niż inne mutacje. Odkrycie takiego
zjawiska może być trudne do ujawnienia przy analizie sekwencji, zwłaszcza gdy biblioteka
klonów jest duża. Problem ten w pewnym stopniu można ominąć przez wykonanie kilku
EP-PCR z zamierzonymi błędami i poprzez skonstruowanie biblioteki łącznej/końcowej.
Inną możliwością uniknięcia takiej sytuacji jest zmniejszenie liczby cykli amplifikacji,
jednak to podejście jest również stosowane jako jedne z najprostszych sposobów kontrolowania stopnia mutagenezy. Najskuteczniej jest połączyć oba sposoby, zmniejszyć liczbę
cykli amplifikacji i wykonać EP-PCR w wielu powtórzeniach. Problem ten nie jest poważny w przypadku, gdy celem badania jest sprawdzenie właściwości zmienionego białka.
Jednak gdy interpretacja wyników opiera się na wnioskach statystycznych przeprowadzonego badania, może to spowodować, że nasze wyniki będą błędne [Neylon 2004].
W 2014 roku ta technika została wykorzystana w mutagenezie ludzkiego cytochromu
P450 2A6. Bierze on udział w metabolizmie ksenobiotyków oraz jest głównym enzymem
w ludzkiej wątrobie biorącym udział w utlenianiu nikotyny, kumaryny i specyficznej dla
tytoniu nitrozaminy. Celem badania było wzmocnienie katalizy enzymatycznej tego białka. Została stworzona biblioteka klonów zmutowanego genu kodującego cytochrom P450
2A6 oparta na EP-PCR, a następnie oznaczono aktywność hydroksylazy kumaryny metodą fluoroscencyjną. Wykonano cztery kolejne rundy mutagenezy i analizy przesiewowej.
W każdej rundzie zostały wyselekcjonowane klony, które były katalitycznie lepsze od
swojego pierwotnego odpowiednika. W przypadku wyselekcjonowanych mutantów wykazano następujące mutacje reszt aminokwasowych w cytochromie P450 2A6: B1 (F209S),
C1 (F209S, S369G), D1 (F209S, S369G, E277K), E1 (F209S, S369G, E277K, A10V).
Mutanty E1 wykazały około 13-krotny wzrost aktywności w teście opierającym się na
fluoroscencyjnej hydroksylazie kumaryny na poziomie całej komórki bakteryjnej. Analiza
strukturalna P450 2A6 F209S wykazała, że mutacja ta jest wystarczająca, aby wpływać na
bezpośrednie oddziaływanie z substratem w miejscu aktywnym [Lee i in. 2014].
Kolejnym przykładem wykorzystania tej techniki była próba zwiększenia tolerancji
bakterii Zymomonas mobilis na etanol. W badaniu tym kierowano się faktem, iż coraz większe znaczenie na rynku odgrywają biopaliwa pochodzące z odnawialnych źródeł energii,
a produkują je mikroorganizmy, których tolerancja na etanol jest czynnikiem limitującym.
Mutowano gen kodujący czynnik transkrypcyjny, jakim jest białko RpoD. Wykorzystano
w tej próbie mutagenezy technikę EP-PCR i polimerazę Mutazyme II DNA. Wyizolowano
cztery mutanty z biblioteki klonów, których tolerancja na etanol była wyższa niż w przypadku szczepu kontrolnego. To doświadczenie dowiodło, że mutacja w genie RpoD może
prowadzić do zwiększenia tolerancji Z. mobilis na etanol [Tan i in. 2016].
Technika EP-PCR jest również wykorzystywana do tworzenia enzymów działających
w niskich temperaturach. Zimne enzymy mogą mieć zastosowanie w detergentach takich
jak proszki do prania, w przemyśle spożywczym czy też bioremediacji. Jedną z grup
takich enzymów są lipazy. Są one postrzegane jako białka bezpieczne i mają duże zastosowanie w przemyśle. W 2016 roku została podjęta próba obniżenia optimum temperatury tego enzymu. W tym celu wykorzystano gen LipJ pochodzący z bakterii z rodzaju
Bacillus. Wyselekcjonowano jeden szczep z mutacją F19L (zamiana fenyloalaniny do
leucyny). W ten sposób otrzymano enzym, który miał swoje optimum już w temperaturze
10oC, podczas gdy wyjściowy enzym miał optimum w 37oC [Goomber i in. 2016].
Acta Sci. Pol.
21
Techniki mutagenezy losowej...
