Rozdział 19 · ODPORNOŚĆ ·
raka piersi, przeciwciała anty-GPIIb/IIIa (abciximab) w zapobieganiu nawrotom zwężenia naczyń
wieńcowych.
Nadzór immunologiczny
Komórki T porozumiewają się
z komórkami B poprzez wzajemne
oddziaływania ligand-receptor
Różnorodność zarodkowa receptorów komórek T
(TCR) znacznie przekracza różnorodność osiąganą
przez immunoglobuliny. Zjawisko to w znacznej części odzwierciedla rozmiar różnorodności połączeń
TCR uzyskanej dzięki przeniesieniu przez terminalną
transferazę deoksynukleotydylową (TdT – terminal
deoxynucleotidyl transferase) tzw. regionów N o różnej długości do połączeń międzygenowych VDJ. TdT
jest modyfikującym DNA enzymem markerowym
dla limfocytów niebędących komórkami B, wykorzystywanym w diagnostyce hematologicznej. Kal17
kulacje wskazują, że możliwe jest istnienie aż 10
różnych heterodimerów TCR, co znacznie przewyższa wartość wyliczoną dla przeciwciał. Jednakże
TCR nie podlegają hipermutacjom somatycznym po
ekspozycji na antygen, co zapobiega rozrostom klonalnym autoreaktywnych klonów T. Ponieważ
dojrzewanie powinowactwa nie wpływa na szerzenie się klonów TCR, zasadniczo TCR wiążą antygen ze znacznie mniejszym powinowactwem
-5
(około 10 M) niż uzyskiwane przez dojrzałe przeciwciała (których powinowactwo może osiągać
-10
wielkość rzędu 10 M).
Odpowiedź immunologiczna na antygen obejmuje na ogół cztery etapy komórkowe: adhezja immunocytu, rozpoznanie antygenu, kostymulacja komórek B i T, proliferacja immunocytu. Wychwycenie
antygenu przez immunoglobulinę powierzchniową
prowadzi pierwotnie do internalizacji antygenu, a
następnie jego procesowania endosomalnego. Powstałe w wyniku degradacji antygenu fragmenty
peptydowe są składane z cząsteczkami MHC klasy
II transportującymi antygeny do powierzchni komórki, gdzie są prezentowane komórkom T ekspresującym receptory o swoistym powinowactwie antygenowym. Tak zwane komórki pomocnicze T pomagają aktywować komórki B na drodze dwukierunkowej. Aktywowane komórki B oddziałują zwrotnie,
zmieniając w obecności antygenu komórkę T. Komunikacja pomiędzy pomocniczymi komórkami T a
komórkami B jest zapewniana przez dwa zestawy
sygnałów:
1. Receptor CD40 komórek B i ligand CD40L komórek T
§ umożliwia komórkom B wspomaganie przez
aktywowane komórki T.
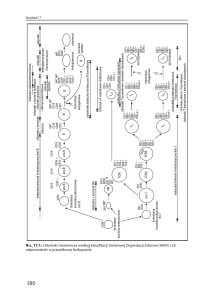
Ryc. 19.4. Dojrzewanie powinowactwa receptora komó-
rek T napędza różnicowanie podtypów komórek T. Wzór
wydzielania limfokin związanych z funkcjonalnymi podtypami komórek T został pokazany poniżej. IFNg – interferon-g; IL – interleukina; TCR – receptor komórek T,
TNF – nowotworowy czynnik nekrotyczny.
2. Receptor CD28 limfocytów TH i ligand B7
(CD80) komórek B
4+
§ umożliwia komórkom T CD odpowiadanie
aktywowanym komórkom B poprzez wydzielanie limfokin różniących się w zależności od
podtypu komórek T pomocniczych (ryc. 19.4):
­ aktywowane komórki TH1 wydzielają TNFa,
IL-2, IFNg oraz IL-12,
­ aktywowane komórki TH2 wydzielają
IL-5, IL-4, IL-3 (± IL-6, IL-10, IL-9).
