RAPORT KOŃCOWY Z REALIZACJI ZADANIA:
„Wpływ pasz GMO na produkcyjność i zdrowotność zwierząt, transfer
transgenicznego DNA w przewodzie pokarmowym oraz jego retencję
w tkankach i produktach żywnościowych pochodzenia zwierzęcego”
CEL BADAŃ:
Celem
badań
było
określenie
wpływu
stosowania
genetycznie
zmodyfikowanych materiałów paszowych (poekstrakcyjnej śruty sojowej i śruty
kukurydzianej) w żywieniu zwierząt gospodarskich (drobiu, trzody chlewnej i
bydła) na uzyskiwane wskaźniki produkcyjne, status metaboliczny i zdrowotny
organizmu, jakość uzyskiwanych produktów oraz transfer transgenicznego DNA w
organizmie, w tym możliwość jego obecności w produktach spożywczych
pochodzenia zwierzęcego.
Główne kierunki badań były następujące:
1. Określenie efektywności materiałów paszowych GM w żywieniu różnych
gatunków i grup technologicznych zwierząt gospodarskich, co obejmowało wpływ
badanych pasz na wskaźniki produkcyjne, strawność składników pokarmowych i
jakość uzyskiwanych produktów spożywczych pochodzenia zwierzęcego.
2. Określenie wpływu materiałów paszowych GM na parametry charakteryzujące
status
zdrowotny
organizmu
zwierzęcego
(m.in.
efektywność
odpowiedzi
immunologicznej, obraz krwi, ewentualne zmiany histopatologiczne i morfologiczne
w wybranych narządów wewnętrznych).
3. Analiza pasażu transgenicznego DNA przez przewód pokarmowy.
4. Wykazanie lub wykluczenie obecności transgenicznego DNA w tkankach,
narządach oraz produktach spożywczych pochodzenia zwierzęcego (mięso, mleko,
jaja).
WYKONAWCY
Jednostką koordynującą zadanie był Instytut Zootechniki PIB w Krakowie, w którym
badania były prowadzone przez zespół w składzie:
Prof. dr hab. Sylwester Świątkiewicz – kierownik zadania i osoba odpowiedzialna za
wykonanie doświadczeń na drobiu, Prof. dr hab. Juliusz Strzetelski – osoba odpowiedzialna za
doświadczenia na bydle, dr hab. Małgorzata Świątkiewicz – osoba odpowiedzialna za
doświadczenia na trzodzie chlewnej, ponadto w realizacji zadania uczestniczyli: dr A.
Arczewska-Włosek, mgr inż. B. Brzóska, prof. dr hab. F. Brzóska, dr I. Furgał-Dzierżuk, prof.
dr hab. E. Hanczakowska, prof. dr hab. J. Koreleski, mgr inż. A. Jarocka, dr W. Korol, mgr inż.
J. Markowski, mgr inż. M. Siemińska, dr M. Twardowska i dr M. Zymon.
Jednostką współpracującą był Państwowy Instytut Weterynaryjny – PIB w Puławach,
zespół wykonawców miał następujący skład osobowy:
Prof. dr hab. Krzysztof Kwiatek – koordynator badań w PIWet-PIB oraz prof. dr hab. D.
Bednarek, dr W. Kozaczyński, mgr M. Mazur, prof. dr hab. Z. Minta, prof. dr hab. Z. Pejsak,
prof. dr hab. M. Reichert, dr Z. Sieradzki i dr G. Tomczyk.
Zakres prac realizowanych przez Państwowy Instytut Weterynaryjny - PIB obejmował
określenie wpływu stosowania pasz zmodyfikowanych genetycznie na szeroko rozumiany
status zdrowotny zwierząt gospodarskich. W tym celu w Państwowym Instytucie
Weterynaryjnym - PIB prowadzone były analizy materiału biologicznego pochodzącego od
zwierząt żywionych bez udziału lub z udziałem pasz GM. Badania te obejmowały m.in. analizy
histopatologiczne narządów wewnętrznych oraz wskaźniki morfologiczne, biochemiczne i
immunologiczne krwi. Ponadto prowadzone były analizy pod kątem obecności transgenicznego
DNA w paszach, w treści pokarmowej poszczególnych odcinków przewodu pokarmowego oraz
w wybranych tkankach i narządach.
STOSOWANE MATERIAŁY PASZOWE GM
Do badań wybrano genetycznie zmodyfikowane (GM) materiały paszowe mające
podstawowe znaczenie w żywieniu zwierząt gospodarskich, tj.:
- poekstrakcyjną śrutę sojową produkowaną z soi MON-40-3-2 (Roundup Ready)
zmodyfikowanej w kierunku tolerancji na glifosat, składnik czynny wielu herbicydów. Śruta
sojowa Roundup Ready jest dopuszczona do obrotu handlowego na terenie Unii Europejskiej i
stanowi podstawowe źródło białka paszowego w żywieniu zwierząt gospodarskich, również w
Polsce;
- śruta kukurydziana produkowana z ziarna kukurydzy Bt zmodyfikowanej w kierunku
2
odporności na żerowanie owada szkodnika z rodziny łuskoskrzydłych – omacnicę prosowiankę.
Do badań użyto odmianę MON810 (DKC 3421YG), która w niektórych krajach UE jest
uprawiana z przeznaczeniem na paszę. Śruta kukurydziana z kukurydzy MON810 jest
dopuszczona do obrotu handlowego na terenie całej UE. Ze względu na rosnący zasięg
występowania omacnicy prosowianki w Europie można przewidywać, że znaczenie tej
odmiany w żywieniu zwierząt gospodarskich będzie się zwiększać.
W celach porównawczych, do badań zabezpieczono również poekstrakcyjną śrutę
sojową i śrutę z ziarna kukurydzy pochodzące z roślin konwencjonalnych. W przypadku
kukurydzy była to odmiana DKC 3420, rodzicielska w stosunku do badanej odmiany GM,
natomiast w przypadku poekstrakcyjnej śruty sojowej – komercyjna (certyfikowana) śruta
wyprodukowana z odmian konwencjonalnych (niemodyfikowanych).
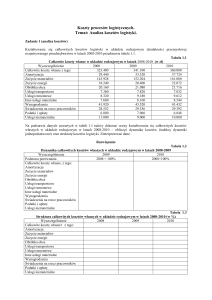
Badane materiały paszowe poddano analizom chemicznym, obejmującym analizę
podstawową, skład aminokwasowy białka, profil kwasów tłuszczowych frakcji lipidowej oraz
zawartość wybranych makro- i mikroelementów. Uzyskane wyniki wykazały, że śruta z
transgenicznej kukurydzy Bt i śruta z jej odmiany konwencjonalnej (linia rodzicielska)
charakteryzowały się podobną zawartością podstawowych składników pokarmowych i
mineralnych oraz aminokwasów (Tabela 1). Można, zatem przyjąć, że były one równoważne
pod względem wartości pokarmowej. W przypadku badanych śrut sojowych różnice były nieco
większe, jednak uzyskane wartości były nadal porównywalne i mieściły się w standardowym
zakresie przyjętym dla tego materiału paszowego. W poekstrakcyjnej śrucie sojowej i śrucie
kukurydzianej wykonano również analizę transgenicznego DNA, która potwierdziła
pochodzenie badanych materiałów.
Tabela 1.
Skład chemiczny badanej śruty kukurydzianej i poekstrakcyjnej śruty sojowej (%).
Śruta kukurydziana
Poekstrakcyjna śruta sojowa
Sucha asa
Białko ogólne
Tłuszcz surowy
Włókno surowe
Popiół surowy
Skrobia
Wapń
Fosfor
Metionina
Lizyna
Cystyna
Treonina
Tryptofan
Konwencjonalna
GM (MON 810)
Konwencjonalna
GM (MON 40-3-2)
86,2
7,67
3,40
1,88
1,30
63,7
0,010
0,287
0,149
0,235
0,159
0,269
0,040
86,3
7,75
3,45
1,87
1,24
62,8
0,010
0,283
0,151
0,252
0,161
0,266
0,044
87,7
47,9
2,04
3,66
5,70
4,08
0,272
0,644
0,642
0,289
0,623
1,75
0,515
88,6
45,7
3,22
4,28
6,31
3,54
0,353
0,701
0,754
0,292
0,700
1,85
0,763
3
Arginina
Walina
0,311
0,354
0,328
0,359
3,29
2,12
3,33
2,12
BADANIA NA KURCZĘTACH RZEŹNYCH
Doświadczenie
żywieniowe
na
kurczętach
rzeźnych
wykonano
w
kurniku
doświadczalnym Instytutu Zootechniki PIB w gospodarstwie Aleksandrowice. Do badań użyto
640 jednodniowych kurcząt Ross 308 pochodzących z wylęgarni komercyjnej. Okres
doświadczalny obejmował 42 dni (1-42 dzień życia ptaków). Okres ten odpowiada warunkom
produkcyjnym, w których młode kurczęta rzeźne odchowuje się do wieku między 35 a 42
dniem życia, kiedy osiągają 2,2-2,5 kg masy ciała. Kurczęta utrzymywano w boksach, na
ściółce z trocin, przy stałym dostępie do paszy i wody. Utworzono 4 grupy doświadczalne,
składające się z 4 powtórzeń (boksów). W każdym boksie utrzymywano 40 kurcząt.
W doświadczeniu stosowano izobiałkowe i izoenergetyczne mieszanki paszowe typu
starter (1-21 dzień życia) i grower-finiszer (22-42 dzień życia), w skład których wchodziła śruta
kukurydziana, poekstrakcyjna śruta sojowa oraz dodatki mineralne, aminokwasy krystaliczne,
olej rzepakowy i premiks witaminowo-mineralny. Zawartość składników pokarmowych i
energii metabolicznej w mieszankach była zgodna z zapotrzebowaniem kurcząt (Tabela 2 i 3).
Prawidłowość wykonania mieszanek została potwierdzona analizami obecności DNA
transgenicznego w poszczególnych dietach doświadczalnych.
Tabela 2.
Skład doświadczalnych mieszanek paszowych dla kurcząt rzeźnych - starter (%).
Składnik
Grupa
1
2
3
4
56,47
54,20
-
-
-
-
56,52
54,25
36,9
-
36,85
-
-
39,0
-
38,95
Olej rzepakowy
2,50
2,70
2,50
2,70
Kreda paszowa
1,70
1,70
1,70
1,70
Fosforan 1-wapniowy
1,40
1,37
1,40
1,37
NaCl
0,30
0,30
0,30
0,30
DL-Met
0,23
0,23
0,23
0,23
Premiks witaminowo-mineralny (0,5%)
0,50
0,50
0,50
0,50
220
220
220
220
Śruta kukurydziana konwencjonalna
Śruta kukurydziana GM
Poekstrakcyjna śruta sojowa konwencjonalna
Poekstrakcyjna śruta sojowa GM
Zawartość składników pokarmowych:
Białko ogólne (g/kg)
4
Energia metaboliczna (MJ/kg)
12,5
12,5
12,5
12,5
Lys (g/kg)
12,0
12,0
12,0
12,0
Met (g/kg)
5,50
5,50
5,50
5,50
Ca (g/kg)
9,40
9,40
9,40
9,40
P przyswajalny (g/kg)
4,30
4,30
4,30
,30
Tabela 3. Skład doświadczalnych mieszanek paszowych dla kurcząt rzeźnych – grower-finiszer (%).
Grupa
Składnik
1
2
3
4
60,25
58,54
-
-
-
-
60,30
58,64
32,0
-
31,95
-
-
33,45
-
33,40
Olej rzepakowy
3,60
3,90
3,60
3,85
Kreda paszowa
1,75
1,75
1,55
1,75
Fosforan 1-wapniowy
1,30
1,26
1,30
1,26
NaCl
0,30
0,30
0,30
0,30
DL-Met
0,21
0,21
0,21
0,21
Chlorowodorek L-Lys
0,09
0,09
0,09
0,09
Premiks witaminowo-mineralny (0,5%)
0,50
0,50
0,50
0,50
Białko ogólne (g/kg)
200
200
200
200
Energia metaboliczna (MJ/kg)
13,0
13,0
13,0
13,0
Lys (g/kg)
11,5
11,5
11,5
11,5
Met (g/ g)
5,2
5,2
5,2
5,2
Ca (g/kg)
9,2
9,2
9,2
9,2
P przyswajalny (g/kg)
4,0
4,0
4,0
4,0
Śruta kukurydziana konwencjonalna
Śruta kukurydziana GM
Poekstrakcyjna śruta sojowa konwencjonalna
Poekstrakcyjna śruta sojowa GM
Zawartość składników pokarmowych:
Schemat doświadczenia był następujący:
Grupa 1 - dieta zawierająca śrutę kukurydzianą i poekstrakcyjną śrutę sojową linii
konwencjonalnych,
Grupa 2 - śruta kukurydziana linii konwencjonalnej i poekstrakcyjna śruta sojowa GM,
Grupa 3 - śruta kukurydziana GM i poekstrakcyjna śruta sojowa linii konwencjonalnej,
Grupa 4 - śruta kukurydziana i poekstrakcyjna śruta sojowa pochodzące z roślin GM.
5
W oparciu o zebrane dane obliczono, w poszczególnych okresach odchowu, podstawowe
wskaźniki produkcyjne, tj. przyrost masy ciała, stopień wykorzystania paszy, procent padnięć
oraz europejski wskaźnik odchowu. Po zakończeniu doświadczenia, z każdej grupy wybrano po
24 sztuki kurcząt (12 kurek i 12 kogutków), które po uboju poddano analizie dysekcyjnej. Przy
uwzględnieniu masy żywej kurczęcia, masy poubojowej, masy mięśni piersiowych oraz masy
tłuszczu sadełkowego, obliczono podstawowe wskaźniki poubojowe, tj. wydajność rzeźną oraz
procentowy udział mięśni piersiowych i tłuszczu sadełkowego w tuszce.
Pobrane zostały próbki wybranych tkanek i narządów (krew, mięsień piersiowy,
wątroba, nerki, śledziona, trzustka, torba Fabrycjusza, płuca) oraz poszczególnych odcinków
przewodu pokarmowego wraz z treścią (wole, żołądek mięśniowy, dwunastnica, jelito czcze,
jelito
biodrowe,
jelita
ślepe,
stek)
w
celu
wykonania
specjalistycznych
analiz,
charakteryzujących status zdrowotny ptaków. Analizy te były wykonywane w Państwowym
Instytucie Weterynaryjnym - PIB w Puławach i obejmowały:
-analizy immunologiczne krwi przy zastosowaniu techniki cytometrii przepływowej
(immunofenotypowanie limfocytów krwi),
-ocenę stanu odpowiedzi immunologicznej po wykonanych szczepieniach profilaktycznych
(rzekomy pomór drobiu, choroba Gumboro, IB),
-ocenę makroskopową zmian anatomopatologicznych wybranych tkanek i narządów,
-badania histopatologiczne i ocenę zmian morfologicznych w wybranych tkankach i narządach,
-określenie obecności transgenicznego DNA, pochodzącego ze śruty sojowej GM i śruty
kukurydzianej GM, w wybranych narządach i tkankach kurcząt.
Wyniki
Badania prowadzone w Instytucie Zootechniki PIB
Wskaźniki produkcyjne kurcząt
W doświadczeniu na kurczętach rzeźnych, we wszystkich grupach doświadczalnych,
uzyskano bardzo dobre wskaźniki produkcyjne. Po starterowym okresie odchowu (21 dzień
życia) średnia masa ciała ptaków dla całego doświadczenia wynosiła 790 g, pobranie paszy –
1150 g/szt., współczynnik wykorzystania paszy - 1,54 kg/kg przyrostu a odsetek sztuk padłych
– 1,65%. W 42 dniu życia średnia masa ciała wynosiła 2460 g, pobranie paszy – 4400 g/szt.,
współczynnik wykorzystania paszy - 1,82 kg/kg przyrostu, odsetek sztuk padłych – 3,70% i
wskaźnik efektywności odchowu (WEO), który w sposób syntetyczny charakteryzuje przebieg
6
tuczu kurcząt – 310 punktów. Analizując statystycznie otrzymane wyniki produkcyjne nie
stwierdzono różnic pomiędzy grupami doświadczalnymi, żywionymi bez udziału lub z
udziałem badanych materiałów paszowych GM (Tabela 4, 5, 6).
Tabela 4.
Wyniki produkcyjne w starterowym okresie odchowu (1-21 dzień życia).
1
Grupa doświadczalna*
2
3
Poziom
P
SEM
4
Przyrost masy ciała (g)
747
741
751
745
0,900
4,502
Pobranie paszy (g)
1146
1143
1157
1155
0,856
6,09
Współczynnik
wykorzystania paszy
(kg/kg przyrostu)
1,534
1,545
1,542
1,550
0,956
0,0094
Padnięcia (%)
2,38
0,60
2,38
1,78
0,390
0,406
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 5. Wyniki produkcyjne w growerowo-finiszerowym okresie odchowu (22-42 dzień życia).
1
Grupa doświadczalna*
2
3
Poziom
P
SEM
4
Przyrost masy ciała (g)
1667
1681
1666
1685
0,851
8,887
Pobranie paszy (g)
Współczynnik
wykorzystania paszy
(kg/kg przyrostu)
3235
3235
3261
3275
0,872
18,94
1,940
1,926
1,958
1,943
0,884
0,0133
1,78
1,98
1,19
1,78
0,467
0,3898
Padnięcia (%)
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 6.
Wyniki produkcyjne w całym okresie odchowu (1-42 dzień życia).
1
Grupa doświadczalna*
2
3
SEM
4
Poziom
P
Przyrost masy ciała (g)
2414
2422
2417
2430
0,974
11,592
Pobranie paszy (g)
4381
4378
4418
4430
0,845
23,043
Współczynnik
wykorzystanie paszy
(kg/kg przyrostu)
1,815
1,808
1,829
1,821
0,934
0,0110
Padnięcia (%)
4,16
2,58
3,57
3,57
0,894
0,3048
WEO (punkty)
309
313
309
312
0,941
2,7604
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
7
Wyniki analizy rzeźnej i skład chemiczny mięśni
We wszystkich grupach doświadczalnych odnotowano bardzo podobne wskaźniki
poubojowe, tj. wydajność rzeźną, zawartość mięśni piersiowych i tłuszczu sadełkowego w
tuszce oraz względną masę wybranych narządów wewnętrznych (Tabela 7), nie wykazując
oddziaływania czynnika doświadczalnego na wymienione parametry (P>0,05). Badane
transgeniczne materiały paszowe nie miały również wpływu na skład chemiczny mięśni
piersiowych kurcząt (Tabela 8).