ERROR-PRONE ROLLING CIRCLE AMPLIFICATION (RCA)
Inną techniką wykorzystywaną w mutagenezie jest amplifikacja według mechanizmu toczącego się koła z zamierzonymi błędami nazwana RCA (ang. error-prone rolling circle
amplification) (rys. 5). W technice tej amplifikacji ulega cały plazmid lub wirusowy DNA
w warunkach PCR z zamierzonymi błędami [Fujii i in. 2006]. Metoda ta składa się tylko
z jednego etapu amplifikacji DNA poprzez transformację szczepu gospodarza. Eliminuje się tu potrzebę użycia enzymów restrykcyjnych i ligazy DNA. Ta technika pozwala
uzyskać częstość mutacji na poziomie 3–4 na 1 kpz. RCA ma kilka zalet w porównaniu
z wcześniej opisywanym EP-PCR.
• Nie są potrzebne w tym przypadku specyficzne startery, wystarczą losowe heksamery
uniwersalne do każdej matrycy [Dean i in. 2001, Fujii i in. 2004].
• Nie wymaga również cykli termicznych, i specjalnych urządzeń (termocykler) ani
żadnych enzymów restrykcyjnych czy ligazy [Fujii i in. 2004].
• Kolisty produkt amplifikacji RCA może być klonowany wprost do bakterii lub drożdży i bezpośrednio stosowany do transformacji szczepu gospodarza [Vakulenko i in.
1999, Ding i in. 2003].
Metoda ta jest uważana za jedną z najprostszych metod mutagenezy losowej (rys. 6). Do
amplifikacji potrzebne są jedynie: wektor kolisty (np. pUC19), heksamery używane jako
niespecyficzne startery, polimeraza φ29 DNA oraz warunki, w których polimeraza zwiększy swój poziom błędów, np. przez zwiększenie w reakcji stężenia jonów manganu [Fujii i
in. 2004]. Reakcja RCA przebiega w warunkach izotermicznych. Do tego procesu została
wybrana polimeraza φ29 DNA, ponieważ ma ona zdolność syntezy fragmentów dłuższych
niż 70 000 pz i pozwala na ciągłą syntezę. Dodatkowo, zastosowanie zmodyfikowanej
polimerazy DNA φ29 bez aktywności egzonukleazy 3’–5’ może zwiększyć częstości mutacji. Wprowadzenie mutacji H61R w polimerazie φ29 DNA powoduje 16-krotny wzrost
wydajności błędnego wprowadzania zasad i 6–23-krotnego zwiększenia wydajności polimeryzacji [de Vega i in. 2000]. Częstość mutacji może być również kontrolowana przez
zmianę ilości jonów manganu. Jednak ich stężenie nie powinno przekraczać 2 mM, ponieważ nadmiar MnCl2 powoduje spadek wydajność RCA [Fujii i in. 2004].
Przyłączenie się losowych
heksametrów do matrycy DNA
w wielu miejscach
Random hexamer primers
anneal to the template DNA
at many sites
Wydłużanie przez pilimerazę
DNA każdego ze starterów
DNA polymerase extend each
of these primers
losowe krótkie startery – random hexamer
polimeraza φ29 DNA – phi29 DNA polymerase
Rys. 5. Schemat przebiegu techniki RCA (modyfikacja: URL4)
Fig. 5. Scheme of technology RCA (modification: URL4)
Biotechnologia 15 (2) 2016
Powstawanie nowych miejsc przełączenia
dla starterów na nowo syntetyzowanej nici
DNA i ich wydłużanie
The formation new sites for primer annealing in the newly synthesized DNA strands
and extend them
22
J. Kustosz
W 2004 roku po raz pierwszy przeprowadzono badania w celu sprawdzenia wydajności tej metody mutagenezy. Stwierdzono, że w obecności jonów manganu częstość mutacji może wynieść około 4 nukleotydy na tysiąc par zasad. Częstotliwość ta odpowiada
1 mutacji aminokwasowej na tysiąc par zasad. Podczas badania różnorodności mutacji
okazało się, że częstość występowania tranzycji (66%) jest większa od transwersji. Może
to powodować, że przy użyciu polimerazy φ29
29 DNA uzyskamy inne mutacje, niespotykane w tradycyjnym PCR z zamierzonymi błędami. Aby stwierdzić, że metoda ta może
być skuteczna w przypadku zmiany właściwości enzymatycznych, wykorzystano ją do
zwiększenia odporności na ceftazydym poprzez modyfikację genu kodującego TEM-1
β-laktamazę. W tym badaniu udało się zidentyfikować 7 szczepów bakterii ze zwiększoną odpornością na ceftazydym. Wszystkie mutanty charakteryzowały się zmianami
w dwóch pozycjach: R164 (H, G lub C) lub D179 (do G) [Fujii i in. 2004]. Oba te aminokwasy znajdują się u nasady pętli Ω TEM-1 β-laktamazy, a zmiany w tych miejscach
znane są z poprawy odporności na ceftazydym [Vakulenko i in. 1995, 1999].