Elementem łączącym obie kaskady jest CD28,
homolog ICOS, indukowalna cząstka kostymulująca (inducible co-stimulatory molecule). Następujące po antygenowo-swoistej aktywacji komórki
T związanie CD28 przez B7 prowadzi do proliferacji komórki T i wydzielenia limfokin. Jednakże zależne od antygenu wydzielanie IL-4 przez komórki B,
tworzenie centrów rozrodczych i przełączanie klas
immunoglobulin (szczególnie IgE) wymaga aktywacji ICOS przez jego antyreceptor B7RP (B7-related protein). Interesujące, że myszy pozbawione
(knockout) ICOS wykazują skłonność do autoimmunologicznej choroby demielinizacyjnej.
ASPEKTY KLINICZNE
Komórki T pomocnicze: pomagają czy
przeszkadzają?
Funkcjonalny podział komórek pomocniczych
T ekspresujących CD4 na typ 1 (TH1) i typ 2 (TH2)
497
Część IV · OD BIOLOGII MOLEKULARNEJ KOMÓRKI DO FIZJOLOGII CZŁOWIEKA ·
jest przeprowadzany nie tylko w zależności od zestawu wydzielanych limfokin, ale także na podstawie dominującego modelu aktywności immunologicznej: wyrzut prozapalnych cytokin inicjowany
przez komórki TH1 promuje głównie odpowiedź
komórkową, podczas gdy komórki TH2 silniej uruchamiają wydzielanie przeciwciał (ryc. 19.5).
Wrzodziejące zapalenie jelita grubego charakteryzuje się aktywacją głównie komórek TH2 z podwyższonym poziomem wydzielania IL-5, IL-10, i
IL-4, podczas gdy choroba Crohna (zapalenie jelita krętego/okrężnicy) jest związana ze wzrostem
wydzielania cytokin TNFa, IL-12, IL-2 i IFNg produkowanych przez komórki TH1 zapalenia śluzówki. Choroba Crohna może być leczona przez podanie IL-10 (która może być dostarczana przez probiotyki, czyli żywe mikroorganizmy skonstruowane tak, by wydzielały IL-10 do jelita) lub przez
przeciwciała neutralizujące TNFa. Z tego powodu
u myszy pozbawionych IL-10 rozwija się zapalenie jelit. Wywołane przez pasożyty podwyższenie
poziomu wydzielanej IL-10 w przewlekłym zakażeniu przywrami wykazuje ujemny związek z atopią, co wskazuje na rolę supresyjną IL-10 w stanach alergicznych.
Układ związany z komórkami TH1 jest głównym
systemem odpowiedzi immunologicznej aktywowanym w reumatoidalnym zapaleniu stawów i w
stwardnieniu rozsianym, podczas gdy system TH2
dominuje w toczniu rumieniowatym układowym i
podczas ciąży. Żeńskie hormony płciowe wzmacniają system odpowiedzi przez TH2, a wyciszają
TH1, co może wyjaśniać, dlaczego ciąża wydaje się
poprawiać stan zdrowia w reumatoidalnym zapaleniu stawów i w stwardnieniu rozsianym, czego
nie obserwuje się w przypadku tocznia rumienio-
Ryc. 19.5. Podział komórek pomocniczych T na podgru-
py TH1 i TH2. Pierwsza z nich „pomaga” głównie w odporności typu komórkowego, podczas gdy druga – w odpowiedzi humoralnej.
498
watego. Leki (np. suplatast tosilate), które hamują
wybiórczo cytokiny wydzielane przez TH2 (IL-4 i
IL-5), podobnie jak IL-12, mogą redukować eozynofilię tkankową w takich schorzeniach jak astma,
choć tego typu terapie wymagają jeszcze klinicznego potwierdzenia skuteczności.