Podsumowując wyniki badań produkcyjnych wykonanych na kurczętach rzeźnych
należy stwierdzić, że nie wykazano wpływu ocenianych materiałów paszowych GM na
podstawowe wskaźniki odchowu, wyniki analizy rzeźnej i skład chemiczny mięśni
piersiowych.
Tabela 7.
Wyniki analizy rzeźnej.
1
Wydajność rzeźna (%)
Udział mięśnia
piersiowego w tuszce
(%)
Udział tłuszczu
sadełkowego w tuszce
(g)
Względna masa
wątroby (% masy
żywej)
Względna masa żołądka
mięśniowego (% masy
żywej)
Względna masa
śledziony (% masy
żywej)
Grupa doświadczalna*
2
3
Poziom
P
SEM
4
77,09
77,87
77,66
77,82
0,1833
0,141
26,47
26,36
26,09
66,82
0,4168
0,157
1,827
1,948
1,978
1,942
0,6979
0,047
2,047
1,902
2,033
1,935
0,4041
0,035
1,177
1,174
1,194
1,166
0,9651
0,019
0,1636
0,1519
0,1496
0,1433
0,6636
0,005
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 8.
Skład chemiczny mięśni piersiowych kurcząt (%).
1
Zawartość suchej masy
Zawartość białka
ogólnego
Zawartość tłuszczu
surowego
Grupa doświadczalna*
2
3
SEM
4
Poziom
P
25,5
25,3
25,5
25,4
0,888
0,087
23,5
23,5
23,7
23,8
0,626
0,071
1,08
1,01
1,04
1,02
0,643
0,020
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
8
Badania prowadzone w Instytucie Weterynaryjnym - PIB
Ocena humoralnej odpowiedzi immunologicznej
W ramach podjętych badań wykonano ocenę stanu odpowiedzi immunologicznej po
szczepieniach profilaktycznych kurcząt przeciwko rzekomemu pomorowi drobiu (ND),
zakaźnemu zapaleniu oskrzeli (IB) i chorobie Gumboro (IBD). W 1 dniu życia kurcząt od 20
sztuk pochodzących z tego samego wylęgu pobrano próbki krwi do badań serologicznych dla
określenia poziomu przeciwciał matczynych i terminów szczepień dla ND i IBD. Szczepienie
kurcząt doświadczalnych przeprowadzono w: 1 dniu życia przeciwko IB (w wylęgarni), 14 dniu
życia przeciwko IBD i 21 dniu życia przeciwko ND i IB przy użyciu żywych szczepionek
komercyjnych. Na zakończenie doświadczenia w 42 dniu życia kurcząt pobrano losowo próbki
krwi od 20 sztuk z każdej grupy doświadczalnej do badań serologicznych w kierunku ND, IB i
IBD. Uzyskaną z krwi surowicę badano metodą ELISA stosując komercyjne zestawy.
Wszystkie kurczęta rzeźne w 1 dniu życia posiadały przeciwciała matczyne przeciwko
rzekomemu pomorowi drobiu (ND), zakaźnemu zapaleniu oskrzeli (IB) i chorobie Gumboro
(IBD), wyrażone 100% serokonwersją i wysokim poziomem miana średniego, jednak o dużym
zróżnicowaniu mian indywidualnych (Tabela 9). Miało to niewątpliwie niekorzystny wpływ na
wyniki przeprowadzonych szczepień czynnych w trakcie doświadczenia. Zastosowany w
doświadczeniu, powszechny w warunkach fermowych, sposób podania szczepionki (w wodzie
pitnej) przeciwko IBD (14 dzień życia) oraz ND i IB (21 dzień życia) mógł także wpłynąć na
końcowe wyniki badań. Za powyższym przemawiają przede wszystkim wyniki końcowe
odpowiedzi immunologicznej po szczepieniu przeciwko chorobie Gumboro, która to
odpowiedź charakteryzowała się niskim odsetkiem serokonwersji, jak też niskim poziomem
średniego miana przeciwciał - przy jednocześnie dużym zróżnicowaniu mian indywidualnych.
Porównując średnie miana przeciwciał w grupie 1 (kontrolnej) i grupach 2-4 znamienną różnicę
wykazano tylko w odniesieniu do grupy 4/IB. Należy zaznaczyć, że wszystkie grupy
żywieniowe, zarówno kontrolna jak i doświadczalne szczepione były według takiej samej
procedury, tak więc czynnik ten nie różnicował uzyskiwanych w poszczególnych grupach
wyników odpowiedzi immunologicznej. Uogólniając, można przyjąć, że stosowane
materiały paszowe GM nie miały negatywnego wpływu na status immunologiczny kurczat
po wykonanych szczepieniach profilaktycznych w kierunku rzekomego pomoru drobiu
(ND), zakaźnego zapalenia oskrzeli (IB) i choroby Gumboro (IBD).
9
Tabela 9.
Stan humoralnej odpowiedzi immunologicznej u kurcząt rzeźnych.
Grupa doświadczalna*
1
2
3
4
ND – termin 0 (1 dzień życia)
Serokonwersja (%)
20/20 (100)
20/20 (100)
20/20 (100
20/20 (100)
Średnie miano
4611,4
4611,4
4611,4
4611,4
Zakres zmian
1713-7235
1713-7235
1713-7235
1713-7235
ND – termin 1 (42 dzień życia)
Serokonwersja (%)
15/20 (75)
13/20 (65)
13/20
10/20 (50)
Średnie miano
1222,8
1122,6
628,6
760,2
Zakres zmian
111-5895
1-8248
15-1772
3-3605
IB – termin 0 (1 dzień życia)
Serokonwersja (%)
20/20 (100)
20/20 (100)
20/20 (100)
20/20 (100)
Średnie miano
6917,5
6917,5
6917,5
6917,5
Zakres zmian
680-7808
680-7808
680-7808
680-7808
IB – termin 1 (42 dzień życia)
Serokonwersja (%)
15/20 (75)
13/20 (65)
8/20
9/20 (45)
Średnie miano
872,0
1156,1
622,3
464,2
Zakres zmian
74-2159
7-4704
219-2376
61-1034
IBD – termin 0 (1 dzień życia)
Serokonwersja (%)
20/20 (100)
20/20 (100)
20/20 (100)
20/20 (100)
Średnie miano
5117,9
5117,9
5117,9
5117,9
Zakres zmian
2739-8297
2739-8297
2739-8297
2739-8297
IBD – termin 1 (42 dzień życia)
Serokonwersja (%)
5/20 (25)
1/20 (5)
3/20 (15)
1/20 (5)
Średnie miano
293,5
107,1
704,1
231,7
Zakres zmian
32-1971
1-969
4-7670
4-605
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Ocena komórkowej odpowiedzi immunologicznej
Przeprowadzono także ocenę wpływu stosowanych pasz GM na wskaźniki nieswoistej
odporności komórkowej u kurcząt. Oceniano skład procentowy poszczególnych subpopulacji
limfocytów krwi obwodowej badanych ptaków z użyciem cytometrii przepływowej i
specyficznego panelu przeciwciał monoklonalnych umożliwiających rozpoznawanie komórek
CD3+ (limf. T), CD4+ (limf. Th; pomocnicze) i CD8a+ (limf. Tc/s; cytotoksyczno-supresorowe).
Zestawienie uzyskanych danych doświadczalnych wskazuje na brak różnic we wskaźnikach
10
odporności komórkowej związanej z kształtowaniem się subpopulacji obwodowych limfocytów
krwi kurcząt rzeźnych, żywionych dietami zawierającymi GM w porównaniu ze zwierzętami
żywionymi dietą konwencjonalną (Tabela 10).
Tabela 10.
Zmiany subpopulacjach limfocytów krwi obwodowej u kurcząt rzeźnych.
Okres
Wskaźnik
+
I („0”)
II
Gr. 1
20,4
±0,7
19,3
±2,3
CD3
Gr. 2
21,1
±2,4
27,3
±4,4
(%)
Gr. 3
21,3
±2,3
29,5
±2,9
Gr. 4
19,8
±0,5
22,4
±3,9
Gr. 1
10,9
±2,2
11,1
±2,9
CD4+(%)
Gr. 2 Gr. 3
13,7
11,4
±1,7 ±1,7
17,8
18,6
±0,6 ±0,7
Gr. 4
10,6
±1,3
14,7
±2,8
Gr. 1
7,5
±0,8
7,6
±1,1
CD8a+(%)
Gr. 2 Gr. 3
10,7
11,0
±2,0
±2,2
8,7
9,4
±1,2
±0,5
Gr. 4
8,4
±1,0
8,0
±0,9
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Przedstawione wyniki analiz krwi kurcząt w doświadczeniu wykonano w celu określenia
czy DNA badanych pasz transgenicznych nie wpływa na procesy immunologiczne, nie
wywołując nadmiernej alergii, czy też procesów zapalnych (zmian w obrazie białokrwinkowym
i subpopulacjach limfocytów) oraz nie zakłóca odpowiedzi humoralnej organizmu na
zastosowane szczepienia profilaktyczne. Badania te są szczególnie ważne u nowoczesnych
krzyżówek kurcząt rzeźnych, gdyż długoletnia selekcja w kierunku szybkiego tempa przyrostu
masy ciała miała ujemny wpływ na swoiste mechanizmy odporności, a głównie na zdolność do
syntezy przeciwciał, natomiast zwiększyła natężenie niekorzystnych reakcji zapalnych.
Przedstawione wyniki nie wykazały negatywnego wpływu badanych materiałów
paszowych GM na działanie układu odpornościowego kurcząt, zarówno w zakresie
odporności humoralnej, jak i komórkowej.
Wyniki badań histopatologicznych i ocena przyżyciowa statusu zdrowotnego
Wyniki badań histopatologicznych przedstawiono w Tabeli 11. Do badań pobierano
wycinki wątroby, nerek, śledziony, trzustki, dwunastnicy, jelita czczego oraz torby Fabrycjusza
(od 10-ciu ptaków z każdej grupy, w wieku 42 dni). Z utrwalonego w 10% roztworze
zbuforowanej formaliny materiału wykonywano skrawki parafinowe, które następnie barwiono
metodą hematoksylina-eozyna (H-E). Analiza otrzymanych wyników nie wykazała różnic
pomiędzy poszczególnymi grupą kontrolną (dieta bez materiałów paszowych GM) a grupami
doświadczalnymi (diety z udziałem materiałów paszowych GM). W przypadku wątroby i nerek
zmiany histopatologiczne stwierdzono we wszystkich próbkach, natomiast w przypadku
śledziony – w części analizowanych próbek (Tabela 11). Stwierdzone zmiany histopatologiczne
występowały w podobnym zakresie we wszystkich grupach, zarówno w grupie kontrolnej, jak i
11
grupach doświadczalnych, niezależnie od udziału w diecie materiałów paszowych GM.
Stwierdzane zmiany stanowią prawdopodobnie efekt intensywnego żywienia, w tym dużej
zawartości białka i energii metabolicznej w dietach, oraz wysokiego tempa metabolizmu u
szybko rosnących kurcząt rzeźnych, natomiast piankowata struktura hepatocytów była
prawdopodobnie artefaktem charakterystycznym dla wątroby, z której wyługowany został
zhydrolizowany w czasie obróbki histologicznej glikogen. Podobnie ocena przyżyciowa statusu
zdrowotnego zwierząt oraz badanie anatomo-patologiczne (obserwacja makroskopowa) tkanek
i narządów wewnętrznych nie wykazały różnic między grupą kontrolną, a grupami
doświadczalnymi żywionymi z udziałem pasz genetycznie zmodyfikowanych. Uzyskane
wyniki wskazują zatem, że badane materiały paszowe GM nie mają wpływu na obraz
histopatologiczny narządów wewnętrznych kurcząt oraz oceniany przyżyciowo status
zdrowotny kurcząt.
Tabela 11.
Wyniki badań histopatologicznych narządów wewnętrznych kurcząt.
Grupa doświadczalna*
Wątroba
Nerki
Śledziona
1
2
3
4
We wszystkich
przypadkach (10/10),
obecność nacieków
komórek limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych) od
nieznacznego do
umiarkowanego stopnia
oraz piankowatej
struktury hepatocytów od
nieznacznego do silnego
stopnia.
We wszystkich
przypadkach (10/10),
przekrwienie miąższu od
nieznacznego do
umiarkowanego stopnia.
W większości
przypadków: ogniskowe
nacieki komórek
limfoidalnych (8/10)
i/lub miejscowa
martwica jąder (rozpad
i/lub zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(7/10).
W niektórych
przypadkach:
przekrwienie miąższu
(4/10) i/lub zwiększona
liczba ośrodków
rozmnażania (4/10).
We wszystkich
przypadkach (10/10),
obecność nacieków
komórek limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych) od
nieznacznego do
umiarkowanego stopnia
oraz piankowatej
struktury hepatocytów od
nieznacznego do silnego
stopnia.
We wszystkich
przypadkach (10/10),
przekrwienie miąższu od
nieznacznego do
umiarkowanego stopnia.
W większości
przypadków: ogniskowe
nacieki komórek
limfoidalnych (8/10)
i/lub miejscowa
martwica jąder (rozpad
i/lub zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(7/10).
W niektórych
przypadkach:
przekrwienie miąższu
(4/10) i/lub zwiększona
liczba ośrodków
rozmnażania (3/10).
We wszystkich
przypadkach (10/10),
obecność nacieków
komórek limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych) od
nieznacznego do
umiarkowanego stopnia
oraz piankowatej
struktury hepatocytów od
nieznacznego do silnego
stopnia.
We wszystkich
przypadkach (10/10),
przekrwienie miąższu od
nieznacznego do
umiarkowanego stopnia.
W większości
przypadków: ogniskowe
nacieki komórek
limfoidalnych (7/10)
i/lub miejscowa
martwica jąder (rozpad
i/lub zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(9/10).
W niektórych
przypadkach:
przekrwienie miąższu
(5/10) i/lub zwiększona
liczba ośrodków
rozmnażania (3/10).
We wszystkich
przypadkach (10/10),
obecność nacieków
komórek limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych) od
nieznacznego do
umiarkowanego stopnia
oraz piankowatej
struktury hepatocytów od
nieznacznego do silnego
stopnia.
We wszystkich
przypadkach (10/10),
przekrwienie miąższu od
nieznacznego do
umiarkowanego stopnia.
W większości
przypadków: ogniskowe
nacieki komórek
limfoidalnych (8/10)
i/lub miejscowa
martwica jąder (rozpad
i/lub zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
9/10).
W niektórych
przypadkach:
przekrwienie miąższu
(4/10) i/lub zwiększona
liczba ośrodków
rozmnażania (3/10).
12
Trzustka
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego narządu.
Dwunastnica
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego narządu.
Jelito czcze
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego n rządu.
Torba Fabrycjusza
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego narządu.
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Analiza obecności transgenicznego DNA w narządach, tkankach i przewodzie pokarmowym
kurcząt rzeźnych
Analizy obecności transgenicznego DNA, pochodzącego ze śruty sojowej i
kukurydzianej GM, w wybranych narządach i tkankach kurcząt oraz w poszczególnych
odcinkach przewodu pokarmowego wykonano w Krajowym Laboratorium Pasz Instytutu
Zootecniki PIB (pracownia w Szczecinie, posiadającym akredytację w zakresie oznaczeń
transgenicznego DNA soi i kukurydzy. Analizowane były następujące próbki materiału
biologicznego:
-krew,
-mięsień piersiowy,
-wątroba,
-płuca,
-śledziona,
-oraz treść wola, żołądka, dwunastnicy, jelita czczego, jelita biodrowego, jelit ślepych i jelita
końcowego ze stekiem.
W grupach żywionych mieszankami paszowymi zawierającymi śrutę z ziarna kukurydzy
GM, lub/i poekstrakcyjną śrutę sojową GM, nie stwierdzano obecności w badanych tkankach i
narządach specyficznego dla danej modyfikacji (transgenicznego) DNA (Tabela 12). W
przypadku przewodu pokarmowego transgeniczny DNA występował jedynie w przedniej części
przewodu pokarmowego, tj. w treści wola i żołądka.
Wyniki uzyskane w Krajowym Laboratorium Pasz Instytutu Zootecniki PIB zostały
potwierdzone w laboratorium Państwowego Instytutu Weterynaryjnego - PIB w Puławach, w
którym, w próbkach materiału biologicznego z doświadczenia, badano obecność genów
referencyjnych
soi
(lektyna)
i
kukurydzy
(inwertaza)
oraz
obecność
sekwencji
charakterystycznych dla stosowanych modyfikacji, tj. promotora 35S i terminatora NOS. W
DNA wyizolowanym z krwi, narządów i tkanek (wątroba, śledziona i nerka) oraz w kale
ptaków żywionych mieszankami paszowymi zawierającymi materiały GM nie stwierdzono
obecności fragmentów DNA charakterystycznych dla zmodyfikowanej soi i kukurydzy
(promotora 35S i terminatora NOS). Wyniki badania treści wola były w pełni skorelowane z
13
rodzajem stosowanej mieszanki doświadczalnej. Podsumowując można stwierdzić, że brak
wykrywalnych fragmentów transgenów już od pierwszego fragmentu jelita cienkiego
(dwunastnicy) wskazuje na fakt, że kwasy nukleinowe, również transgeniczne DNA, są u
drobiu efektywnie denaturowane w niskim odczynie pH treści żołądka, a następnie
trawione przez enzymy (nukleazy) trzustkowe i jelitowe. Wyklucza to praktycznie
możliwość transportu czynnych fragmentów transgenicznego DNA przez barierę jelitową
do narządów i tkanek, jak również ich pasaż w formie niestrawionej przez jelita i
wydalanie wraz kałem do środowiska.