W 2011 roku naukowcy pod kierownictwem Tuomasa Huovinena poprawili wydajność reakcji RCA. Ponieważ jedną z głównych wad tego procesu jest niska sprawność
produkcji konkatamerów DNA podczas amplifikacji, badacze wprowadzili do docelowych plazmidów dodatkowe miejsce rekombinacji loxP bakteriofaga P1 rozpoznawanych przez rekombinazę Cre. Zwiększyło to ilość trasformantów 50-krotnie w warunkach
normalnych i 13-krotnie w warunkach z zamierzonymi błędami [Huovinen i in. 2011].
Jednym z przykładów wykorzystania tego typu mutagenezy losowej była próba poprawy aktywności celulazy. Naukowcy z Wietnamu użyli techniki RCA z dodatkiem 1,7
mM MnCl2. Zostały wprowadzone losowe mutacje w genie endoglukanazy. Sześć wyselekcjonowanych szczepów wykazało wyższą aktywność w porównaniu ze szczepem
typu dzikiego. Poddano sekwencjonowaniu zmutowane geny i okazało się, że znaleziono
pięć zmian w sekwencji nukleotydowej prowadzących do następujących zmian w aminokwasach: A15T, P24A, P26Q, G27A i E289V. Wśród tych pięciu podstawień określono
E289V jako odpowiedzialny za zwiększoną aktywność enzymu. Obserwacja ta została
potwierdzona ukierunkowaną mutagenezą [Van Hanh i Kim 2012].
WYKORZYSTANIE ZMUTOWANYCH SZCZEPÓW W MUTAGENEZIE
Błędy w replikacji materiału genetycznego mogą zajść spontanicznie, w każdej komórce. Jednak niektóre geny kodują białka, które spełniają funkcję systemów naprawczych
błędnie sparowanych nukleotydów [Brown 2009]. Wprowadzenie zmian w genach kodujących takie białka może spowodować zwiększenie częstości mutacji. Utworzono szczepy zawierające mutacje w genach ich systemów naprawczych, co prowadzi do wzrostu
spontanicznych mutacji. W przypadku szczepu E. coli XL1-Red wprowadzono zmiany
w trzech niezależnych ścieżkach naprawczych DNA [Greener i in. 1997]. Są to geny:
• mutS – kodujący białko odpowiedzialne za skorygowanie mutacji punktowych insercji/delecji podczas replikacji DNA [Nag i in. 2007];
• mutT – ważny czynnik w uniknięciu błędów replikacyjnych, jego inaktywacja powoduje zwiększenie błędów transwersyjnych A.T-->C.G, uniemożliwia hydrolazę
8-oxodGTP [Fowler i Schaaper 1997];
Acta Sci. Pol.
23
Techniki mutagenezy losowej...
mutD – gen ten koduje jedną z podjednostek polimerazy DNA III u E. coli, jest jednym z głównych białek kompleksu polimeraza – DNA, działa jako korekta endonukleazy, która usuwa nieprawidłowo włączone nukleotydy podczas replikacji DNA
[Scheuermann i in. 1983].
W tak skonstruowanym szczepie E. coli zaobserwowano 500-krotny wzrost częstości mutacji niż w przypadku szczepu wyjściowego. Pozwala to na uzyskanie zmian
w klonowanym genie już po jednodniowym wzroście takiego szczepu. Taki wybór metody klonowania ma jednak swoje wady. Ponieważ mutacje mogą zajść też w innych
genach komórki, nie są ograniczone tylko do wektora, w którym wprowadziliśmy interesujący nas gen [Greener i in. 1997].
Użycie tej techniki mutagenezy dla genu BetL z Listerii monocytogenes spowodowało
wzrost odporności mikroorganizmu na stres osmotyczny. Pozwala to na hodowanie Listerii w buforze tryptofanowym ze zwiększoną osmolalnością [Hoffmann i in. 2013].