Los komórek grasicy zależy
od czasu trwania sygnału
pochodzącego z komórek T
+
Aktywacja CD40 przez komórki CD4 prowadzi
do rozrostu klonalnego aktywowanych komórek B,
stymulacji przełączenia klas immunoglobulin, różnicowania komórek plazmatycznych syntetyzujących
właściwe przeciwciała, supresji apoptozy limfocytów B w centrach zarodkowych. Przeciwciała neutralizujące przeciwko CD40 mogą więc hamować
rozwój chorób zapalnych i autoimmunologicznych.
Wobec powyższego, wnosić można, że aktywacja
+
CD40 (na komórkach B) przez komórki T CD4 (pomocnicze) wywołuje następujące efekty:
1. Klonalny rozrost aktywowanych komórek B.
2. Stymulacja przełączenia klas immunoglobulin.
3. Różnicowanie syntetyzujących przeciwciała klonów komórek plazmatycznych.
4. Supresja apoptozy limfocytów B w centrach
rozrodczych.
Natomiast indukowana przez krzyżowe związanie antygenu do immunoglobulin powierzchniowych transdukcja sygnału jest związana z aktywacją
cytozolowej kinazy tyrozynowej Lck. Powstała fala
fosforylacji tyrozynowej prowadzi do internalizacji
kompleksów antygen-receptor z powierzchni komórki. Taka kolejność przekazywania sygnału jest
podobna do tej inicjowanej przez aktywację TCR.
+
Podczas gdy pomocnicze komórki T (CD4 , TH)
poszukują komórek ekspresujących MHC-II (makrofagi i komórki B), komórki T cytotoksyczne+
-supresorowe (CD8 , TS) wiążą powszechniej występujące cząsteczki MHCI. Rozpoznanie białka
MHC jest podwójną funkcją TCR i wspomagającego
ko-receptora CD4/CD8. Czas trwania sygnału z TCR
wpływa na to, czy komórki prekursorowe grasicy dojrzewają w kierunku komórek T pomocniczych czy supresorowych: dłuższy czas (np. > 12 godzin) trwania
+
sygnału prowadzi do powstania komórek CD4 , podczas gdy krótszy czas (np. < 2 godzin) – do powsta+
nia komórek CD8 .
Dalszym efektem utworzenia kompleksu TCR-antygen jest wydzielanie przez komórki TH IL-2,
wywołującej różnicowanie komórek B do wydzielających immunoglobuliny komórek plazmatycznych. Związanie antygenu przez TCR, podobnie jak
przez immunoglobuliny powierzchniowe na komórkach B, prowadzi do fosforylacji tyrozyny i rozrostu
Rozdział 19 · ODPORNOŚĆ ·
klonalnego komórek T o identycznym powinowactwie antygenowym.
ASPEKTY MOLEKULARNE
SAP i SLAM
SLAM (signaling lymphocyte-activation molecule, CDw150) – cząsteczka sygnałowa aktywacji
limfocytów – jest limfocytarnym białkiem transmembranowym o masie 70 kDa, działającym jako
receptor dla wirusa odry. Białko SLAM jest ekspresowane zarówno przez komórki B, jak i T,
tworząc tam homotypowe aktywujące pary receptor-ligand regulujące proliferację i funkcjonowanie limfocytów obu klas.