Tabela 12.
Wyniki jakościowej analizy DNA w kierunku obecności specyficznych dla GMO
transgenów w tkankach i narządach kurcząt.
Grupa doświadczalna*
Materiał biologiczny
RR
1
MON810
RR
2
MON810
RR
Treść wol
-
-
+
Treść żołądka
Treść dwunastnicy
Treść jelita biodrowego
Treść jelita czczego
Treść jelita ślepych
Treść jelita końcoweg i
steku
Krew
Wątroba
Płuca
Śledziona
Mięsień piersiowy
-
-
-
-
3
MON810
RR
4
MON810
+
-
n/a**
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
+
+
+
+
-
+
-
+
-
-
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
n/a
-
-
-
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
**/ nie analizowano
14
BADANIA NA KURACH NIEŚNYCH
Doświadczenie żywieniowe na kurach nieśnych krzyżówki komercyjnej Bovans Brown
wykonano w kurniku doświadczalnym Zakładu Doświadczalnego IZ PIB Rudawa Sp. z o.o.
Okres doświadczalny obejmował 29 tygodni, tj. okres od 25 do 54 tygodnia życia ptaków.
Nioski utrzymywano w pojedynczych klatkach o wymiarach 40 x 35 cm, na podłodze z siatki,
przy stałym dostępie do paszy i wody.
W doświadczeniu stosowano izobiałkowe i izoenergetyczne mieszanki paszowe, w skład
których wchodziła śruta kukurydziana, poekstrakcyjna śruta sojowa oraz dodatki mineralne,
aminokwasy krystaliczne, olej rzepakowy i premiks witaminowo-mineralny (Tabela 13).
Zawartość składników pokarmowych i energii metabolicznej w mieszankach pokrywała
zapotrzebowanie wysoko produkcyjnych kur nieśnych.
Tabela 13.
Skład doświadczalnych mieszanek paszowych dla kur nieśnych (%).
Składnik
Grupa
1
2
3
4
Śruta kukurydziana konwencjonalna
Śruta kukurydziana GM
61,75
-
60,20
-
61,80
60,15
Poekstrakcyjna śruta sojowa konwencjonalna
25,70
-
25,65
-
-
27,00
-
27,05
Olej rzepakowy
1,00
1,25
1,00
1,25
Kreda paszowa
9,40
9,40
9,40
9,40
Fosforan 1-wapniowy
1,25
1,25
1,25
1,25
NaCl
0,30
0,30
0,30
0,30
DL-Met
0,10
0,10
0,10
0,10
Premiks witaminowo-mineralny (0,5%)
0,50
0,50
0,50
0,50
Białko ogólne (g/kg)
170
170
170
170
Energia metaboliczna (MJ/kg)
11,6
11,6
11,6
11,6
Lys (g/kg)
Met (g/kg)
Ca (g/kg)
P przyswajalny (g/kg)
8,80
3,50
37,0
3,75
8,80
3,50
37,0
3,75
8,80
3,50
37,0
3,75
8,80
3,50
37,0
3,7
Poekstrakcyjna śruta sojowa GM
Zawartość skład ików pokarmowych:
Utworzono 4 grupy doświadczalne, składające się z 24 niosek utrzymywanych w
indywidualnych klatkach. Czynnikiem doświadczalnym była obecność w mieszance paszowej
materiałów paszowych pochodzących z roślin genetycznie zmodyfikowanych.
15
Układ grup doświadczalnych był identyczny jak w przypadku kurcząt rzeźnych:
Grupa 1 - dieta zawierająca śrutę kukurydzianą i poekstrakcyjną śrutę sojową linii
konwencjonalnych,
Grupa 2 - śruta kukurydziana linii konwencjonalnej i poekstrakcyjna śruta sojowa GM,
Grupa 3 - śruta kukurydziana GM i poekstrakcyjna śruta sojowa linii konwencjonalnej,
Grupa 4 - śruta kukurydziana i poekstrakcyjna śruta sojowa z roślin GM.
W trakcie doświadczenia notowano liczbę i masę zniesionych jaj oraz ilość pobranej
paszy. Na tej podstawie obliczono wydajność nieśną, dzienne pobranie paszy oraz zużycie
paszy w przeliczeniu na 1 kg jaj i na 1 jajo. Wykonano również badania jakości jaj, z
uwzględnieniem takich cech jak barwa i masa żółtka, jednostki Haugha, masa, grubość, gęstość
i wytrzymałość skorupy oraz ilość plam krwawych i mięsnych. Jaja pobierano również w celu
analizy ewentualnej obecności transgenicznego DNA w ich treści.
Po zakończeniu doświadczenia żywieniowego wykonano ubój doświadczalny niosek w
celu pobrania wybranych tkanek, narządów i treści przewodu pokarmowego do oznaczeń
zawartości transgenicznego DNA oraz wykonania specjalistycznych analiz parametrów
charakteryzujących status zdrowotny ptaków. Pobrano próbki następujących narządów i
tkanek: krew, wątroba, nerki, śledziona, trzustka, torba Fabrycjusza, płuca, mięśnie piersiowe,
poszczególne odcinki przewodu pokarmowego wraz z treścią (wole, żołądek mięsniowy,
dwunastnica, jelito czcze, jelito biodrowe, jelita ślepe, stek i kloaka).
Wyniki
Badania wykonane w Instytucie Zootechniki PIB
Wskaźniki wydajności nieśnej i jakość jaj
We wszystkich grupach, tj. w grupie kontrolnej i doświadczalnych, uzyskano podobną
wartość wskaźników charakteryzujących produkcyjność kur i jakość jaj. Analiza statystyczna
przedstawionych wyników nie wykazała wpływu badanych materiałów paszowych GM na
wskaźniki produkcyjne, tj. wydajność nieśną, pobranie i wykorzystanie paszy w
okresie
od 25 do 54 tygodnia życia kur (Tabela 14). Nie stwierdzono także takiego wpływu w zakresie
parametrów charakteryzujących jakość jaj, badanych w kolejnych okresach doświadczenia, tj.
w 36 i 48 tygodniu życia (Tabela 15, 16, 17, 18). Podsumowując uzyskane wyniki można
stwierdzić, że poekstrakcyjna śruta sojowa GM i śrucie kukurydzianej GM, stosowane w
badaniach, nie wpływają na wskaźniki produkcyjne kur i jakość jaj.
16
Tabela 14.
Produkcyjność nieśna za cały okres doświadczenia (25-54 tydzień życia niosek).
Grupa doświadczalna*
2
3
1
Nieśność (%)
Dzienna produkcja jaj (g/szt.)
Średnia masa 1 jaja (g)
Dzienne pobranie paszy (g/szt.)
Wykorzystanie paszy (g/1 jajo)
Wykorzystanie aszy (g/1 kg
jaj)
95,6
60,7
63,4
125
130
2,05
94,0
60,0
63,8
123
130
2,05
95,6
61,5
64,3
125
131
2,03
SEM
4
Poziom
P
95,5
61,2
64,1
125
131
2,05
0,283
0,517
0,798
0,111
0,897
0,950
0,344
0,365
0,330
0,377
0,464
0,010
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 15.
Wyniki analizy jakości treści jaj (36 tydzień życia).
Grupa doświadczalna*
2
3
1
Poziom
P
SEM
4
Wysokość białka (mm)
10,9
10,7
11,0
10,9
0,705
0,084
Jednostki Haugha
102,4
102,1
103,0
102,5
0,81
0,327
Masa żółtka (g)
15,6
15,8
15,7
15,9
0,798
0,128
Barwa żółtka (punkty w
skali Roche’a)
Indeks kształtu
4,37
4,62
4,37
4,58
0,559
0,080
78,4
79,0
79,8
79,7
0,180
0,256
Plamy krwawe
0,021
0,017
0,025
0,025
0,924
0,004
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 16.
Wyniki analizy jakości skorupy jaj (36 tydzień życia).
1
Względna masy skorupy
(%)
Grubość skorupy (μm)
Grupa doświadczalna*
2
3
Poziom P
SEM
4
11,1
11,2
11,2
11,1
0,929
0,060
393,8
395,5
392,5
386,9
0,714
2,723
Gęstość skorupy (mg/cm )
88,1
90,5
90,3
88,8
0,608
0,766
Wytrzymałość skorupy (N)
35,1
35,1
34,5
34,5
0,979
0,708
2
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
17
Tabela 17.
Wyniki analizy jakości treści jaj (48 tydzień życia).
Grupa doświadczalna*
2
3
1
Poziom P
SEM
4
Wysokość białka (mm)
8,34
8,22
8,40
8,01
0,661
0,117
Jednostki Haugha
88,9
89,7
90,3
88,5
0,828
0 688
Masa żółtka (g)
17,2
17,2
17,1
17,2
0,999
0,115
Barwa żółtka (punkty w
skali Roche’a)
Indeks kształtu
3,33
3,50
3,67
3,42
0,606
0,089
77,3
77,7
78,4
76,7
0,460
0,37
Plamy krwawe
0,025
0,017
0,008
0,025
0,775
0,029
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 18.
Wyniki analizy jakości skorupy jaj (48 tydzień życia).
1
Grupa doświadczalna*
2
3
Poziom P
SEM
4
Względna masy skorupy
(%)
Grubość skorupy (μm)
11,4
11,4
11,2
11,2
0,831
0,106
402,4
411,2
402,9
407,5
0,911
4,83
Gęstość skorupy (mg/cm2)
91,4
92,3
91,1
91,4
0,968
0,904
Wytrzymałość skorupy
N)
32,3
34,2
35,4
32,6
0,834
0,32
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Badania strawnościowe
W trakcie badań przeprowadzono u niosek test bilansowo-strawnościowy. Uzyskane
wyniki nie wykazały różnic pomiędzy grupami doświadczalnymi. Badane materiały paszowe
GM nie miały wpływu na strawność pozorną składników pokarmowych, tj. suchej masy, masy
organicznej, białka ogólnego, tłuszczu surowego, substancji bezazotowych wyciągowych,
włókna surowego i popiołu surowego (Tabela 19). Również stopień wykorzystania energii
zawartej w mieszankach paszowych był we wszystkich grupach podobny. Nie odnotowano
także oddziaływania śruty z ziarna kukurydzy Bt i poekstrakcyjnej śruty sojowej RR na wyniki
bilansu azotu, wapnia i fosforu u niosek (Tabela 20). Podsumowując, można stwierdzić, że
powyższe wyniki, razem z rezultatami produkcyjnymi, wskazują na równowartość
żywieniową u kur nieśnych badanych materiałów paszowych GM i ich konwencjonalnych
18
odpowiedników.
Tabela 19.
Strawność pozorna składników pokarmowych (%) i współczynnik metaboliczności energii diety
(44 tydzień życia niosek).
Grupa doświadczalna*
Poziom P
SEM
1
2
3
4
Sucha masa
71,2
71,2
71,6
71,5
0,978
0,377
Masa organiczna
75,9
75,7
76,3
76,4
0,891
0,362
Białko ogólne
44,4
43,6
45,6
44,4
0,790
0,689
Tłuszcz surowy
80,6
79,8
80,5
81,1
0,5 6
0,284
Bezazotowe
wyciągowe
88,4
88,8
89,0
89,3
0,873
0,375
Włókno surowe
6,56
6,97
7,24
6,70
0,995
0,947
Popiół surowy
40,8
42,7
41,6
40,2
0,802
0,912
0,724
0,720
0,730
0,729
0,776
0,004
0,699
0,695
0,703
0,704
0,795
0,003
Współczynnik
metaboliczności
energii, AME
Współczynnik
metaboliczności
energii, AMEN
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 20.
Bilans azotu, wapnia i fosforu (44 tydzień życia).
Grupa doświadczalna*
1
2
3
Poziom P
SEM
4
N pobrany (mg/dz/szt.)
N wydalony (mg/dz/szt.)
N zatrzymany (mg/dz/szt.)
N zatrzymany (% N
pobranego)
Ca pobrany (mg/dz/szt.)
Ca wydalony (mg/dz/szt.)
2873
600
1273
44,4
2914
1644
1270
43,6
2894
1572
1322
45,6
2823
1569
1255
44,4
0,874
0,81
0,837
0,790
38,5
29,7
26,1
0,689
4065
2186
4185
2161
4017
2143
4004
2087
0,683
0,842
56,1
38,2
Ca zatrzymany (mg/dz/szt.)
1879
2024
1874
1917
0,746
51,9
Ca zatrzymany (% Ca
pobranego)
P pobrany (mg/dz/szt.)
46,4
48,0
46,4
47,9
0,889
0,913
668
681
668
648
0,671
9,17
P wydalony (mg/dz/szt.)
452
454
440
430
0,789
9,10
P zatrzymany (mg/dz/szt.)
216
227
229
218
0,953
9,20
P zatrzymany (% P
pobranego)
32,4
33,2
34,2
33,6
0,966
1,22
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
19
Badania prowadzone w Instytucie Weterynaryjnym - PIB
Ocena humoralnej odpowiedzi immunologicznej
W trakcie badań wykonano ocenę stanu odpowiedzi immunologicznej kur po
szczepieniach profilaktycznych przeciwko rzekomemu pomorowi drobiu (ND), zakaźnemu
zapaleniu oskrzeli (IB) i chorobie Gumboro (IBD). Ptaki zostały zaszczepione zgodnie z
zaleconym przez lekarza weterynarii programem immunoprofilaktycznym, w tym 3-krotnie
przeciwko ND i IB oraz 2-krotnie IBD szczepionkami żywymi i ponadto 1-krotnie przeciwko
ND i IB poliwalentną szczepionką inaktywowaną. Z każdej grupy doświadczalnej pobierano
próbki krwi do badań serologicznych w terminach: 2 tygodnie przed rozpoczęciem
doświadczenia (termin 0) od 26 sztuk, a w trakcie trwania doświadczenia (okres produkcji
nieśnej) 3-krotnie, w odstępach około 2 miesięcznych (terminy I, II i III), od 10 sztuk z każdej
grupy doświadczalnej. Do badania przeciwciał w surowicy użyto metody ELISA.
Badając serologicznie kury nioski przed rozpoczęciem doświadczenia (termin 0) można
stwierdzić, że w wyniku przeprowadzonych szczepień w okresie wychowu posiadały one dobry
status immunologiczny wyrażony wysokim odsetkiem serokonwersji i wysokim poziomem
średniego miana (odpowiednio: 28 144,8 dla ND, 8872,6 dla IB i 7110, 06 dla IBD).
Porównując oceniane parametry odpowiedzi immunologicznej po 6 miesięcznym okresie
trwania doświadczenia (termin III) nie wykazano różnic pomiędzy grupą 1 (kontrolną) a
pozostałymi grupami (grupy 2, 3 i 4), które otrzymywały w tym czasie diety z materiałami
paszowymi GM (Tabela 21). W grupach 2-4 odsetek serokonwersji wynosił od 90% do 100%
(przy 100% w grupie 1), a średnie miana przeciwciał były tylko nieznacznie niższe (grupa 2 i
4/IB oraz grupy 3 i 4/IBD) lub wyższe (grupy 2-4/ND, grupa 3/IB czy grupa 2/IBD) od
średnich mian w grupie 1. Podsumowując uzyskane wyniki badań immunologicznych,
można stwierdzić, że stosowanie materiałów paszowych GM nie miało negatywnego
wpływu na status immunologiczny kur nieśnych po wykonanych szczepieniach
profilaktycznych przeciwko rzekomemu pomorowi drobiu (ND), zakaźnemu zapaleniu
oskrzeli (IB) i chorobie Gumboro (IBD). Powyższy wniosek potwierdza wyniki analiz
immunologicznych otrzymane u kurcząt rzeźnych.
20
Tabela 21.
Stan odpowiedzi immunologicznej u niosek.
Grupa doświadczalna*
2
3
ND – termin 0
1
Serokonwersja (%)
Średnie miano
Zakres zmian
22/26 (84)
7355,2
1-10715
Serokonwersja (%)
Średnie miano
Zakres zmian
15/15 (100)
28144,8
1551-40927
Serokonwersja (%)
Średnie miano
Zakres zmian
10/10 (100)
12425,9
741-18185
Serokonwersja (%)
Średnie miano
Zakres zmian
10/10 (100)
11688,9
8632-15456
Serokonwersja (%)
Średnie miano
Zakres zmian
24/26 (92)
11627,2
95-12382
Serokonwersja (%)
Średnie miano
Zakres zmian
15/15 (100)
8872,6
732-11276
Serokonwersja (%)
Średnie miano
Zakres zmian
9/10 (90)
9731,3
334-13711
Serokonwersja (%)
Średnie miano
Zakres zmian
10/10 (100)
7809,5
2238-13165
Serokonwersja (%)
Średnie miano
Zakres zmian
26/26 (100)
5934,1
2630-9989
Serokonwersja (%)
Średnie miano
Zakres zmian
15/15 (100)
7110,1
1495-12055
Serokonwersja (%)
Średnie miano
Zakres zmian
10/10 (100)
7585,8
4845-11475
Serokonwersja (%)
Średnie miano
Zakres zmian
10/10 (100)
3757,9
858-7479
22/26 (85)
22/26 (85)
7355,2
7355,2
1-10715
1-10715
ND – termin 1
14/15 (93)
12/15 (80)
36275.9
35744,8
315-48789
1-48268
ND - termin 2
9/10 (90)
9/10 (90)
11939,6
12343,2
271-18841
180-19382
ND – termin 3
9/10 (90)
10/10 (100)
15220,8
15165,7
126-21795
3288-21386
IB – termin 0
24/26 (92)
24/26 (92)
11627,2
11627,2
95-12382
95-12382
IB – termin 1
14/15 (93)
15/15 (100)
5200,8
6234,5
70-10786
862-13228
IB – termin 2
9/10 (90)
10/10 (100)
5353,6
6110,4
118-11295
1241-12526
IB – termin 3
10/10 (100)
10/10 (100)
7540,5
8985,5
1599-12273
796-15030
IBD – termin 0
26/26 (100)
26/26 (100)
5934,1
5934,1
2630-9989
2630-9989
IBD – termin 1
15/15 (100)
15/15 (100)
5329,5
5399,5
2114-8531
2650-10180
IBD – termin 2
10/10 (100)
10/10 (100)
5775,4
5460,2
2564-8855
2822 8440
IBD – termin 3
10/10 (100)
10/10 (100)
4642,6
3266,4
3085-6878
2042-5601
4
22/26 (85)
7355,2
1-10715
15/15 (100)
36582,3
19478-51301
9/10 (90)
14417,8
392-20140
9/10 (90)
14972,3
311-21819
24/26 (92)
11627,2
95-12382
15/15 (100)
6472,6
1897-11193
9/10 (90)
6983,4
275-10557
10/10 (100)
6118,9
1182-11287
26/26 (100)
5934,1
2630-9989
15/15 (100)
5285,9
3014-8821
9/10 (90)
4838,5
116-7968
9/10 (90)
2907,7
1-6510
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
21
Ocena komórkowej odpowiedzi immunologicznej
Statystyczne zestawienie wyników analiz wykonanych przy użyciu metody cytometrii
przepływowej wskazuje na brak różnic (P>0,05) we wskaźnikach odporności komórkowej
związanej z kształtowaniem się subpopulacji obwodowych limfocytów krwi niosek żywionych
dietą kontrolną, nie zawierającą pasz transgenicznych, lub dietami doświadczalnymi (Tabela
22). Można zatem stwierdzić, że badane materiały paszowe GM nie wpływają u kur
nieśnych na analizowane parametry chcarakteryzujące odporność typu komórkowego.