Innym przykładem próby mutagenezy wykorzystującej to podejście była próba zmiany aktywności beta-glukuronidazy kodowanej prze gen gusA. Gen ten jest wykorzystywany w biologii molekularnej jako gen reporterowy. Jego maksymalna aktywność została zidentyfikowana w pH 5, natomiast neutralne pH limituje jego działanie. W celu
zmiany jego optymalnego pH i zmodyfikowania białka zastosowano szczep XL1-Red. Zidentyfikowano dwa allele gusA2 i gusA3 ze zwiększoną aktywnością beta-glukuronidazy
w neutralnym pH. Jeden z nich gusA3 wykazywał wysoką aktywność w przedziale
pH 4–8. Analiza sekwencji nukleotydowych gusA2 i gusA3 wykazała zmiany pojedynczych nukleotydów powodujących substytucję aminokwasów D524G i D573A. Zmodyfikowany gen gusA3 może być używany jako potencjalny enzym reporterowy do transformacji komórek, które nie są acidofilne [Callanan i in. 2007].
•
RCA z zamierzonymi błędami
Error-prone RCA
PCR z zamierzonymi błędami
Error-prone PRC
zmutowany szczep
transformacja zmutowanego
szczepu
transformation of mutator
strain
trawienie enzymami restrykcyjnymi
digestion with restriction enzymes
separacja fragmentów
separation of fragments
ligacja
ligation
transformacja
transformation
transformacja
transformation
transformacja
transformation
Rys. 6. Porównanie metod mutagenezy losowej wykorzystujących technikę transformacji komórek gospodarza (modyfikacja: Fujii i in. 2004)
Fig. 6. Comparison of methods random mutagenesis technique for transformed host cells (modification: Fujii i in. 2004)
Biotechnologia 15 (2) 2016
24
J. Kustosz
TASOWANIE DNA (ANG. DNA SHUFFING)
Gdy została uzyskana już biblioteka klonów zmutowanego białka i otrzymaliśmy kilka
mutantów, które wykazują wyższe zdolności niż to było w przypadku wariantu wyjściowego, to istnieje możliwość wymiany fragmentów DNA takich klonów. Możliwe jest
sprawdzenie, czy połączenie kilku pozytywnych zmian dodatkowo wzmocni aktywność
enzymatyczną. Ta technika nazywana jest tasowaniem DNA i jest potężnym narzędziem
w inżynierii białek. Może być ona stosowana również w przypadku tasowania DNA homologicznych genów [Stemmer 1994]. Metoda ta przebiega w kilku etapach. Na początku homologiczne geny są trawione DNazą, która dzieli materiał genetyczny ma mniejsze
fragmenty długości około 50–100 pz. Fragmenty niestrawione są usuwane. Następnie
fragmenty są łączone ze sobą za pomocą PCR bez starterów. W kolejnym etapie uzyskane
fragmenty są aplikowane już ze specyficznymi starterami i wklonowywane do wektora
[Stemmer 1994]. Ta metoda może być również użyta ze specyficznymi enzymami restrykcyjnymi z grupy II. W tym przypadku homologiczne geny są najpierw trawione
enzymem restrykcyjnym, a w kolejnym następuje ligacja fragmentów [Engler i in. 2009].
Dodatkowo metodę tę można przeprowadzić w warunkach error-prone, co zwiększy poziom błędów podczas amplifikacji [Zhao i Arnold 1997].
W jednym z pierwszych badań wykorzystujących tasowania DNA Stemmer w roku
1994 użył tej techniki w celu zwiększenia zdolności TEM-1 p-laktamazy do przekazania
oporności bakterii na antybiotyk cefotaksym [Stemmer 1994]. Tasowanie przeprowadzono w celu wymieszania mutacji punktowych zgromadzonych w czasie amplifikacji PCR.
Najbardziej aktywne mutanty otrzymane wykazywały 32 000-krotny wzrost aktywności,
ale stwierdzono jedynie 6 mutacji punktowych w porównaniu z macierzystym białkiem.
Takie podejście generuje niewiele mutacji punktowych [Stemmer 1994, Stevenson i Benkovic 2002].
W 2016 roku wykorzystano tę metodę mutagenezy do stworzenia skuteczniejszej
szczepionki przeciwko szerokiej gamie zakażeń bakteryjnych [Li i in. 2016]. W tym celu
tasowano homologiczny gen OmpA (białko błony zewnętrznej) z kilku szczepów chorobotwórczych. Uzyskano dwie poliwalentne szczepionki.