Związana z chromosomem X choroba limfoproliferacyjna (XLP – X-linked lymphoproliferative
disease) rozwija się w wyniku mutacji typu null w
położonym na chromosomie X genie kodującym
białko związane z SLAM (SAP – SLAM-associated
protein). SAP jest białkiem wewnątrzkomórkowym zawierającym domenę SH-2, konkurującym
o miejsce wiążące SLAM, zawierającej SH-2 fosfatazy tyrozynowej SHP-2. Brak ekspresji SAP u
chorych z XLP prowadzi do zerwania „komunikacji” między komórkami T i B. To powoduje z kolei
poważne zaburzenie proliferacji komórek B w odpowiedzi na zakażenie wirusem Epsteina-Barr
(EBV). Osoby zdrowe z ostrym pierwotnym zakażeniem EBV mogą mieć do 10% obwodowych komórek B zakażonych wirusem, co prowadzi do silnej odpowiedzi ze strony limfocytów T i zniszczenia większości zakażonych EBV komórek. U chorych z XLP regulacja odpowiedzi ze strony komórek T ulega zaburzeniu na poziomie wzajemnych
oddziaływań SLAM-SAP, co prowadzi do wzmożonej odpowiedzi ze strony cytotoksycznych komórek T i rozregulowania poliklonalnej proliferacji komórek B kończącej się rozwojem chłoniaków nieziarniczych (non-Hodgkin lymphomas).
Natychmiastowe reakcje
nadwrażliwości są wywoływane
przez IgE
Termin „alergia” oznacza wachlarz klinicznych
form natychmiastowych zespołów nadwrażliwości,
w których uczestniczą IgE. Skłonności alergiczne
mogą wykształcać się jako obrona przeciwko chorobom pasożytniczym, w której IgE uruchamia wydzielanie cytokin, takich jak histamina z limfocytów
reaktywnych zwanych eozynofilami. Inne mediatory uczestniczące w reakcji nadwrażliwości obejmują leukotrieny, czynnik aktywujący płytki i chemokiny, takie jak eotaksyny 1-3.
Osoby lub rodziny mające predyspozycje do egzogennej (niepasożytniczej) zależnej od IgE odpowiedzi są określane jako atopowe. Gen uczest-
Ryc. 19.6. Modulacja odpowiedzi komórek TH2 na aler-
gen. Komórki B są przeprogramowywane tak, by produkować IgE w odpowiedzi na wydzielane przez komórki
TH2 Il-4 i IL-13, podczas gdy wydzielanie IL-3 i IL-5 angażuje odpowiednio bazofile i eozynofile.
niczący prawdopodobnie w regulacji poziomu IgE
w surowicy zmapowano w grupie chorych z astmą
atopową na chromosomie 5q31.1. Ten locus obejmuje gen IL-4; ponieważ komórki pomocnicze T
wydzielają IL-4 aktywującą komórki B, IL-4 może
wpływać na zmianę syntetyzowanych łańcuchów ciężkich immunoglobulin na klasę IgE (ryc. 19.6). Zgodnie z tym tokiem rozumowania, powodująca nabycie
nowej funkcji mutacja Gln576Arg w obrębie receptora dla IL-4 jest również związana z atopią. Innymi
loci uwikłanymi w nadwrażliwość z udziałem IgE są:
polimorfizm Ile181Leu w FceR1-b oraz polimorfizm
Gln27Glu b2-adrenoceptora (w astmie).
Jeśli alergia jest po prostu kwestią nadwrażliwości typu IgE, to dlaczego pewne osoby są uczulone
na pewne czynniki (np. indukowana przez ćwiczenia astma), a inne nie (np. uczuleniowe zapalenie
błony śluzowej nosa)? Pewną wskazówką jest
związek między atopową egzemą i polimorfizmem
zlokalizowanego na chromosomie 14 genu kodującego proteazę serynową (chymaza komórek tucznych) syntetyzowaną w skórze. Narządowo-swoista nadwrażliwość może być częściowo spowodowana polimofizmami genetycznymi wpływającymi
na czułość detekcji powszechnie występujących
antygenów.
ASPEKTY KLINICZNE
Reakcje nadwrażliwości typu późnego
Komórki T uczestniczą w reakcjach nadwrażliwości typu późnego, takich jak kontaktowe zapalenie skóry i pewne odmiany trądu. Te niezależne
od IgE (niealergiczne) reakcje nadwrażliwości
obejmują aktywację TCR, co z kolei indukuje chemotaksję raczej monocytów niż eozynofili. Nie-
499