Tabela 22.
Zmiany w subpopulacjach limfocytów krwi obwodowej u niosek.
Okres
Wskaźnik
CD3+(%)
CD4+(%)
CD8a+(%)
Gr. 1
Gr. 2
Gr. 3
Gr. 4
Gr. 1
Gr. 2
Gr. 3
Gr. 4
Gr. 1
Gr. 2
Gr. 3
Gr. 4
I („0”)
29,4
±2,0
24,4
±1,9
22,5
±2,8
21,6
±3,1
18,9
±2,9
14,7
±1,2
12,3
±1,5
13,5
±2,1
10,1
±0,46
11,5
±2,5
11,4
±0,8
9,9
±0,36
II
27,9
±2,4
29,6
±3,9
20,0
±5,1
28,8
±3,2
12,5
±2,8
19,5
±2,7
17,7
±2,1
15,5
±2,8
7,8
±2,7
9,9
±1,1
7,7
±1,5
8,6
±3,9
III
31,0
±6,1
30,9
±6,3
23,2
±4,3
24,7
±2,8
18,2
±2,6
18,3
±3,5
16,1
±2,9
14,7
±3,2
9,7
±2,8
11,2
±2,7
8,0
±0,6
8,5
±0,8
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Wyniki badań histopatologicznych i przyżyciowa ocena statusu zdrowotnego
Wyniki badań histopatologicznych przedstawiono w Tabeli 23. Do badania
histopatologicznego pobierano od 10 niosek z każdej grupy (łącznie 40 sztuk), ubitych po
zakończeniu doświadczenia produkcyjnego, wycinki wątroby, nerek, śledziony, trzustki,
dwunastnicy, jelita czczego oraz mięśni szkieletowych (mięśnie piersiowe). Z utrwalonego w
10% roztworze zbuforowanej formaliny materiału wykonywano skrawki parafinowe, które
następnie barwiono metodą hematoksylina-eozyna (H-E). Badanie histopatologiczne wątroby,
nerek, śledziony, trzustki, dwunastnicy, jelita czczego oraz mięśni szkieletowych kur niosek nie
wykazało znaczących różnic pomiędzy grupą kontrolną, żywionych dietą bez materiałów
paszowych GM, oraz poszczególnymi grupami doświadczalnymi (diety z udziałem materiałów
paszowych GM). W przypadku niektórych narządów i tkanek (wątroba, nerki, śledziona,
trzustka, mięśnie szkieletowe) obserwowano zmiany, które występowały w podobnym zakresie
liczbowym i miały podobny wygląd, zarówno w grupie kontrolnej, jak i w grupach
doświadczalnych, niezależnie od stosowania badanych materiałów paszowych GM (Tabela 23).
22
Obserwowane zmiany stanowią prawdopodobnie efekt intensywnego żywienia, w tym dużej
zawartości białka i energii metabolicznej w dietach, oraz szybkiego tempa metabolizmu u
wysokoprodukcyjnych kur nieśnych, natomiast piankowata struktura hepatocytów była
prawdopodobnie artefaktem charakterystycznym dla wątroby, z której wyługowany został
zhydrolizowany w czasie obróbki histologicznej glikogen. Ocena przyżyciowa statusu
zdrowotnego zwierząt oraz badanie anatomo-patologiczne (obserwacja makroskopowa) tkanek
i narządów wewnętrznych również nie wykazały różnic między grupą kontrolna, a grupami
doświadczalnymi żywionymi z udziałem poeksktrakcyjnej śruty sojowej GM oraz śruty z
ziarna kukurydzy GM. Uzyskane wyniki wskazują zatem, że badane materiały paszowe
GM nie mają wpływu na obraz histopatologiczny narządów wewnętrznych oraz oceniany
przyżyciowo status zdrowotny kur nieśnych.
Tabela 23.
Wyniki badań histopatologicznych narządów wewnętrznych niosek.
Grupa doświadczalna*
Wątroba
Nerki
Śledziona
Trzustka:
1
2
3
4
W niektórych
przypadkach:
przekrwienie miąższu
(5/10), nacieki komórek
limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych)
nieznacznego stopnia
(4/10), piankowa
struktura hepatocytów
nieznacznego stopnia
(4/10).
Przekrwienie miąższu
(10/10).
W niektórych
przypadkach:
ogniskowe nacieki
komórek limfoidalnych
(7/10),
miejscowa martwica
jąder (rozpad i/lub
zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(3/10), obecność
pojedynczych do
zlokalizowanych
skupisk kropli tłuszczu
w miąższu (4/10).
W niektórych
przypadkach:
przekrwienie miąższu
(5/10), obecność
nacieków komórek
limfoidalnych (1/10).
W niektórych
przypadkach: obecność
pojedynczych,
ogniskowych nacieków
limfoidalnych (2/10).
W niektórych
przypadkach:
przekrwienie miąższu
(3/10), nacieki komórek
limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych)
nieznacznego stopnia
(4/10), piankowa
struktura hepatocytów
nieznacznego stopnia
(5/10).
Przekrwienie miąższu
(5/10).
W niektórych
przypadkach:
ogniskowe nacieki
komórek limfoidalnych
(7/10),
miejscowa martwica
jąder (rozpad i/lub
zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(4/10), obecność
pojedynczych do
zlokalizowanych
skupisk kropli tłuszczu
w miąższu (4/10).
W niektórych
przypadkach:
przekrwienie miąższu
(5/10), obecność
nacieków komórek
limfoidalnych (2/10).
W niektórych
przypadkach: obecność
pojedynczych,
ogniskowych nacieków
limfoidalnych (2/10).
W niektórych
przypadkach:
przekrwienie miąższu
(4/10), nacieki komórek
limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych)
nieznacznego stopnia
(3/10), piankowa
struktura hepatocytów
nieznacznego stopnia
(5/10).
Przekrwienie miąższu
(10/10).
W niektórych
przypadkach:
ogniskowe nacieki
komórek limfoidalnych
(6/10),
miejscowa martwica
jąder (rozpad i/lub
zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(6/10), obecność
pojedynczych do
zlokalizowanych
skupisk kropli tłuszczu
w miąższu (7/10).
W niektórych
przypadkach:
przekrwienie miąższu
(3/10), obecność
nacieków komórek
limfoidalnych (2/10).
W niektórych
przypadkach: obecność
pojedynczych,
ogniskowych nacieków
limfoidalnych (2/10).
W niektórych
przypadkach:
przekrwienie miąższu
(4/10), nacieki komórek
limfoidalnych
(ogniskowe i/lub wokół
triad wątrobowych)
nieznacznego stopnia
(4/10), piankowa
struktura hepatocytów
nieznacznego stopnia
(5/10).
Przekrwienie miąższu
(10/10).
W niektórych
przypadkach:
ogniskowe nacieki
komórek limfoidalnych
(7/10),
miejscowa martwica
jąder (rozpad i/lub
zagęszczenie
chromatyny jądrowej)
komórek nabłonka
kanalików nerkowych
(3/10), obecność
pojedynczych do
zlokalizowanych
skupisk kropli tłuszczu
w miąższu (4/10).
W niektórych
przypadkach:
przekrwienie miąższu
(4/10), obecność
nacieków komórek
limfoidalnych (2/10).
W niektórych
przypadkach: obecność
pojedynczych,
ogniskowych nacieków
limfoidalnych (2/10).
23
Dwunastnica
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego
narządu.
Jelito czcze
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego
narządu. W pojedynczym przypadku (1/40, grupa II) odnotowano jedynie ścieńczenie błony śluzowej.
Mięśnie
W niektórych
W niektórych
W niektórych
W niektórych
szkieletowe
przypadkach: obecność
przypadkach: obecność
przypadkach: obecność
przypadkach: obecność
nacieków komórek
nacieków komórek
nacieków komórek
nacieków komórek
limfoidalnych
limfoidalnych
limfoidalnych
limfoidalnych
(okołonaczyniowe i/lub
(okołonaczyniowe i/lub
(okołonaczyniowe i/lub
(okołonaczyniowe i/lub
ogniskowe) (5/10).
ogniskowe) (3/10).
ogniskowe) (3/10).
ogniskowe) (5/10).
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Analiza obecności transgenicznego DNA w narządach, tkankach i przewodzie pokarmowym
niosek
W ramach wykonywanych badań na kurach nieśnych przeprowadzono analizy obecności
transgenicznego DNA, pochodzącego z soi i kukurydzy GM, w wybranych narządach i
tkankach niosek, jak również w poszczególnych odcinkach przewodu pokarmowego.
W Krajowym Laboratorium Pasz Instytutu Zootechniki PIB (pracownia w Szczecinie)
analizowane były następujące próbki materiału biologicznego:
-krew,
-wątroba,
-płuca,
-śledziona,
-oraz treść wola, żołądka, dwunastnicy, jelita czczego, jelita biodrowego i jelita końcowego ze
stekiem.
W grupach żywionych mieszankami paszowymi zawierającymi śrutę z kukurydzy GM,
lub/i poekstrakcyjną śrutę sojową GM, nie stwierdzano obecności w badanych tkankach
(narządach) specyficznego dla danej modyfikacji DNA transgenicznego (Tabela 24). W
przypadku przewodu pokarmowego transgeniczny DNA występował jedynie w początkowej
jego części, tj. w treści wola i żołądka, a w pojedynczych przypadkach także w treści
dwunastnicy (Tabela 24).
Wyniki te zostały potwierdzone w laboratorium Państwowego Instytutu Weterynaryjnego
- PIB w Puławach, w którym w próbkach materiału biologicznego z doświadczenia badano
obecność genów referencyjnych soi (lektyna) i kukurydzy (inwertaza) oraz obecność sekwencji
charakterystycznych dla stosowanych modyfikacji (innych niż oznaczane KLP IZ PIB), tj.
promotora 35S i terminatora NOS. W DNA wizolowanym z krwi, narządów i tkanek (wątroba,
śledziona i nerka) oraz w kale ptaków żywionych mieszankami zawierającymi materiały
paszowe GM nie stwierdzono obecności fragmentów DNA charakterystycznych dla
24
zmodyfikowanej soi i kukurydzy (promotora 35S i terminatora NOS). Wyniki badania treści
wola były w pełni skorelowane z rodzajem stosowanej mieszanki doświadczalnej.
Analizy jaj przeprowadzone w Państwowym Instytucie Weterynaryjnym - PIB w
Puławach oraz Krajowym Laboratorium Pasz IZ PIB w Szczecinie, zebranych po 3 lub 6
miesiącach stosowania diet doświadczalnych, w żadnej z grup nie wykazały obecności
sekwencji DNA charakterystycznych dla kukurydzy oraz soi GM (promotor 35S i terminator
NOS) (Tabela 24). Podobnie nie odnotowano w jajach przypadku występowania fragmentów
naturalnego DNA roślinnego (soi – genu lektyny i kukurydzy – genu inwertazy).
Podsumowując można stwierdzić, że powyższe rezultaty potwierdzają obserwację z
doświadczenia na kurczętach rzeźnych, że kwasy nukleinowe, również transgeniczne
DNA, są u drobiu efektywnie denaturowane, a następnie trawione przez enzymy
(nukleazy) trzustkowe i jelitowe. Wyklucza to praktycznie możliwość transferu
biologicznie aktywnych fragmentów transgenicznego DNA przez barierę jelitową do krwi
oraz innych tkanek i narządów, jak również ich pasaż w formie niestrawionej przez jelita
i wydalanie wraz kałem do środowiska.
Tabela 24.
Wyniki jakościowej analizy DNA w kierunku obecności specyficznych dla GMO
transgenów – kury nieśne (analizy wykonane w KLP IZ-PIB).
Materiał biologiczny
Grupa doświadczalna*
1
2
3
RR
MON810
RR
MON810
RR
Treść wola
-
-
+
n/a**
n/a
Treść żołądka
-
-
+
n/a
n/a
Treść dwunastnicy
-
-
-
n/a
n/a
Treść j. biodrowego
-
-
-
n/a
Treść j. czczego
-
-
-
Treść j. końcowego i steku
-
-
Krew
-
Wątroba
4
MON810
RR
MON810
+
+
+
+
+
-
n/a
-/+ (1 próbka
dodatnia)
-
-
-/+ (3 próbki
dodatnie)
-
n/a
n/a
-
-
-
-
n/a
n/a
-
-
-
-
-
n/a
n/a
-
-
-
-
-
-
n/a
n/a
-
-
-
Płuca
-
-
-
n/a
n/a
-
-
-
Śledziona
-
-
-
n/a
n/a
-
-
-
Jaja
-
-
-
n/a
n/a
-
-
-
+
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
**/ nie analizowano
25
BADANIA NA TUCZNIKACH
W ramach zadania przeprowadzono doświadczenie wzrostowe na tucznikach.
Doświadczenie wykonano na fermie trzody chlewnej Zakładu Doświadczalnego Instytutu
Zootechniki PIB Rudawa Sp. z o.o. w Brzeziu (woj. małopolskie). Materiał zwierzęcy
stanowiły 72 tuczniki pochodzące od loch (pbz x wbp) pokrytych knurem (Pi x Du).
W ramach testu wzrostowego utworzono 6 grup doświadczalnych, dodatkowo do
schematu badań wprowadzając dwie grupy, w których skład mieszanek paszowych został
oparty o śruty zbożowe (grupa 5 i 6) i był zbliżony do mieszanek stosowanych w naszym kraju
w warunkach fermowych.
Do każdej z 6 grup przydzielono po 6 loszek i 6 wieprzków. W ciągu całego
doświadczenia zwierzęta utrzymywane były w kojcach indywidualnych, miały stały dostęp do
wody i otrzymywały dawkowane ilości mieszanki paszowej odpowiednio do masy ciała, którą
kontrolowano co dwa tygodnie. Tucz doświadczalny trwał od około 30 do 110 kg masy ciała.
Zwierzęta otrzymywały izoenergetyczne i izobiałkowe mieszanki paszowe zawierające w
swym składzie jęczmień, pszenicę, kukurydzę, poekstrakcyjną śrutę sojową, otręby pszenne,
dodatki mineralne i witaminowe oraz aminokwasy krystaliczne, pokrywającej zapotrzebowanie
tuczników. We wszystkich grupach mieszanki grower (przeznaczone na okres tuczu od 30 do
60 kg mc) charakteryzowały się koncentracją 12,6 MJ EM oraz zawartością 176 g białka
ogólnego, 9,7 g Liz i 6,1 g Met+Cys, natomiast mieszanki finiszer (przeznaczone na okres
tuczu od 60 do 110kg mc) zawierały 12,6 MJ EM, 160 g białka ogólnego oraz 8,1 g Liz i 5,2 g
Met+Cys. Mieszanki stosowane w doświadczeniu różniły się obecnością lub brakiem
genetycznie zmodyfikowanej kukurydzy MON810 (w ilości 13% w growerze i 10% w
finiszerze) oraz poekstrakcyjnej śruty sojowej MON40-3-2 (w ilości 18% w growerze i 14% w
finiszerze).
Układ doświadczenia był następujący:
Grupa 1 – mieszanka zawierająca śrutę z kukurydzy niemodyfikowanej + poekstrakcyjną śrutę
sojową niemodyfikowaną.
Grupa 2 - mieszanka zawierająca śrutę z kukurydzy niemodyfikowanej + poekstrakcyjną śrutę
sojową GM.
Grupa 3 - mieszanka zawierająca śrutę z kukurydzy GM + poekstrakcyjną śrutę sojową
26
niemodyfikowaną.
Grupa 4 - mieszanka zawierająca śrutę z kukurydzy GM + poekstrakcyjną śrutę sojową GM.
Grupa 5 – mieszanka zawierająca poekstrakcyjną śrutę sojową niemodyfikowaną + śruty
zbożowe.
Grupa 6 - mieszanka zawierająca poekstrakcyjną śrutę sojową GM + śruty zbożowe.
Wyniki analiz zawartości genetycznie zmodyfikowanej soi Roundup Ready i śruty z
kukurydzy MON810, potwierdziły poprawność wykonania doświadczalnych mieszanek
paszowych. Wszystkie pasze wykorzystywane w doświadczeniu poddano analizie ilościowej na
zawartość genetycznie zmodyfikowanych materiałów paszowych (soi i kukurydzy).
Po zakończeniu doświadczenia świnie zostały ubite. Po 24 godzinnym chłodzeniu w
temp. +4ºC prawe półtusze poddano dysekcji. Z okolicy pomiędzy ostatnim kręgiem
piersiowym, a pierwszym lędźwiowym pobrano próbkę longissimus m. w celu wykonania
podstawowej analizy chemicznej (AOAC, 1990) oraz określenia cech jakościowych mięsa.