Jedną z odmian tasowania DNA jest technika ang. random priming recombination
(RPR). W tym przypadku zamiast użycia DNazy I do generowania krótkich fragmentów
DNA, wykorzystano losowe startery DNA. Uzyskane fragmenty mogą zawierać mutacje
punktowe. Ta technika ma kilka zalet: wymagane są tylko małe ilości matrycy DNA,
do generacji fragmentów nie jest potrzebna DNaza I, którą trzeba usuwać w późniejszych etapach tasowania DNA oraz syntezowane losowe startery są niezależne od długości DNA. Dzięki tej metodzie zwiększono nawet 8-krotnie okres półtrwania szczepu
z gatunku Bacillus subtilis w temperaturze 65°C, doprowadzając do mutacji genów RC1
i RC2 kodujących subtilizynę E [Shao i in. 1998].
W kolejnej metodzie ang. staggered extension process (StEP) wykorzystano inne
podejście. W tym przypadku wyeliminowano użycie fragmentów genu, a zamiast tego
podmieniano matrycę DNA, na której była budowana himeryczna sekwencja. W tym
procesie określanym jako stopniowe rozszerzanie synteza nowej nici DNA następuje
w bardzo krótkim czasie, tak że każdy nowo syntezowany fragment DNA zmienia matrycę wielokrotnie. Zaletą tej metody jest również to, że nie trzeba używać DNazy I oraz
Acta Sci. Pol.
25
Techniki mutagenezy losowej...
potrzebna jest niewielka ilość DNA. Wykorzystując to podejście, uzyskano 50-krotny
wzrost termostabilności Bacillus subtilis w temperaturze 65°C [Zhao i in. 1998].
Ostatnią już metodą, która opiera swoje podstawy na tasowaniu DNA jest ang. random chimeragenesis on transient templates (RACHITT) (rys. 7). Na pierwszym etapie tej
techniki jedna nić rodzicielska jest używana jako matryca, zostaje ona wytworzona z uracylem (dUTP), pozostałe są dzielone za pomocą DNazy I. Następnie powstałe fragmenty
i matrycę łączy się na zasadzie komplementarności. Niezwiązane ogonki są trawione
egzonukleazą, luki są wypełniane przez polimerazę DNA, a pęknięcia łączone przez ligazę. Nić dUTP jest usuwana przez trawienie endonukleazą V. Jednoniciowa chimera
jest kopiowana na dsDNA [Coco i in. 2001]. W tej metodzie unika się problemu zbytniego rozproszenia genów rodzicielskich w niedopuszczalnie wysokich częstotliwościach
w końcowej bibliotece [Coco, 2003].
Tasowanie DNA – DNA shuffling
geny wyjściowe – rodzicielskie
parent sequence
fragmentacja
fragmentation
StEP
geny wyjściowe – rodzicielskie
parent sequence
RACHITT
geny wyjściowe – rodzicielskie
parent sequence
nić rodzicielska służąca
jako matryca DNA z dUTP
bottom strand scaffold dUTP
wydłużanie starterów
terminal primed
synthesis
hybrydyzacja
hybridization
zamiana matrycy DNA
template switching
składanie
reassemby
zamiana matrycy DNA
template switching
klonowanie i selekcja
cloning and selection
klonowanie i selekcja
cloning and selection
trawienie niezwiązanych ogonków,
wypełnianie luk przez polimerazę
i łączenie pęknięć przez ligazę
digest flaps, fill gaps and ligate nicks
usunięcie matrycy DNA
i stworzenie drugiej nici DNA
desrtoy scaffolding,
make double stranded
klonowanie i selekcja
cloning and selection
Rys. 7. Porównanie metod tasowania DNA (modyfikacja: Coco i in. 2001, Stevenson i Benkovic
2002)
Fig. 7. Comparison methods of DNA shuffling (modification: Coco i in. 2001, Stevenson i Benkovic 2002)
PODSUMOWANIE
Obecny rozwój inżynierii genetycznej stwarza nam ogromne możliwości do usprawnienia działania enzymów poprzez testowanie wprowadzanych zmian aminokwasów. Dzięki
zaprezentowanym technikom losowym wykorzystywanym w mutagenezie niezależnie
od znajomości sekwencji nukleotydowej i aminokwasowej białek możemy otrzymać mutanty (lub zmutowane geny) o ulepszonych właściwościach. Jest to jedna z głównych
zalet w porównaniu z metodami ukierunkowanymi na konkretne zmiany (mutageneza
ukierunkowana). Techniki te pozwalają nam na badanie zależności strukturalno-funkcjonalnych białek i wpływanie na ich aktywność. Za główną wadę można uznać czasochłonną selekcję mutantów, która może nie wykazać oczekiwanych przez nas rezultatów.
Biotechnologia 15 (2) 2016
26
J. Kustosz
PIŚMIENNICTWO
Balci H., Ozturk M.T., Pijning T., Ozturk S.I., Gumusel F., 2014. Improved activity and pH stability of E. coli ATCC 11105 penicillin acylase by error-prone PCR. Applied Microbiology and
Biotechnology, 98 (10), 4467–4477.
Bessler C., Schmitt J., Maurer K.H., Schmid R.D., 2003. Directed evolution of a bacterial alpha-amylase: toward enhanced pH-performance and higher specific activity. Protein Sci., 12, 2141–2149.
Brandenburger A., Godson G.N., Radman M., Glickman B.W., Van Sluis C.A., Doubleday
O.P., 1981. Radiation-induced base substitution mutagenesis in single-stranded DNA phage
M13. Nature, 294, 180–182.
Cadwell R.C., Joyce G.F., 1992. Randomization of genes by PCR mutagenesis. Genome Research, 2 (1), 28–33.
Cadwell R.C., Joyce G.F., 1994. Mutagenic PCR. PCR Methods Appl., 3, S136–S40.
Callanan M.J., Russell W.M., Klaenhammer T.R., 2007. Modification of Lactobacillus
β-glucuronidase activity by random mutagenesis. Gene, 389 (2), 122–127.
Cirino P.C., Mayer K.M, Umeno D., 2003. Generating mutant libraries using error-prone PCR.
Methods Mol. Biol., 231, 3–9.
Coco W.M., 2003. Gene Family Shuffling by Random Chimeragenesis on Transient Templates. Directed Evolution Library Creation: Methods and Protocols, 231, 111.
Coco W.M., Levinson W.E., Crist M.J., Hektor H.J., Darzins A., Pienkos P.T., Squires C.H., Monticello D.J., 2001. DNA shuffling method for generating highly recombined genes and evolved
enzymes. Nature Biotechnology, 19 (4), 354–359.
de Vega M., Lazaro J. M., Salas M., 2000. Phage φ29 DNA polymerase residues involved in the proper
stabilisation of the primer-terminus at the 3′–5′ exonuclease active site. J. Mol. Biol., 304, 1–9.
Dean F.B., Nelson J.R., Giesler T.L., Lasken R.S. 2001. Rapid amplification of plasmid and phage
DNA using Phi 29 DNA polymerase and multiply-primed rolling circle amplification. Genome
Res., 11, 1095–1099.
Ding X., Snyder A.K., Shaw R., Farmerie W.G., Song W.Y., 2003. Direct retransformation of yeast
with plasmid DNA isolated from single yeast colonies using rolling circle amplification. BioTechniques, 35, 774–779.
Engler C., Gruetzner R., Kandzia R., Marillonnet S., 2009. Golden gate shuffling: a one-pot DNA
shuffling method based on type IIs restriction enzymes. PloS one, 4 (5), e5553.
Ermakova-Gerdes S., Shestakov S., Vermaas W., 1996. Random chemical mutagenesis of a specific
psbDI region coding for a lumenal loop of the D2 protein of photosystem II in Synechocystis
sp. PCC 6803. Plant molecular biology, 30 (2), 243–254.
Fowler R.G., Schaaper R.M., 1997. The role of the mutT gene of Escherichia coli in maintaining
replication fidelity. FEMS Microbiology Reviews, 21(1), 43–54.
Fujii R., Kitaoka M., Hayashi K., 2004. One-step random mutagenesis by error-prone rolling circle
amplification. Nucleic Acids Research, 32 (19), e145–e145.
Fujii R., Kitaoka M., Hayashi K., 2006. Error-prone rolling circle amplification: the simplest random mutagenesis protocol. Nature Protocols, 1 (5), 2493–2497.
Goomber S., Kumar A., Kaur, J. 2016. Disruption of N terminus long range non covalent interactions shifted temp. opt 25°C to cold: Evolution of point mutant Bacillus lipase by error prone
PCR. Gene, 576 (1), 237–243.
Gopinath K.P., Murugesan S., Abraham J., Muthukumar K., 2009. Bacillus sp. mutant for improved
biodegradation of Congo red: random mutagenesis approach. Bioresource Technology, 100
(24), 6295–6300.