Barwę mięsa (jasność, wysycenie barwy w kierunku czerwieni oraz w kierunku żółci) mierzono
za pomocą kolorymetru Minolta CR-310. Wskaźnik wodochłonności został określony metodą
opisaną przez Grau i Hamm (1953). Pomiarów kwasowości mięsa dokonano przy pomocy pHmetru przenośnego CP-215 wyposażonego w elektrodę sztyletową o symbolu ES Ag P-306, w
czasie 45 min po uboju oraz po 24 godzinach.
Wyniki
Badania prowadzone w Instytucie Zootechniki PIB
Wskaźniki produkcyjne
W Tabeli 25 przedstawiono średnie dzienne przyrosty masy ciała tuczników oraz
zużycie paszy w przedziale wagowym od 30 do 110 kg. Zarówno w pierwszym jak i drugim
okresie tuczu nie obserwowano różnic między grupami w tempie wzrostu świń (P>0,05).
Średnie przyrosty masy ciała wszystkich tuczników w całym okresie tuczu były bardzo
zbliżone i wynosiły od 824 g do 852 g. Nie stwierdzono także wpływu badanych pasz GM na
zużycie paszy przez tuczniki (Tabela 25).
27
Tabela 25.
Średnie dzienne przyrosty masy ciała i zużycie paszy u tuczników.
Grupa doświadczalna *
Średnie dzienne przyrosty
masy ciała (g):
od 30 do 60 kg
od 60 do 110 kg
od 30 do 110 kg
Średnie zużycie paszy na
przyrost 1 kg m.c. (kg/kg):
od 30 do 60 kg
od 60 do 110 kg
od 30 do 110 kg
Poziom
P
SEM
1
2
3
4
5
6
771
876
824
757
884
831
780
893
852
758
889
835
767
886
837
768
885
837
0,726
0,980
0,698
4,636
5,949
4,813
2,65
3,47
3,16
2,66
3,46
3,16
2,64
3,42
3,12
2,67
3,43
3,15
2,66
3,44
3,14
2,65
3,45
3,15
0,993
0,992
0,993
0,016
0,023
0,018
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
Jakość tuszy i mięsa
Wyniki oceny jakości tusz wieprzowych przedstawiono w Tabeli 26. Analizując
stytystycznie otrzymane rezultaty, nie stwierdzono różnic pomiędzy grupami w żadnym z
badanych parametrów (P>0,05). Zawartości mięsa w wyrębach podstawowych półtuszy była
we wszystkich grupach zbliżona. Mięsność tuczników doświadczalnych wynosiła od około
53,7% w grupach 1, 5 i 6 do 54,6% w grupie 2, natomiast powierzchnia oka polędwicy wahała
się od 59,3 cm2 w grupie 3 do 61,4 cm2 w grupie 6.
Tabela 26.
Jakość tuszy.
Grupa doświadczalna*
Po iom
P
SEM
1
2
3
4
5
6
Wydajność rzeźna (%)
78,26
79,06
79,36
79,22
79,05
79,24
0,529
0,177
Powierzchnia oka polędwicy
(cm2)
Średnia grubość słoniny z 5
pomiarów (cm)
Mięso polędwicy (kg)
60,53
59,92
59,29
61,25
61,08
61,44
0,899
0,575
2,09
2,01
2,22
1,97
1,98
2,13
0,3 5
0,036
7,20
7,41
7,53
7,52
7,67
7,24
0,393
0,073
Mięso szynki (kg)
7,97
8,13
8,36
8,13
8,18
8,09
0,796
0,073
Mięso karkówki (kg)
6,18
6,05
6,34
6,07
6,33
6,04
0,481
0,058
Polędwiczka (kg)
0,52
0,52
0,51
0,54
0,54
0,52
0,576
0,007
Łopatka (kg)
5,24
5,31
5,28
5,27
5,38
5,37
0,934
0,044
Mięso wyrębów
podstawowych (kg)
24,15
24,52
24,89
24,68
24,97
24,31
0,717
0,172
Mięsność tuszy (%)
53,69
54,59
53,89
54,54
53,67
53,66
0,888
0,303
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
28
Analiza stytystyczna nie wykazała różnic w badanych wskaźnikach jakości mięsa
(Tabela 27). We wszystkich grupach doświadczalnych odnotowano zbliżoną kwasowość mięsa,
barwę, wskaźnik wodochłonności oraz podobną zawartość suchej masy, białka i tłuszczu.
Podsumowując wyniki doświadczenia produkcyjnego przeprowadzonego na
tucznikach można stwierdzić, że nie wykazano wpływu ocenianych materiałów paszowych
GM na wskaźniki wzrostowe świń, jak również jakość tusz i mięsa wieprzowego.
Tabela 27.
Jakość i skład chemiczny mięsa (lonissimus m.).
Grupa doświadczalna*
Poziom
SEM
1
2
3
4
5
6
P
pH 45 min po uboju
6,33
6,28
6,28
6,24
6,28
6,38
0,504
0,023
pH po 24h chłodzeniu (+4ºC)
5,58
5,55
5,62
5,59
5,63
5,62
0,451
0,013
Wskaźnik wodochłonności (%)
21,14
21,36
21,04
20,81
21,09
21,15
0,994
0,232
Barwa mięsa: Jasność *L
46,19
46,57
46,40
46,29
46,36
46,24
0,999
0,260
13,42
13,81
13,40
13,37
13,62
13,43
0,570
0,077
2,06
2,19
2,14
2,15
2,14
2,18
0,987
0,050
Sucha masa (%)
24,95
24,91
25,08
25,05
24,81
24,99
0,986
0,105
Białko (%)
22,59
22,81
22,99
22,93
22,37
22,95
0,512
0,106
Tłuszcz (%)
1,14
1,05
1,12
1,04
1,12
1,15
0,806
0,028
Barwa mięsa: Wysycenie w
kierunku czerwieni *a
Barwa mięsa: Wysycenie w
kierunku żółci *b
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
Oznaczenie strawnościowe
W ramach badań wykonanych na tucznikach wykonano także oznaczenie
strawnościowe. Strawność składników pokarmowych mieszanek zawierających lub nie
zawierających materiały paszowe genetycznie zmodyfikowane, tj. poekstrakcyjną śrutę sojową
oraz kukurydzę, określono metodą standardową (Kamiński i wsp., 1991). Doświadczenie
strawnościowe przeprowadzono w grupach doświadczalnych 1–4 (grupa 1 – mieszanka
zawierająca
śrutę
z
kukurydzy
niemodyfikowanej
+
poekstrakcyjną
śrutę
sojową
niemodyfikowaną, grupa 2 - mieszanka zawierająca śrutę z kukurydzy niemodyfikowanej +
poekstrakcyjną śrutę sojową GM, grupa 3 - mieszanka zawierająca śrutę z kukurydzy GM +
poekstrakcyjną śrutę sojową niemodyfikowaną, grupa 4 - mieszanka zawierająca śrutę z
kukurydzy GM + poekstrakcyjną śrutę sojową GM), na 16 wieprzkach (po 4 sztuki w każdej
grupie) w indywidualnych klatkach strawnościowo-bilansowych. Okres wstępny trwał 10 dni, a
okres kolekcji kału 5 dni. Doświadczenie przeprowadzono w dwóch etapach, osobno dla
29
mieszanek na pierwszy okres tuczu (grower), a następnie dla mieszanek na drugi okres tuczu
(finiszer). W etapie pierwszym wieprzki otrzymywały 2,0 kg/dzień mieszanki grower, a w
etapie drugim 2,6 kg mieszanki finiszer. Paszę podawano w dwóch odpasach każdego dnia.
Dostęp do wody dla wszystkich zwierząt był stały. Kał każdego zwierzęcia był ważony
codziennie, a pobrane próbki łączone w indywidualne próbki zbiorcze dla każdej sztuki,
przechowując je w temp. -18º C do czasu wykonania analiz chemicznych. Zawartość
składników podstawowych w próbkach paszy i odchodów oznaczono standardową metodą
chemiczną (AOAC, 1990).
Nie stwierdzono wpływu genetycznie zmodyfikowanego materiału paszowego (śruta
kukurydziana lub/i poekstrakcyjna śruta sojowa) zastosowanego w mieszankach paszowych,
przeznaczonych na pierwszy i drugi okres tuczu świń, na strawność podstawowych składników
pokarmowych (Tabela 28 i 29). Współczynniki strawności składników pokarmowych nie
różniły się statystycznie istotnie pomiędzy grupami (P>0,05). W przypadku żywienia świń
mieszanką growerową średnia dla wszystkich zwierząt wartość współczynnika strawności
suchej masy wynosiła 83,6%, białka ogólnego – 80%, tłuszczu surowego – 53,3%, włókna
surowego – 31,9% oraz substancji bez-N wyciągowych – 92,1%. Natomiast średnia strawność
poszczególnych składników pokarmowych mieszanki finiszerowej wynosiła odpowiednio:
84,9; 78,2; 49,6; 46,5 oraz 92,6%. Obserwowano poprawę strawności włókna surowego, która
u zwierząt w drugim okresie tuczu była o ponad 14 pkt. procentowych wyższa niż u zwierząt
młodszych.
Tabela 28. Współczynniki strawności (%) składników pokarmowych mieszanki paszowej grower
stosowanej w pierwszym okresie tuczu (30 – 60 kg masy ciała)
Grupa doświadczalna*
Poziom
SEM
1
2
3
4
P
Sucha masa
83,64
82,85
83,86
83,97
0,648
0,320
Białko
80,44
79,59
80,17
79,73
0,958
0,555
Tłuszcz
55,69
50,23
50,31
57,15
0,112
1,316
Włókno
29,08
34,31
35,65
28,50
0,176
1,419
Substancje bez-N wyciągowe
91,85
92,02
92,21
92,38
0,701
0,159
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
30
Tabela 29. Współczynniki strawności (%) składników pokarmowych mieszanki paszowej finiszer
stosowanej w drugim okresie tuczu (60 – 110 kg masy ciała)
Grupa doświadczalna*
Poziom
4
SEM
P
1
2
3
Sucha masa
84,73
84,24
85,57
84,9
0,400
0,269
Białko
77,05
77,21
80,03
78,52
0,256
0,588
Tłuszcz
49,92
51,18
45,78
51,62
0,805
2,162
Włókno
45,50
46,96
47,26
46,27
0,949
1,047
Substancje bez-N wyc.
92,64
92,23
92,87
92,64
0,624
0,166
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Badania wykonane w Państwowym Instytucie Weterynaryjnym - PIB
Ocena wskaźników hematologicznych krwi i komórkowej odpowiedzi immunologicznej
W ramach podjętych badań dokonano oceny wpływu pasz GM na parametry
charakteryzujące status immunologiczny świń. Stan odporności komórkowej tuczników badano
pobierając próbki od 6 zwierząt z każdej grupy (3 loszki i 3 wieprzki). Krew do badań
immunologicznych wybranych wskaźników nieswoistej odporności komórkowej pobierano
dwukrotnie, tj. przed rozpoczęciem doświadczenia i po skarmianiu różnymi rodzajami pasz
GM. W ramach przeprowadzonych badań immunologicznych oceniano skład procentowy
poszczególnych subpopulacji limfocytów krwi obwodowej z użyciem cytometrii przepływowej
i specyficznego panelu przeciwciał monoklonalnych umożliwiających rozpoznawanie komórek
CD3+ (limfocyty T), CD4+ (limfocyty Th; pomocnicze) i CD8+ (limfocyty Tc/s;
cytotoksyczno-supresorowe). Ponadto, wykonano analizę ogólnych wskaźników statusu
zdrowotnego, tj. wskaźników erytrocytarnych (liczba czerwonych krwinek – RBC, ilość
hemoglobiny – HGB, hematokryt – HCT, średnia objętość erytrocytu – MCV, średni ciężar i
stężenie hemoglobiny w jednej krwince – MCH i MCHC) i trombocytarnych (liczba płytek
krwi – PLT i ich średnia objętość – MVP) oraz wybranych parametrów odporności
komórkowej, takich jak liczba białych ciałek krwi (WBC) i leukogram, tj. jakościowy skład
poszczególnych subpopulacji leukocytów z podziałem na granulocyty obojętnochłonne –
PMNL, komórki średniej wielkości – MID (sumaryczna wartość monocytów, bazofilów,
eozynofilów w jednostce objętości krwi) i limfocytów – LYM. Badane wskaźniki
immunologiczne odzwierciedlają charakter zmian w ramach nieswoistej odporności
komórkowej.
W badaniach tzw. ogólnych wskaźników statusu zdrowotnego, tj. parametrów układu
erytrocytarnego i trombocytarnego nie obserwowano różnic pomiędzy grupami tuczników
31
(Tabela 30, 31), żywionych dietą bez udziału lub z udziałem badanych materiałów paszowych
GM. Podobnie, brak jakichkolwiek zależności stwierdzono też w odniesieniu do pozostałych
ocenianych parametrów nieswoistej odporności komórkowej, tj. ogólnej liczby leukocytów we
krwi (WBC), leukogramu (LYM, PMNL, MID) i immunofenotypu limfocytów z podziałem na
komórki CD3+, CD4+ i CD8+ (Tabela 32, 33).
Tabela 30. Średnie wartości wskaźników erytrocytarnych krwi obwodowej u tuczników.
0
RBC (x 1012/l)
1
0
MCV (fl)
1
0
HCT (l/l)
1
0
HGB (mmol/l)
1
0
MCH (fmol)
1
0
MCHC (mmol/l)
1
1
5,84
±0,56
5,43
±0,26
63,5
±0,71
66,0
±2,83
0,37
±0,04
0,36
±0,03
11,05
±1,48
10,45
±0,78
1,85
±0,07
1,85
±0,07
29,55
±0,92
29,20
±0,42
2
5,62
±0,32
5,41
±0,52
71,5
±0,71
74,0
±11,31
0,4
±0,04
0,39
±0,04
11,35
±0,49
11,50
±1,13
1,95
±0,07
2,15
±0,35
28,25
±1,48
29,25
±0,07
Grupa doświadczalna*
3
4
4,98
6,08
±0,8
±0,04
5,07
6,10
±0,6
±0,13
64,0
66,0
±0,0
±0,0
67,0
64,0
±1,41
±4,24
0,3
0,4
±0,05
±0,0
0,33
0,39
±0,08
±0,03
8,85
11,7
±1,48
±0,28
9,45
11,35
±2,19
±1,34
1,8
1,85
±0,0
±0,07
1,9
1,85
±0,0
±0,21
29,45
29,0
±0,07
±0,99
28,8±
29,05
0,14
±1,34
5
6,75
±1,02
6,01
±1,23
61,5
±2,12
60,50
±24,12
0,38
±0,05
0,38
±0,14
11,95
±1,48
11,4
±4,25
1,85
±0,07
1,75
±0,71
31,0
±0,14
30,0
±12,44
6
5,11
±0,19
4,82
±0,12
65,0
±7,07
65,0
±1,41
0,33
±0,02
0,27
±0,01
10,10
±0,42
7,85
±0,07
1,9
±0,14
1,85
±0,07
30,40
±0,99
29,05
±0,49
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
0 – wartości wyjściowe przed rozpoczęciem doświadczenia; 1 – wartości po żywieniu paszami GM.
Tabela 31. Średnie wartości wskaźników trombocytarnych krwi obwodowej u tuczników.
PLT (109/L)
0
1
0
MPV (fl)
1
1
211,5
±152,03
2
361,5
±126,57
198,5
±17,68
5,7
±0,42
5,7
±0,28
342,5
±36,06
5,65
±0,35
5,7
±0,14
Grupa doświadczalna *
3
4
158,0
293,5
±39,6
±44,55
160,0
±72,12
5,9
±0,57
5,7
±0,28
380,5
±44,55
5,5
±0,0
5,6
±0,0
5
120,0
±32,53
6
115,0
±50,91
88,0
±40,97
5,85
±0,21
5,75
±2,29
119,5
±15,85
5,75
±0,07
5,9
±0,57
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
0 – wartości wyjściowe przed rozpoczęciem doświadczenia; 1 – wartości po żywieniu paszami GM.
32
Tabela 32. Średnie wartości wskaźników leukocytarnych krwi obwodowej u tuczników.
WBC (x 109/L)
0
1
0
PMNL (%)
1
0
LYM (%)
1
0
MID (%)
1
1
11,45
±3,15
10,85
±1,06
46,5
±16,26
2
9,65
±2,59
8,75
±3,04
31,5
±4,95
60,0
±4,24
43,5
±14,85
31,0
±4,24
10,0
±1,41
9,0
±0,0
38,0
±11,31
57,5
±6,36
49,5
±10,61
11,0
±1,41
12,5
±0,71
Grupa doświadczalna *
3
4
7,3
10,05
±3,96
±0,21
8,65
9,65
±4,17
±1,48
54,0
32,0
±12,73
±5,66
52,5
±6,36
36,5
±3,54
35,5
±10,61
11,0
±2,83
10,5
±2,12
33,0
±0,0
57,5
±4,95
55,0
±2,83
10,5
±0,71
12,0
±2,83
5
15,75
±1,63
18,1
±6,44
51,0
±4,24
6
15,05
±3,18
13,10
±0,85
49,0
±9,9
51,5
±20,38
40,0
±4,24
37,5
±15,57
9,0
±0,0
11,0
±4,17
46,5
±2,12
41,5
±7,78
41,0
±2,83
9,5
±2,12
12,5
±0,71
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
0 – wartości wyjściowe przed rozpoczęciem doświadczenia; 1 – wartości po żywieniu paszami GM
Tabela 33. Zmiany w subpopulacjach limfocytów krwi obwodowej u tuczników.