Greener A., Callahan M., Jerpseth B., 1997. An efficient random mutagenesis technique using
an E. coli mutator strain. Molecular Biotechnology, 7 (2), 189–195.
Acta Sci. Pol.
Techniki mutagenezy losowej...
27
Hoffmann R.F., McLernon S., Feeney A., Hill C., Sleator R.D., 2013. A single point mutation
in the listerial betL σA-dependent promoter leads to improved osmo-and chill-tolerance and
a morphological shift at elevated osmolarity. Bioengineered, 4 (6), 401–407.
Huovinen T., Julin M., Sanmark H., Lamminmäki U., 2011. Enhanced error-prone RCA mutagenesis by concatemer resolution. Plasmid, 66 (1), 47–51.
Labrou N.E., 2010. Random mutagenesis methods for in vitro directed enzyme evolution. Current
Protein and Peptide Science, 11 (1), 91–100.
Lee H., Kim J.H., Han S., Lim Y.R., Park H.G., Chun Y.J., Park W.S, Kim D., 2014. Directed-Evolution Analysis of Human Cytochrome P450 2A6 for Enhanced Enzymatic Catalysis. Journal
of Toxicology and Environmental Health, Part A, 77 (22–24), 1409–1418.
Li H., Chu X., Li D., Zeng Z.H., Peng X.X., 2016. Construction and immune protection evaluation
of recombinant polyvalent OmpAs derived from genetically divergent ompA by DNA shuffling. Fish & Shellfish Immunology, 49, 230–236.
Little J.B., Nagasawa H., Pfenning T., Vetrovs H., 1997. Radiation-induced genomic instability: delayed mutagenic and cytogenetic effects of X rays and alpha particles. Radiation Research, 148
(4), 299–307.
Miyazaki K., Arnold F.H., 1999. Exploring nonnatural evolutionary pathways by saturation mutagenesis: rapid improvement of protein function. Journal of Molecular Evolution, 49, 716–720.
Mohan U., Banerjee U.C., 2008. Molecular evolution of a defined DNA sequence with accumulation of mutations in a single round by a dual approach to random chemical mutagenesis (DuARCheM). Chembiochem, 9 (14), 2238–2243.
Nag N., Rao B.J., Krishnamoorthy G., 2007. Altered dynamics of DNA bases adjacent to a mismatch: a cue for mismatch recognition by MutS. J. Mol. Biol., 374 (1), 39–53.
Neylon C., 2004. Chemical and biochemical strategies for the randomization of protein encoding DNA sequences: library construction methods for directed evolution. Nucleic Acids Research, 32 (4), 1448–1459.
Rasila T.S., Pajunen M.I., Savilahti H., 2009. Critical evaluation of random mutagenesis by errorprone polymerase chain reaction protocols, Escherichia coli mutator strain, and hydroxylamine
treatment. Analytical Biochemistry, 388 (1), 71–80.
Ribeiro O., Magalhães F., Aguiar T.Q., Wiebe M.G., Penttilä M., Domingues L., 2013. Random
and direct mutagenesis to enhance protein secretion in Ashbya gossypii. Bioengineered, 4 (5),
322–331.
Rosenbluth R.E., Cuddeford C., Baillie, D.L., 1985. Mutagenesis in Caenorhabditis elegans. II.
A spectrum of mutational events induced with 1500 r of γ-radiation. Genetics, 109 (3), 493–511.
Scheuermann R., Tam S., Burgers P.M., Lu C., Echols H., 1983. Identification of the epsilon-subunit of Escherichia coli DNA polymerase III holoenzyme as the dnaQ gene product: a fidelity
subunit for DNA replication. Proceedings of the National Academy of Sciences of the United
States of America Dec., 80 (23), 7085–7089.
Selifonova O., Valle F., Schellenberger V., 2001. Rapid evolution of novel traits in microorganisms. Applied and Environmental Microbiology, 67 (8), 3645–3649.
Shao Z., Zhao H., Giver L., Arnold F.H., 1998. Random-priming in vitro recombination: an effective tool for directed evolution. Nucleic Acids Research, 26 (2), 681–683.
Sobkowiak L., Szweykowska-Kulinska Z., 2007. Fizyczne, chemiczne i genetyczne metody mutagenezy roślin. Biotechnologia, 4, 157–169.
Stemmer W.P., 1994. DNA shuffling by random fragmentation and reassembly: in vitro recombination for molecular evolution. Proceedings of the National Academy of Sciences, 91 (22),
10747–10751.
Stevenson J.D., Benkovic S.J., 2002. Combinatorial approaches to engineering hybrid enzymes. Journal of the Chemical Society, Perkin Transactions 2, (9), 1483–1493.
Biotechnologia 15 (2) 2016
28
J. Kustosz
Tan F., Wu B., Dai L., Qin H., Shui Z., Wang J., Zhu Q., Hu G., He M., 2016. Using global transcription machinery engineering (gTME) to improve ethanol tolerance of Zymomonas mobilis. Microbial Cell Factories, 15 (1), 1–9.
Tanooka H., Munakata N., 1978. Mutagenic effect of tritiated water on spores of Bacillus subtilis. Radiation Research, 73 (3), 581–584.
Tessman I., Poddar R.K., Kumar S., 1964. Identification of the altered bases in mutated singlestranded DNA: I. In vitro mutagenesis by hydroxylamine, ethyl methanesulfonate and nitrous
acid. Journal of Molecular Biology, 9 (2), 352–363.
URL1:
http://4.bp.blogspot.com/-D0f-VsqHIUQ/UjQxnRyfkaI/AAAAAAAABzw/fflvfi7ICu8/
s1600/EMS.png
URL2:
http://2012.igem.org/wiki/images/thumb/0/08/Mutagenesis_library_selection_system2.
JPG/900px-Mutagenesis_library_selection_system2.JPG
URL3: http://spot.pcc.edu/~jvolpe/b/bi234/lec/hw/thymineDimer.jpg
URL4: http://www.mclab.com/images/Products/MCRCA%20DNA%20Amplification%20Kit%20
figure%201.png
Vakulenko S.B., Taibi-Tronche P., Tóth M., Massova I., Lerner S.A., Mobashery S., 1999. Effects on substrate profile by mutational substitutions at positions 164 and 179 of the class A
TEMpUC19 β-lactamase from Escherichia coli. Journal of Biological Chemistry, 274 (33),
23052–23060.
Vakulenko S.B., Toth M., Taibi P., Mobashery S. Lerner S.A., 1995 Effects of Asp-179 mutations
in TEMpUC19 β-lactamase on susceptibility to β-lactams. Antimicrob. Agents Chemother., 39,
1878–1880.
Vakulenko S.B., Taibi-Tronche P., Toth M., Massova I., Lerner S.A. Mobashery S., 1999. Effects on substrate profile by mutational substitutions at positions 164 and 179 of the class A
TEMpUC19 β-lactamase from Escherichia coli. J. Biol. Chem., 274, 23052–23060.
Van Hanh V., Kim K., 2012. Improvement of cellulase activity using error-prone rolling circle
amplification and site-directed mutagenesis. Journal of Microbiology and Biotechnology,
22 (5), 607–613.
Zaccolo M., Williams D.M., Brown D.M., Gheradi E., 1996. An approach to random mutagenesis
of DNA using mixtures of triphosphate derivatives of nucleoside analogues. J. Mol. Biol., 255,
589–603.
Zhao H., Arnold F.H., 1997. Optimization of DNA shuffling for high fidelity recombination. Nucleic
Acids Research, 25 (6), 1307–1308.
Zhao H., Giver L., Shao Z., Affholter J.A., Arnold F.H., 1998. Molecular evolution by staggered
extension process (StEP) in vitro recombination. Nature Biotechnology, 16 (3), 258–261.
Acta Sci. Pol.
Techniki mutagenezy losowej...
29
RANDOM MUTAGENESIS TECHNIQUES RESULTING IN INCREASED
POLYMORPHISM
Abstract. Protein engineering is now a very important sector in modern biotechnology.
They are used random mutagenesis, which allows production enzymes and proteins with
the desired characteristics or increased activity. These techniques and the appropriate method of bacterial selection permit to accelerate the evolution of genomes for industry. There
are many techniques to create random changes in the amino acid sequence of the protein.
This can be achieved by treating the whole bacteria or DNA chemicals or UV radiation,
using modified techniques of PCR or RCA in error-prone mix, mutator strains and using
one of the methods of DNA shuffling. The following article provides a description of these
methods and examples of successful use of them.
Key words: physical and chemical mutagenesis, EP-PCR, RCA, mutator strain, DNA shuffling, StEP, RACHITT
Zaakceptowano do druku – Accepted for print: 30.06.2016
Do cytowania – For citation: Kustosz J., 2015. Techniki mutagenezy losowej prowadzącej
do zwiększenia polimorfizmu. Acta Sci. Pol. Biotechnol., 15 (2), 15–30.
Biotechnologia 15 (2) 2016