CD3+ (%)
0
1
CD4+ (%)
0
1
CD8+ (%)
0
1
1
48,93
±3,66
2
48,47
±0,45
46,87
±5,33
29,43
±2,38
32,1
±5,1
37,30
±3,15
38,92
±3,03
43,33
±6,52
29,83
±3,06
33,07
±6,39
34,90
±5,13
37,0
±4,03
Grupa doświadczalna *
3
4
47,47
50,10
±2,8
±2,1
54,27
±2,83
30,50
±6,59
27,30
±2,10
36,0
±9,10
37,73
±3,15
57,97
±4,83
32,03
±1,8
28,07
±3,40
27,35
±3,30
29,30
±5,14
5
48,27
±5,49
6
49,57
±0,9
55,47
±2,85
30,27
±1,66
29,37
±2,40
28,60
±6,68
25,70
±9,82
56,91
±4,50
29,80
±1,35
29,2
±4,10
21,15
±4,33
20,10
±1,25
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
0 – wartości wyjściowe przed rozpoczęciem doświadczenia; 1 – wartości po żywieniu paszami GM
Ocena humoralnej odpowiedzi immunologicznej
W czasie trwania doświadczenia dokonano oceny odpowiedzi immunologicznej po
szczepieniach biopreparatami atenuowanymi lub inaktywowanymi.
33
Oceny dokonano
wykorzystując szczepionki przeciw chorobie Aujeszkyego oraz zespołowi rozrodczooddechowemu PRRS. Dokonano także oceny klinicznej stanu zdrowia świń. Grupy 1 - 4
zaszczepiono biopreparatem Akipor 6.3 (atenuowana szczepionka przeciw chorobie
Aujeszkyego) w dwóch dawkach. Dawki w ilości 2 ml podano domięśniowo w odstępie 5
tygodni. Grupy świń 5 i 6 zaszczepiono biopreparatem Progressis (inaktywowana szczepionka
przeciw zespołowi rozrodczo-oddechowemu – PRRS) w tych samych terminach co
szczepionka wymieniona powyżej. Każdej grupie doświadczalnej obejmującej 8 zwierząt
(szczepionych) odpowiadały grupy kontrolne obejmujące 4 zwierzęta (nieszczepione). Krew do
badań serologicznych pobrano od wszystkich zwierząt w czterech terminach, w odstępach 3tygodniowych. Próbki krwi w kierunku obecności przeciwciał dla wirusa PRRS przebadano
testem ELISA, a poziom przeciwciał humoralnych określono w zakresie od 0 (brak
przeciwciał) do 6 (maksymalny poziom przeciwciał). Surowice krwi przeznaczone do badania
w kierunku poziomu przeciwciał dla choroby Aujeszkyego przebadano testem seroneutralizacji
i ustalono miana ND50 poszczególnych surowic.
Wyniki odpowiedzi immunologicznej po wykonanych szczepieniach tuczników, przeciw
chorobie Aujeszkyego i PRRS, przedstawiono w tabeli 34. Nie odnotowano różnic między
średnim poziomem miana przeciwciał w grupach zwierząt żywionych dietą opartą o materiały
paszowe GM oraz mieszanką kontrolną. Analiza uzyskanych danych wskazuje, że szczepienia
biopreparatem Akipor 6.3 we wszystkich grupach doświadczalnych uwidoczniły się w postaci
wzrostu miana swoistych przeciwciał humoralnych dla choroby Aujeszkyego. We wszystkich
grupach doświadczalnych miano to osiągnęło najwyższe wartości w 1 i 2 próbkobraniu. W
grupach kontrolnych nie wykazano obecności przeciwciał. Nie można jednakże wyciągnąć
obiektywnych wniosków w zakresie wpływu materiałów paszowych GM na efektywność
szczepień przeciwko PRRS, gdyż w trakcie doświadczenia okazało się bowiem, że świnie w
chlewni uległy naturalnemu zakażeniu wirusem PRRS w związku z czym u wszystkich
zwierząt stwierdzano obecność swoistych przeciwciał. Nie było możliwości odróżnienia
przeciwciał indukowanych poprzez podanie szczepionki od przeciwciał powstałych po infekcji.
Podsumowując
wykonane
oznaczenia
hematologiczno-immunologiczne
krwi
tuczników można stwierdzić, że uzyskane wyniki, wskazują na brak wpływu stosowanych
materiałów paszowych GM (poekstrakcyjna śruta sojowa, śruta kukurydziana) na
przebieg komórkowych i humoralnych procesów immunologicznych.
34
Tabela 34.
Wyniki badań serologicznych krwi tuczników w kierunku obecności i poziomu
przeciwciał dla wirusa Aujeszkyego i PRRS – średnie wartości przeciwciał.
Grupa doświadczalna*
3
4
Osobniki
szczepione
(Aujeszky)
Osobniki
kontrolne
Osobniki
szczepione
(PRRS)
<0,3
<0,3
<0,3
<0,3
<0,3
<0,3
4,9
6,0
4,0
3,7
Pobranie po I
szczepieniu
0,40
<0,3
0,38
<0,3
0,27
<0,3
<0,3
<0,3
5,5
4,5
5,0
4,7
Pobranie po II
szczepieniu
1,48
<0,3
1,25
<0,3
1,36
<0,3
1,08
<0,3
4,6
3,2
3,4
4,0
Ostatnie
pobranie
1,08
<0,3
0,66
<0,3
0,8
<0,3
0,36
<0,3
2,0
2,0
1,1
2,0
Osobniki
kontrolne
Osobniki
kontrolne
<0,3
Osobniki
szczepione
(PRRS)
Osobniki
szczepione
(Aujeszky)
<0,3**
Pobranie przed
szczepieniem
Osobniki
kontrolne
Osobniki
kontrolna
6
Osobniki
szczepione
(Aujeszky)
5
Osobniki
kontrolne
2
Osobniki
szczepione
(Aujeszky)
1
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
**/ log10 ND50
Analiza mikroflory przewodu pokarmowego
Badania mikroflory przewodu pokarmowego dotyczyły przede wszystkim oceny
możliwości transferu transgenicznego DNA do wybranych grup mikroorganizmów.
Bezpośrednio po uboju, od 6 zwierząt (3 loszki i 3 wieprzki) z każdej grupy, pobrano próbki
treści pokarmowej z jelita ślepego i grubego w celu oceny mikroflory przewodu pokarmowego
pod względem liczebności takich gatunków bakterii jak Escherichia coli oraz Enterococcus
faecalis i Enterococcus faecium, a także zbadania możliwości transferu transgenicznego DNA
do wybranych grup mikroorganizmów. Bakterie izolowano z treści wybranych odcinków
przewodu pokarmowego. Do hodowli mikroorganizmów wykorzystywano podłoża selektywne.
Dla E. coli stosowano podłoże TBX – pożywkę tryptono-żółciowo glukuronidynową, a dla
Enterococcus podłoże Slanetz’a z azydkiem sodu. Do namnażania drobnoustrojów wybrano
podłoże płynne BHI – wyciąg mózgowo – sercowy.
W żadnej z grup doświadczalnych nie stwierdzono obecności w bakteryjnym DNA
elementów transgenicznych: promotora 35S i terminatora NOS. Badania dotyczące składu
ilościowego wybranych mikroorganizmów nie wykazały różnic ilościowych pomiędzy grupami
żywionymi mieszankami paszowymi z materiałem genetycznie zmodyfikowanym lub bez.
Podsumowując można stwierdzić, że stosowanie w żywieniu tuczników diet zawierających
badane materiały paszowe GM nie wpłynęło na skład ilościowy mikroflory przewodu
pokarmowego, jak również nie stwierdzono transferu transgenicznego DNA do bakterii
zasiedlających badane odcinki przewodu pokarmowego.
35
Wyniki badań histopatologicznych
Wyniki badań histopatologicznych przedstawiono w tabeli 35. Bezpośrednio po uboju,
od 6 zwierząt (3 loszki i 3 wieprzki) z każdej grupy pobrano próbki mięsa (mięsień smukły),
narządów wewnętrznych (wątroba, nerki, śledziona, trzustka) oraz wybranych odcinków
przewodu pokarmowego (dwunastnica, jelito czcze) w celu wykonania badań histologicznych.
Z utrwalonego w 10% roztworze zbuforowanej formaliny materiału wykonywano skrawki
parafinowe, które następnie barwiono metodą hematoksylina-eozyna (H-E).
W niektórych przypadkach obserwowano zmiany histopatologiczne w wątrobie,
nerkach, śledzionie, trzustce, dwunastnicy, jelicie czczym i mięśniach (Tabela 35). Analiza
uzyskanych wyników pozwala na stwierdzenie, że występowanie tych zmian nie było związane
ze sposobem żywienia, tj. obecnością w diecie materiałów paszowych GM. Odnotowane
zmiany stanowią prawdopodobnie efekt intensywnego żywienia, w tym dużej zawartości białka
i energii metabolicznej w dietach, oraz wysokiego tempa metabolizmu u szybko rosnących
tuczników.
Podsumowując
należy
stwierdzić,
że
żywienie
tuczników
dietami
zawierającymi badane materiały paszowe GM nie miało negatywnego wpływu na obraz
histopatologiczny narządów wewnętrznych.
Tabela 35. Wyniki badań histopatologicznych narządów wewnętrznych tuczników.
1
2
Grupa doświadczalna*
3
4
5
Wątroba
Przekrwienie
miąższu (3/6),
obecność
ogniskowych
nacieków
komórek
limfoidalnych
nieznacznego
stopnia (2/6)
oraz rozlanych
nacieków
komórek
plazmatycznych
umiarkowanego
stopnia (2/6).
Przekrwienie
miąższu (3/6),
obecność
ogniskowych
nacieków
komórek
limfoidalnych
nieznacznego
stopnia (1/6).
Przekrwienie
miąższu (6/6),
obecność
ogniskowych
nacieków
komórek
limfoidalnych
nieznacznego
stopnia (2/6).
Przekrwienie
miąższu (3/6),
obecność
ogniskowych
nacieków
komórek
limfoidalnych
nieznacznego
stopnia (1/6)
oraz rozlanych
nacieków
komórek
plazmatycznych
umiarkowanego
stopnia (1/6).
Przekrwienie
miąższu (3/6),
obecność
ogniskowych
nacieków
komórek
limfoidalnych
nieznacznego
stopnia (1/6).
Przekrwienie
miąższu (3/6),
obecność
ogniskowych
nacieków
komórek
limfoidalnych
nieznacznego
stopnia (1/6)
oraz rozlanych
nacieków
komórek
plazmatycznych
umiarkowanego
stopnia (2/6).
Nerki
Przekrwienie
miąższu
nieznacznego
stopnia (3/6),
ogniskowe
nacieki
komórek
limfoidalnych
(3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6),
ogniskowe
nacieki
komórek
limfoidalnych
(3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6),
ogniskowe
nacieki
komórek
limfoidalnych
(3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6),
ogniskowe
nacieki
komórek
limfoidalnych
(3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6),
ogniskowe
nacieki
komórek
limfoidalnych
(3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6),
ogniskowe
nacieki
komórek
limfoidalnych
(3/6).
36
Śledziona
Przekrwienie
miąższu
nieznacznego
stopnia (3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6).
Przekrwienie
miąższu
nieznacznego
stopnia (3/6).
Trzustka:
Obecność
pojedynczych,
ogniskowych
nacieków
komórek
limfoidalnych
(5/6), obecność
nacieków
komórek
limfoidalnych
umiarkowanego
stopnia (1/6).
Nagromadzenie
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej od
nieznacznego
do silnego
stopnia (6/6),
martwica mniej lub
bardziej
głęboka błony
śluzowej (3/6).
Obecność
pojedynczych,
ogniskowych
nacieków
komórek
limfoidalnych
(5/6), obecność
nacieków
komórek
limfoidalnych
umiarkowanego
stopnia (1/6).
Nagromadzenie
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej od
nieznacznego
do silnego
stopnia (6/6),
martwica mniej lub
bardziej
głęboka błony
śluzowej (3/6).
Obecność
pojedynczych,
ogniskowych
nacieków
komórek
limfoidalnych
(3/6), obecność
nacieków
komórek
limfoidalnych
umiarkowanego
stopnia (1/6).
Nagromadzenie
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej od
nieznacznego
do silnego
stopnia (6/6),
martwica mniej lub
bardziej
głęboka błony
śluzowej (3/6).
Obecność
pojedynczych,
ogniskowych
nacieków
komórek
limfoidalnych
(4/6).
Nagromadzenie
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej od
nieznacznego
do silnego
stopnia (6/6),
martwica mniej lub
bardziej
głęboka błony
śluzowej (3/6).
Obecność
pojedynczych,
ogniskowych
nacieków
komórek
limfoidalnych
(4/6), obecność
nacieków
komórek
limfoidalnych
umiarkowanego
stopnia (1/6).
Nagromadzenie
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej od
nieznacznego
do silnego
stopnia (6/6),
martwica mniej lub
bardziej
głęboka błony
śluzowej (3/6).
Obecność
pojedynczych,
ogniskowych
nacieków
komórek
limfoidalnych
(5/6), obecność
nacieków
komórek
limfoidalnych
umiarkowanego
stopnia (1/6).
Nagromadzenie
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej od
nieznacznego
do silnego
stopnia (6/6),
martwica mniej lub
bardziej
głęboka błony
śluzowej (3/6).
Jelito czcze
Przekrwienie
błony śluzowej
i warstwy
podśluzowej od
nieznacznego
do
umiarkowanego
stopnia (3/6),
zwiększona
liczba (niekiedy
znacznie)
granulocytów
kwasochłonn. w
błonie śluzowej
(3/6),
nagromadzenie
silnego stopnia
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej
(3/6).
Przekrwienie
błony śluzowej i
warstwy
podśluzowej od
nieznacznego
do
umiarkowanego
stopnia (3/6),
zwiększona
liczba (niekiedy
znacznie)
granulocytów
kwasochłonn. w
błonie śluzowej
(3/6),
nagromadzenie
silnego stopnia
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej
(3/6).
Przekrwienie
błony śluzowej
i warstwy
podśluzowej od
nieznacznego
do
umiarkowanego
stopnia (3/6),
zwiększona
liczba (niekiedy
znacznie)
granulocytów
kwasochłonn. w
błonie śluzowej
(3/6),
nagromadzenie
silnego stopnia
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej
(3/6).
Przekrwienie
błony śluzowej i
warstwy
podśluzowej od
nieznacznego
do
umiarkowanego
stopnia (3/6),
zwiększona
liczba (niekiedy
znacznie)
granulocytów
kwasochłonn. w
błonie śluzowej
(3/6),
nagromadzenie
silnego stopnia
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej
(3/6).
Przekrwienie
błony śluzowej i
warstwy
podśluzowej od
nieznacznego
do
umiarkowanego
stopnia (3/6),
zwiększona
liczba (niekiedy
znacznie)
granulocytów
kwasochłonn. w
błonie śluzowej
(3/6), martwica
błony śluzowej
(1/6).
Mięśnie
szkieletowe
Nacieki
komórek
limfoidalnych
nieznacznego
stopnia (2/6).
Nacieki
komórek
limfoidalnych
nieznacznego
stopnia (2/6).
Przekrwienie
błony śluzowej i
warstwy
podśluzowej od
nieznacznego
do
umiarkowanego
stopnia (3/6),
zwiększona
liczba (niekiedy
znacznie)
granulocytów
kwasochłonn. w
błonie śluzowej
(3/6),
nagromadzenie
silnego stopnia
tkanki
limfatycznej w
błonie śluzowej
i warstwie
podśluzowej
(3/6).
Nie stwierdzono
istotnych
odstępstw od
prawidłowego
obrazu
histologicznego
tkanki.
Nacieki
komórek
limfoidalnych
nieznacznego
stopnia (1/6).
Nacieki
komórek
limfoidalnych
nieznacznego
stopnia (2/6).
Nie stwierdzono
istotnych
odstępstw od
prawidłowego
obrazu
histologicznego
tkanki.
Dwunastnica
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
37
Analiza obecności transgenicznego DNA w narządach, tkankach i przewodzie pokarmowym
tuczników
Tabela 36 zawiera wyniki, przeprowadzonej w Krajowym Laboratorium Pasz Instytutu
Zootechniki PIB (Pracownia w Szczecinie), analizy obecności transgenicznego DNA,
pochodzącego z badanych materiałów paszowych genetycznie zmodyfikowanych w wybranych
narządach i odcinkach przewodu pokarmowego tuczników. Bezpośrednio po uboju, od 6
zwierząt (3 loszki i 3 wieprzki) z każdej grupy, pobrano próbki mięsa (longissimus m.),
narządów wewnętrznych (płuca, wątroba, śledziona), krwi oraz treść pokarmową z wybranych
odcinków przewodu pokarmowego (żołądek, dwunastnica, jelito cienkie, jelito ślepe, jelito
grube) do oznaczeń obecności transgenicznego DNA. W celu stwierdzenia bądź braku
obecności produktu specyficznego dla RR lub MON810 zastosowano badanie jakościowe
GMO metodą PCR.
DNA pochodzące z soi (RR) i/lub kukurydzy (MON810) stwierdzono w treści żołądka
wszystkich zwierząt otrzymujących dany materiał w mieszance paszowej. Obecność DNA
transgenicznego w dwunastnicy stwierdzono tylko u osobników z grupy 2 oraz pojedynczych
zwierząt z grupy 3 i 4. W treści pokarmowej dwunastnicy zwierząt z grupy 5 i 6 oraz w treści
dalszych odcinków przewodu pokarmowego wszystkich zwierząt nie obserwowano obecności
DNA specyficznego dla soi i/lub kukurydzy GM. Nie odnotowano także obecności DNA
transgenicznego we krwi ani w żadnym z badanych narządów (Tabela 36). Podsumowując
można stwierdzić, że DNA, także transgeniczne, jest skutecznie trawione przez enzymy
przewodu pokarmowego, co wyklucza jego przenikanie przez ściany jelita do krwi i narządów
wewnętrznych organizmu oraz wydalanie wraz z kałem do środowiska.
Tabela 36.
Wyniki jakościowej analizy DNA w kierunku obecności specyficznych dla GMO
transgenów – materiał biologiczny z doświadczenia na tucznikach (analizy wykonane w
KLP IZ-PIB).
Materiał biologiczny
Treść żołądka
1
2
RR
MON
810
-
-
3
RR
MON
810
RR
+
n/a**
n/a
Grupa doświadczalna*
4
MON
RR
810
5
MON
810
+
+
+
+/-
+/-
+/-
(2 próbki
(3 próbki
(2 próbki
6
RR
MON
810
RR
MON
810
+
-
+
-
-
-
-
-
Treść dwunastnicy
-
-
+
n/a
n/a
dodatnie)
dodatnie)
dodatnie)
Treść jelita cienkiego
-
-
-
n/a
n/a
-
-
-
-
-
-
-
Treść jelita ślepego
-
-
-
n/a
n/a
-
-
-
-
-
-
-
Treść jelita grubego
-
-
-
n/a
n/a
-
-
-
-
-
-
-
Krew
-
-
-
n/a
n/a
-
-
-
-
-
-
-
38
Wątroba
-
-
-
n/a
n/a
-
-
-
-
-
-
-
Śledziona
-
-
-
n/a
n/a
-
-
-
-
-
-
-
Płuco
-
-
-
n/a
n/a
-
-
-
-
-
-
-
Mięsień
-
-
-
n/a
n/a
-
-
-
-
-
-
-
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna, 3 - śruta sojowa
konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa konwencjonalna + śruty zbożowe, 6 – śruta
sojowa GM + śruty zbożowe
**/ nie analizowano
W Tabeli 37 przedstawiono wyniki badań porównawczych, określających obecność
DNA transgenicznego w treści przewodu pokarmowego oraz w tkankach, wykonanych w
Państwowym Instytucie Weterynaryjnym - PIB w Puławach. Bezpośrednio po uboju, od 6
zwierząt (3 loszki i 3 wieprzki) z każdej grupy pobrano próbki mięsa (miesień najdłuższy
grzbietu, mięsień smukły) narządów wewnętrznych (płuca, nerka, wątroba, śledziona, trzustka),
krwi, treści wybranych odcinków przewodu pokarmowego (żołądek, dwunastnica, jelito
cienkie, jelito ślepe, jelito grube) oraz kału w kierunku wykrywania transgenów: terminatora
NOS, promotora 35S oraz genów referencyjnych: lektyny dla soi i inwertazy dla kukurydzy.
W wyizolowanym DNA pochodzącym z tkanek, narządów oraz treści układu
pokarmowego, z wyjątkiem treści żołądka, nie wykryto obecności poszukiwanych elementów
genetycznych: promotora 35S, terminatora NOS, genu lektyny i genu inwertazy. Obecność
poszukiwanych genów w treści żołądka była ściśle związana z rodzajem paszy stosowanej w
odpowiednich grupach zwierząt. Charakterystyczne dla soi sekwencje genetyczne GM tj.
promotor 35S i terminator NOS zidentyfikowano w grupie 2, 4 i 6, w których do żywienia
zwierząt wykorzystywano soję genetycznie zmodyfikowaną. W grupie 3 wykryto promotor 35S
występujący w kukurydzy MON810. Ponadto w zawartości żołądka wszystkich grup tuczników
badania wykazały obecność genów referencyjnych lektyny dla soi oraz inwertazy dla
kukurydzy. W kolejnych odcinkach przewodu pokarmowego nie wykryto żadnego z
poszukiwanych elementów łańcucha DNA (Tabela 37).
Tabela 37. Wyniki analizy obecności transgenicznego DNA – materiał biologiczny z doświadczenia na
tucznikach (analizy wykonane w PIWet-PIB).
35 S
lektyna
inwertaza
NOS
35 S
lektyna
inwertaza
NOS
35 S
lektyna
inwertaza
NOS
35S
NOS
35 S
lektyna
-
-
+
+
+
+
+
+
-
+
+
+
+
+
+
+
-
-
+
n/a**
+
+
+
n/a
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
39
lektyna
NOS
6
inwertaza
5
inwertaza
Grupa doświadczalna*
4
lektyna
3
35 S
2
NOS
Treść żołądka
Treść
dwunastnicy
Treść jelita
cienkiego
Treść jelita
ślepego
1
inwertaza
Materiał
biologiczny
Treść jelita
grubego
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Kał
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Krew
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Nerka
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Wątroba
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Śledziona
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Trzustka
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
Płuco
Mięsień
najdłuższy
grzbietu
Mięsień
smukły
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
n/a
-
-
-
n/a
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM, 5 – śruta sojowa
konwencjonalna + śruty zbożowe, 6 – śruta sojowa GM + śruty zbożowe
**/ nie analizowano
Podsumowując można stwierdzić, DNA badanych materiałów paszowych, w tym
DNA transgeniczne jest u świń skutecznie trawione przez nukleazy trzustkowe i jelitowe.
Wyklucza to praktycznie możliwość transferu biologicznie aktywnych fragmentów
transgenicznego DNA przez barierę jelitową do tkanek i narządów organizmu, jak
również ich pasaż w formie niestrawionej przez jelita i wydalanie wraz kałem do
środowiska.
40
WYNIKI DOŚWIADCZEŃ ŻYWIENIOWYCH NA LOCHACH
W ramach realizacji badań przeprowadzono doświadczenie nad wpływem pasz GM na
wskaźniki reprodukcyjne loch, wyniki odchowu prosiąt oraz status zdrowotny zwierząt.
Materiał doświadczalny stanowiły 24 lochy (pbz x wbp) pokryte knurem (Du x Pi). Lochy po
pokryciu podzielono na 4 grupy (po 6 sztuk). W ciągu całego doświadczenia lochy
utrzymywane były w kojcach indywidualnych, miały stały dostęp do wody i otrzymywały
dawkowane ilości paszy, zgodnie z zaleceniami żywienia świń. Użyte mieszanki paszowe były
we wszystkich grupach izobiałkowe i izoenergetyczne, a w ich skład wchodził jęczmień,
pszenica, otręby pszenne, susz z lucerny, śruta kukurydziana, poekstrakcyjna śruta sojowa, olej
rzepakowy, dodatki mineralne i witaminowe oraz aminokwasy krystaliczne. Mieszanki
przeznaczone dla loch niskoprośnych zawierały 11,8 MJ EM oraz 132 g białka ogólnego, 6,8 g
Liz i 4,5 g Met+Cys, natomiast mieszanki paszowe przeznaczone dla loch wysokoprośnych i
karmiących zawierały 12,5 MJ EM, 162 g białka ogólnego oraz 8,2 g Liz i 5,5 g Met+Cys.
Mieszanki paszowe stosowane w doświadczeniu różniły się obecnością lub brakiem
genetycznie zmodyfikowanej kukurydzy MON810 (w ilości 5% dla loch niskoprośnych i 8%
dla wysokoprośnych i karmiących) oraz poekstrakcyjnej śruty sojowej MON40-3-2 (w ilości
4% dla loch niskoprośnych i 14% dla wysokoprośnych i karmiących).
Układ doświadczenia był następujący:
Grupa 1 – mieszanka zawierająca śrutę z kukurydzy niemodyfikowanej + poekstrakcyjną śrutę
sojową niemodyfikowaną.
Grupa 2 - mieszanka zawierająca śrutę z kukurydzy niemodyfikowanej + poekstrakcyjną śrutę
sojową GM.
Grupa 3 - mieszanka zawierająca śrutę z kukurydzy GM + poekstrakcyjną śrutę sojową
niemodyfikowaną.
Grupa 4 - mieszanka zawierająca śrutę z kukurydzy GM + poekstrakcyjną śrutę sojową GM.
Masę ciała loch kontrolowano w dniu krycia, w 100 dniu ciąży, po wyproszeniu oraz po
odsadzeniu. Badaniami objęto także potomstwo loch. Od 7 dnia życia prosięta otrzymywały
izobiałkowe i izoenergetyczne mieszanki paszowe zawierające śrutę z identycznych odmian
kukurydzy (w ilości 10%) i poekstrakcyjnej śruty sojowej (w ilości 26%), jak w mieszankach
dla loch, od których dany miot pochodził. Mieszanki te charakteryzowały się koncentracją 13,3
MJ EM oraz zawartością 214 g białka ogólnego, 13,2 g Liz i 7,8 g Met+Cys. W skład
41
mieszanek dla prosiąt wchodziły następujące materiały paszowe: śruta jęczmienna, śruta
pszenna, śruta kukurydziana, poekstrakcyjna śruta sojowa, mleko odtłuszczone w proszku,
serwatka suszona, dodatki mineralne i witaminowe oraz aminokwasy krystaliczne. Wszystkie
prosięta odsadzano od loch w 28 dniu życia, a następnie odchowywano w kojcach zbiorowych
(każdy miot osobno) do 84 dnia życia.
Wyniki
Badania prowadzone w Instytucie Zootechniki PIB
Wskaźniki reprodukcyjne i wyniki odchowu prosiąt
Średnia masa ciała loch przy kryciu, tj. przy rozpoczęciu doświadczenia była we
wszystkich grupach zbliżona i wynosiła od 206,5 do 225,5 kg (Tabela 38). Różnica w wielkości
przyrostu masy ciała loch, mierzona we wszystkich grupach pomiędzy dniem krycia i
pierwszym dniem po wyproszeniu, wynosiła jedynie 3,8 kg. W czasie laktacji lochy straciły od
9,9 (grupa 2) do 12,6 kg (grupa 4), a po zakończeniu całego cyklu reprodukcyjnego masa
średnia ciała loch we wszystkich grupach wzrosła o 19,1 – 22,3 kg w stosunku do masy ciała na
początku doświadczenia. W żadnym okresie badanego cyklu masy ciała loch nie różniły się
statystycznie istotnie pomiędzy grupami. We wszystkich grupach lochy były żywione
dawkowanymi ilościami paszy i pobrały zbliżoną ilość paszy, która w okresie ciąży wynosiła
około 242-244 kg/szt., a w okresie laktacji od 155 do 160 kg/szt. (Tabela 38). Nie stwierdzono
różnic w wielkości pobrania paszy przez lochy w poszczególnych okresach cyklu
reprodukcyjnego (P>0,05).
W całym doświadczeniu urodziło się 265 prosiąt o średniej masie ciała około 1,46 kg
(Tabela 39). Średnia ilość prosiąt w miocie wynosiła od 10,8 w grupie 1 i 3 do 11,5 w grupie 4.
W czasie odchowu przy lochach prosięta we wszystkich grupach przyrastały średnio nieco
ponad 200 g dziennie, a po odsadzeniu od loch 341-351 g/dz, przy czym nie stwierdzono
istotnych statystycznie różnic mędzy grupami. Masa ciała prosiąt z grup doświadczalnych w
dniu odsadzenia (28 dz.) oraz zakończenia doświadczenia (84 dz.) nie różniła się od masy ciała
prosiąt z grupy kontrolnej. We wszystkich grupach zwierzęta padłe, przygniecione i
wybrakowane stanowiły od 9,2 do 12,1% ilości prosiąt urodzonych. Nie stwierdzono także
różnic między grupą kontrolną a grupami doświadczalnymi analizując zużycie paszy przez
prosięta. Podsumowując, uzyskane wyniki wskazują na brak wpływu udziału badanych
materiałów paszowych GM w diecie na wskaźniki reprodukcyjne loch i parametry
odchowu prosiąt.
42
Tabela 38.
Zmiany masy ciała loch oraz pobranie paszy w czasie cyklu reprodukcyjnego.
Grupa doświadczalna*
2
3
1
Średnia masa ciała loch (kg)
krycie
225,5
206,5
100 dzień ciąży
266,3
248,0
wyproszenie
256,8
238,7
odsadzenie
245,2
228,8
Zmiany masy ciała loch w poszczególnych okresach cyklu (kg)
od krycia do 100 dnia ciąży
40,8
41,5
od 100 dnia ciąży do wyproszenia
9,5
9,3
od krycia do wyproszenia
31,3
32,2
laktacja
11,6
9,9
cały cykl
19,7
22,3
Pobranie paszy (kg)
od krycia do 100 dnia ciąży
244,2
242,8
od 100 dnia ciąży do wyproszenia
63,6
62,4
od krycia do wyproszenia
307,8
305,2
laktacja
155,2
155,2
cały cykl
463,0
460,4
SEM
4
Poziom
P
215,7
255,8
246,2
234,8
225,0
267,7
259,3
246,7
0,602
0,473
0,465
0,550
5,514
4,938
5,061
4,915
40,1
9,6
30,5
11,4
19,1
42,7
8,4
34,3
12,6
21,7
0,965
0,931
0,928
0,832
0,939
1,671
0,739
2,005
1,037
1,967
243,8
57,2
301,0
156,8
457,8
241,7
63,6
305,3
160,0
465,3
0,836
0,248
0,621
0,972
0,925
0,991
1,295
1,800
3,802
3,918
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 39.
Wyniki odchowu prosiąt.
Grupa doświadczalna*
1
2
3
Ilość prosiąt urodzonych (szt.)
65
66
65
Ilość prosiąt odsadzonych (szt.)
58
58
59
Ilość prosiąt urodzonych w miocie (szt.)
10,8
11,0
10,8
Ilość prosiąt odsadzonych w miocie (szt.)
9,7
9,7
9,8
Odsetek padnięć (%)
10,8
12,1
9,2
Średnia indywidualna masa ciała prosiąt w kolejnych dniach odchowu (kg)
1 dzień życia
1,47
1,48
1,50
28 dzień życia
7,16
7,18
7,32
84 dzień życia
26,27
26,47
27,04
Średnie dzienne przyrosty masy ciała prosiąt w wybranym okresie odchowu (g)
1 – 28 dzień życia
211
211
221
28 – 84 dzień życia
341
344
349
1 – 84 dzień życia
299
301
308
Zużycie paszy na przyrost 1 kg masy ciała (kg)
1 – 28 dzień życia
0,12
0,12
0,11
28 – 84 dzień życia
2,13
2,08
2,06
1 – 84 dzień życia
1,64
1,62
1,59
Poziom
P
-
SEM
4
69
61
11,5
10,2
11,6
1,40
7,20
26,85
0,265
0,686
0,853
0,020
0,099
0,341
215
351
307
0,675
0,911
0,834
3,426
5,329
4,045
0,12
2,01
1,57
0,857
0,911
0,912
0,002
0,054
0,033
-
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
43
Badania wykonane w Państwowym Instytucie Weterynaryjnym - PIB
Ocena wskaźników hematologicznych krwi i komórkowej odpowiedzi immunologicznej
W ramach realizacji badań określono wpływ stosowanych pasz GM na efektywność
działania układu immunologicznego loch. Krew do badań immunologicznych wybranych
wskaźników nieswoistej odporności komórkowej pobierano dwukrotnie, tj. przed i po
skarmianiu badanych materiałów paszowych GM. W ramach przeprowadzonych badań
immunologicznych oceniano skład procentowy poszczególnych subpopulacji limfocytów krwi
obwodowej z użyciem cytometrii przepływowej i specyficznego panelu przeciwciał
monoklonalnych umożliwiających rozpoznawanie komórek CD3+ (limf. T), CD4+ (limf. Th;
pomocnicze) i CD8+ (limfocyty Tc/s; cytotoksyczno-supresorowe). Wykonano również
oznaczenie ogólnych wskaźników statusu zdrowotnego, tj. wskaźników erytrocytarnych
(liczba czerwonych krwinek – RBC, ilość hemoglobiny – HGB, hematokryt – HCT, średnia
objętość erytrocytu – MCV, średni ciężar i stężenie hemoglobiny w jednej krwince – MCH i
MCHC) i trombocytarnych (liczba płytek krwi – PLT i ich średnia objętość – MVP) oraz
wybranych parametrów odporności komórkowej, takich jak liczba białych ciałek krwi (WBC) i
leukogram, tj. jakościowy skład poszczególnych subpopulacji leukocytów z podziałem na
granulocyty obojętnochłonne – PMNL, komórki średniej wielkości – MID (sumaryczna
wartość monocytów, bazofilów, eozynofilów w jednostce objętości krwi) i limfocytów – LYM..
Badane wskaźniki immunologiczne odzwierciedlają charakter zmian w ramach nieswoistej
odporności komórkowej.
W
badaniach
tzw.
ogólnych
wskaźników
zdrowia,
tj.
parametrów
układu
erytrocytarnego i trombocytarnego nie obserwowano różnic pomiędzy grupą kontrolną, w
której zwierzęta żywione były dietą bez materiałów paszowych GM oraz grupami
doświadczalnymi (Tabela 40 i 41). Podobnie, brak efektu stosowania pasz GM stwierdzono też
w odniesieniu do pozostałych ocenianych parametrów nieswoistej odporności komórkowej, tj.
ogólnej liczby leukocytów we krwi (WBC), leukogramu (LYM, PMNL, MID) i
immunofenotypu limfocytów z podziałem na komórki CD3+, CD4+ i CD8+ (Tabela 42 i 43).
Tabela. 40. Średnie wartości wskaźników erytrocytarnych krwi obwodowej u loch.
RBC (x 1012/l)
MCV (fl)
0
1
1
6,13±0,49
6,41±0,36
Grupa doświadczalna*
2
3
5,91±0,60
5,67±0,4
5,69±0,31
5,89±0,5
4
7,05±0,2
6,96±0,4
0
1
0
61,4±1,53
63,9±1,81
0,65±0,10
55,9±1,18
49,9±0,89
0,67±0,10
70,1±3,01
72,0±10,1
0,64±0,21
44
60,2±1,41
64,7±2,12
0,63±0,22
HCT (l/l)
1
0
1
0
1
0
1
HGB (mmol/l)
MCH (fmol)
MCHC (mmol/l)
0,66±0,14
10,01±1,59
10,65±1,08
1,8±0,01
1,7±0,04
28,2±0,2
29,1±0,3
0,65±0,32
10,11±0,51
9,50±1,10
1,20±0,04
1,75±0,07
26,5±0,27
28,5±0,21
0,60±0,24
9,14±1,16
9,63±2,10
1,15±0,07
1,35±0,04
28,7±0,37
27,5±0,71
0,62±0,14
11,9±2,06
11,2±1,46
1,91±0,04
2,10±0,09
29,2±0,14
24,5±0,17
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
0 – wartości wyjściowe przed rozpoczęciem doświadczenia; 1 – wartości po żywieniu paszami GM
Tabela 41. Średnie wartości wskaźników trombocytarnych krwi obwodowej u loch.
PLT (109/L)
MPV (fl)
0
1
230,2±110,0
Grupa doświadczalna *
2
3
321,9±122,7
218,0±58,9
4
299,5±54,09
1
299,8±77,8
310,5±66,96
230,0±78,33
305,5±94,11
0
5,6±0,31
5,7±0,45
5,6±0,17
5,6±0,11
1
5,5±0,19
5,9±0,44
5,7±0,33
5,8±0,45
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
0 – wartości wyjściowe przed rozpoczęciem doświadczenia, 1 – wartości po żywieniu paszami GM
Tabela 42.
Średnie wartości wskaźników leukocytarnych krwi obwodowej u loch.
1
11,59
±5,10
Grupa doświadczalna *
2
3
12,5
9,4
±5,13
±3,05
4
10,9
±1,22
1
10,93
±2,04
13,5
±4,15
10,5
±4,11
11,5
±2,98
0
40,51
±1,34
39,9
±5,11
37,9
±10,01
41,1
±6,15
1
42,1
±5,24
36,8
±4,12
40,0
±5,01
39,1
±4,39
0
48,1
±11,5
50,3
±5,11
49,9
±4,13
48,9
±4,35
1
49,0
±3,14
53,1
±9,01
48,9
±10,1
51,0
±5,13
0
11,39
±3,04
9,8
±2,88
12,2
±1,02
10,0
±1,91
1
8,9
±2,07
10,1
±2,44
11,1
±3,4
9,9
±3,11
0
9
WBC (x 10 /L)
PMNL (%)
LYM (%)
MID (%)
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
0 – wartości wyjściowe przed rozpoczęciem doświadczenia, 1 – wartości po żywieniu paszami GM
45
Tabela 43.
Zmiany w subpopulacjach limfocytów krwi obwodowej u loch.
1
48,23
±3,1
Grupa doświadczalna *
2
3
53,11
49,91
±4,10
±5,14
4
47,97
±3,91
1
50,13
±2,96
51,95
±4,12
52,58
±3,65
49,97
±4,61
0
27,58
±4,44
31,07
±2,24
29,14
±2,96
26,92
±3,3
1
28,22
±2,20
29,74
±3,61
30,18
±2,81
29,56
±4,2
0
29,31
±3,06
33,47
±4,63
33,10
±5,12
27,91
±3,11
1
35,11
±4,01
36,10
±4,08
35,81
±3,55
30,21
±5,22
0
CD3+ (%)
+
CD4 (%)
+
CD8 (%)
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
0 – wartości wyjściowe przed rozpoczęciem doświadczenia, 1 – wartości po żywieniu paszami GM
Ocena humoralnej odpowiedzi immunologicznej
W trakcie realizacji doświadczenia przeprowadzono także ocenę odpowiedzi
immunologicznej po szczepieniach loch biopreparatami atenuowanymi lub inaktywowanymi.
W każdej z grup po 4 wybrane losowo lochy zostały zaszczepione wybraną szczepionką, a 2
pozostawały nieszczepione (kontrolne). Układ szczepień: grupy I i II - zaszczepiono
domięśniowo preparatem Respisure (szczepionka inaktywowana przeciw mykoplazmowemu
zapaleniu płuc) dwukrotnie w odstępie 3 tygodni; grupy III i IV - zaszczepiono preparatem
Akipor 6.3 (przeciw chorobie Aujeszkiego) w analogicznych terminach. Krew do analizy
poziomu przeciwciał pobierano od wszystkich loch czterokrotnie, w odstępach około 3tygodniowych. W celu oceny poziomu odporności biernej krew pobrano także od 5 prosiąt z
jednego miotu lochy szczepionej biopreparatem Akipor 6.3 i od 5 prosiąt z jednego miotu lochy
immunizowanej biopreparatem Respisure. Dla porównania pobrano krew także od prosiąt z
miotów loch kontrolnych z odpowiednich grup. Próbki krwi w kierunku obecności przeciwciał
dla wirusa PRRS i Mycoplasma hyopneumoniae przebadano testami ELISA. Do badań
wykorzystano zestawy firmy IDEXX w przypadku Mycoplasma hyopneumoniae oraz zestaw
własny w przypadku badań w kierunku PRRS. W odniesieniu do PRRS poziom swoistych
przeciwciał humoralnych określono w zakresie od 0 (brak przeciwciał) do 6 (maksymalny
poziom przeciwciał).
W odniesieniu do Mycoplasma hyopneumoniae poziom przeciwciał
46
humoralnych ustalono na podstawie pomiaru OD badanych surowic. Surowice krwi
przeznaczone do badania w kierunku poziomu obecności przeciwciał dla wirusa chA
przebadano odczynem seroneutralizacji. Ustalono miana ND50 poszczególnych badanych
surowic.
Wyniki badań serologicznych loch w kierunku Mycoplasma hyopneumoniae i choroby
Aujeszkiego przedstawiono w Tabeli 44. Analiza uzyskanych danych wskazuje, że szczepienia
biopreparatem
Akipor
6.3
przeciw
chorobie
Aujeszkiego
we
wszystkich
grupach
doświadczalnych uwidoczniły się w postaci wzrostu miana swoistych przeciwciał humoralnych
dla wirusa chA. W grupach loch immunizowanych miana przeciwciał były po drugim
szczepieniu wyraźnie wyższe niż po pierwszym. W grupach kontrolnych nie wykazano
obecności przeciwciał. Analiza rezultatów szczepień loch przeciwko zakażeniom wirusem chA
dowodzi efektywności szczepień. Analiza wyników w grupach loch szczepionych szczepionką
Respisure przeciwko Mycoplasma hyopneumoniae uwidoczniła, że miana swoistych
przeciwciał w grupie świń doświadczalnych żywionych paszą GM nie różniły się istotnie od
tych jakie zarejestrowano w grupie zwierząt kontrolnych (Tabela 44). Przyczyną spadku miana
swoistych przeciwciał powstałych po zakażeniu świń, a później szczepionych MPS mogła być
interferencja przeciwciał powstałych po naturalnym zakażeniu oraz antygenu zawartego w
szczepionce. W przypadku MPS, odpowiedź immunologiczna na szczepienie, w postaci
wzrostu poziomu przeciwciał wykrywanych za pomocą zastosowanego w badaniach testu
diagnostycznego (ELISA), może w warunkach standardowej fermy zachodzić w różnym czasie
po szczepieniu, na co wpływ mogą mieć między innymi zakaźne i niezakaźne czynniki
immunomodulujące a także cechy osobnicze zwierząt.
W Tabeli 45 wyraźnie uwidocznił się wpływ szczepień loch na poziom odporności
biernej ich potomstwa. Wykazano obecność statystycznie istotnych różnic między średnim
poziomem
miana
przeciwciał
u
osesków
pochodzących
od
loch
szczepionych
i
nieszczepionych. Na podstawie otrzymanych wyników stwierdzono brak potwierdzonych
różnic między średnim poziomem miana przeciwciał w grupach świń żywionych mieszanką
paszową zawierającą materiały GM oraz dietą kontrolną.
Podsumowując, analiza wykonanych oznaczeń hematologiczno-immunologicznych
krwi loch potwierdza wyniki uzyskane w tym zakresie u tuczników, wskazując na brak
wpływu badanych materiałów paszowych GM na przebieg komórkowych i humoralnych
procesów immunologicznych.
47
Tabela 44.
Wyniki badań serologicznych krwi loch w kierunku obecności i poziomu przeciwciał
przeciwko mykoplazmowemu zapaleniu płuc (MPS) oraz chorobie Aujeszkyego
(średnie wartości przeciwciał).
Grupa doświadczalna *
Lochy kontrolne
Lochy
szczepione
(Aujeszky)
Lochy kontrolne
4
Lochy
szczepione
(Aujeszky)
Lochy
szczepione
(MPS)
Lochy
szczepione
(MPS)
Pobranie przed szczepieniem
3
Lochy kontrolne
2
Lochy kontrolne
1
0,720**
0,809
0,875
0,605
< 0,3
< 0,3
< 0,3
< 0,3
Pobranie po 1 szczepieniu
0,058
0,478
0,056
0,301
0,42
< 0,3
< 0,3
< 0,3
Pobranie po 2 szczepieniu
0,060
0,534
0,184
0,351
1,42
< 0,3
1,17
< 0,3
Ostatnie pobranie
0,133
0,806
0,070
0,475
0,99
< 0,3
0,80
< 0,3
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
**/ log10 ND50
Poziom humoralnych przeciwciał biernych u prosiąt pochodzących od loch szczepionych
przeciwko mykoplazmowemu zapaleniu płuc (MPS) oraz chorobie Aujeszkiego.
Grupa doświadczalna *
2
3
Lochy
kontrolne
Lochy
szczepione
(MPS)
Lochy
kontrolne
Lochy
szczepione
(Aujeszky)
Lochy
kontrolne
Lochy
szczepione
(Aujeszky)
Poziom przeciwciał we
krwi prosiąt
4
Lochy
szczepione
(MPS)
1
0,905**
0,269
0,848
0,140
0,490
<0,3
0,330
Lochy
kontrolne
Tabela 45.
<0,3
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
**/ log10 ND50
Analiza obecności transgenicznego DNA we krwi
W czasie doświadczenia od wszystkich loch pobrano próbki krwi w celu analizy
obecności
transgenicznego
DNA,
pochodzącego
z
badanych
roślin
genetycznie
zmodyfikowanych. Oznaczenia wykonane w Krajowym Laboratorium Pasz Instytutu
Zootechniki PIB (Pracownia w Szczecinie), przeprowadzone we krwi pełnej pobranej od loch
doświadczalnych, nie wykazały obecności sekwencji DNA specyficznych dla genetycznie
zmodyfikowanej soi i kukurydzy (Tabela 46).
48
Tabela 46. Wyniki jakościowej analizy DNA w kierunku obecności specyficznych dla GMO
transgenów we krwi loch.
1
RR
Krew
-
MON
810
-
Grupa doświadczalna *
2
3
RR
MON
RR
MON
810
810
n/a
n/a
-
4
RR
-
MON
810
-
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
(-) nie stwierdzono obecności sekwencji specyficznej dla soi RR lub kukurydzy MON810, przy granicy wykrywalności metody 0,1%.
(n/a) – nie analizowano
BADANIA NA CIELĘTACH
Doświadczenie żywieniowe na cielętach-buhajkach rasy czarno-białej w wieku od 7-10
do 90 dnia życia wykonano w Zakładzie Doświadczalnym Instytutu Zootechniki PIB Rudawa
Sp. z o.o, w oborze doświadczalnej w Aleksandrowicach. Cielęta utrzymywano w
indywidualnych klatkach firmy Alfa-Laval wyścielanych słomą i wyposażonych w poidła,
żłoby i obręcze na wiadra. Do 56 dnia życia cielęta otrzymywały z wiader ze smoczkiem paszę
płynną w postaci preparatu mleko-zastępczego, a od początku doświadczenia miały stały dostęp
do mieszanki paszowej. Zwierzęta żywiono do woli mieszanką treściwą zbilansowaną pod
względem energetycznym i białkowym według norm IZ-INRA (2001).
Utworzono 4 grupy doświadczalne, po 10 zwierząt, żywionych mieszankami
paszowymi, w których głównym źródłem energii i białka były materiały zmodyfikowane lub
niezmodyfikowane genetycznie:
Grupa 1 – śruta z ziarna kukurydzy niemodyfikowanej + poekstrakcyjna śruta sojowa
niemodyfikowana.
Grupa 2 – śruta z ziarna kukurydzy niemodyfikowanej + poekstrakcyjna śruta sojowa GM.
Grupa 3 – śruta z ziarna kukurydzy GM + poekstrakcyjna śruta sojowa niemodyfikowana.
Grupa 4 – śruta z ziarna kukurydzy GM + poekstrakcyjna śruta sojowa GM.
Określano pobranie i wykorzystanie paszy i dzienne przyrosty masy ciała w kolejnych
okresach odchowu oraz zawartość suchej masy, białka i tłuszczu i kwasów tłuszczowych w
tłuszczu śródmięśniowym Musculus Thoracis. Po zakończeniu doświadczenia 5 cieląt z każdej
grupy poddano ubojowi w celu oznaczenia transferu transgenicznego DNA w przewodzie
pokarmowym oraz jego retencję w tkankach i mięśniach (laboratorium Państwowego Instytutu
Weterynaryjnego - PIB, Krajowe Laboratorium Pasz Instytutu Zootechniki PIB). Dodatkowo
badano wpływ materiałów paszowych GM na obraz histopatologiczny narządów wewnętrznych
oraz status immunologiczny cieląt (Państwowy Instytut Weterynaryjny - PIB).
49
Wyniki
Badania prowadzone w Instytucie Zootechniki PIB
Wskaźniki wzrostowe i skład chemiczny cielęciny
Analiza statystyczna wyników nie wykazała różnic pomiędzy grupą kontrolną (dieta bez
materiałów paszowych GM) a grupami doświadczalnymi w pobraniu mieszanki treściwej i
suchej masy, końcowej masie ciała, średnich dziennych przyrostach masy ciała cieląt oraz w
zużyciu paszy na 1 kg przyrostu (Tabela 47, 48 i 49). Podobnie nie odnotowano różnic między
grupami cieląt, analizując dane doświadczalne dotyczące składu chemicznego mięsa (zawartość
suchej masy, białka ogólnego, tłuszczu i popiołu) oraz profil kwasów tłuszczowych lipidów
mięsa (50 i 51). Podsumowując, można stwierdzić, że stosowane materiały paszowe GM
nie wpływają na wskaźniki wzrostowe cieląt oraz skład chemiczny cielęciny.
Tabela 47.
Masa ciała i dzienne przyrosty masy ciała cieląt.
Grupa doświadczalna*
SEM
1
2
3
4
45,25
42,75
42,85
47,22
0,84
w 56 dniu życia
73,75ab
68,95a
70,05a
76,00b
0,96
końcowa (w 90 dniu życia)
113,00
109,15
107,65
119,50
1,85
Dzienne przyrosty masy ciała
od 10 do 56 dnia życia, g
od 56 do 90 dnia życia, g
605,70
1154,40
558,80
1182,50
577,30
1105,80
626,00
1279,44
16,42
39,35
od 10 do 90 dnia życia, g
837,40
820,10
799,7
902,77
21,72
Masa ciała, kg
początkowa
a, b – P ≤ 0.05
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 48.
Pobranie paszy i składników pokarmowych w poszczególnych okresach odchowu cieląt.
Grupa doświadczalna*
1
Przed odsadzeniem (od 10 do 56 dnia życia):
preparat mlekozastępczy, kg/d
0,83a
mieszanka treściwa kg/dzień
0,39
sucha masa, kg/dzień
1,16
białko ogólne, g/dzień
234,60
BTJN**, g/dzień
167,59
BTJE***, g/dzień
255,03
JPM****/dzień
1,76
50
2
3
4
0,83a
0,33
1,10
220,27
155,90
246,27
1,70
0,82a
0,35
1,10
223,61
159,29
245,81
1,69
0,87b
0,38
1,19
240,16
168,74
264,87
1,83
SEM
0,007
0,018
0,019
3,951
2,876
3,118
0,023
Po odsadzeniu (od 56-90 dnia życia) :
mieszanka treściwa, kg/dzień
sucha masa, kg/dzień
białko ogólne, g/dzień
BTJN**, g/dzień
BTJE***, g/dzień
JPM****/dzień
3,32
2,95
614,52
455,07
395,28
3,19
3,01
2,67
542,22
406,67
358,47
2,89
3,32
2,94
614,15
454,80
395,05
3,19
3,72
3,30
673,25
498,43
442,63
3,57
0,095
0,085
17,519
12,867
11,365
0,092
Cały okres doświadczenia (od 1-90 dnia życia):
mieszanka treściwa, kg/dzień
1,63
sucha masa, kg/dzień
2,25ab
białko ogólne, g/dzień
462,42ab
BTJN**, g/dzień
361,81ab
BTJE***, g/dzień
401,61ab
JPM****/dzień
2,95ab
1,46
2,10a
423,38a
351,00a
379,41a
2,78a
1,59
2,20ab
455,60ab
387,77b
392,99a
2,88a
1,80
2,44b
496,61b
385,51b
432,06b
3,17b
0,047
0,040
9,229
5,261
6,416
0,050
a, b – P ≤ 0.05
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
**/BTJN - białko paszy nie ulegające rozkładowi w żwaczu i białko mikoorganizmów żwaczowych trawione w jelicie cienkim, obliczone
na podstawie dostępnego azotu w żwaczu
***/BTJE - białko paszy nie ulegające rozkładowi w żwaczu i białko mikoorganizmów żwaczowych trawione w jelicie cienkim,
obliczone na podstawie dostępnej energii w żwaczu
****/JPM - jednostka paszowa produkcji mleka = 1700 kcal
Tabela 49.
Zużycie paszy i wykorzystanie składników pokarmowych na 1 kg przyrostu masy ciała
cieląt w całym okresie doświadczanym.
Grupa doświadczalna*
Mieszanka treściwa, kg
Sucha masa, kg
Białko ogólne, g
BTJ, g
JPM
SEM
1
2
3
4
1,93
2,72
559,31
489,56
3,58
1,77
2,57
517,45
464,48
3,40
2,01
2,79
577,59
498,95
3,65
2,00
2,73
555,71
485,03
3,56
0,065
0,087
17,644
17,526
0,123
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 50.
Skład chemiczny mięsa (% w SM).
Grupa doświadczalna*
SEM
1
2
3
4
Sucha masa
22,790
22,192
22,146
22,422
0,110
Białko ogólne
92,424
91,470
91,798
90,494
0,359
Tłuszcz
3,920
3,876
3,994
3,540
0,143
Popiół
4,896
4,736
4,842
4,682
0,059
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
51
Tabela 51. Profil kwasów tłuszczowych w tłuszczu mięsa cieląt (Musculus thoracis ), g/100g wszystkich
oznaczonych kwasów tłuszczowych.
Kwasy tłuszczowe
1
Grupa doświadczalna*
2
3
4
SEM
C 8:0
C 10:0
C 12:0
C 14:0
C 16:0
C 16:1
C 18:0
C 18:1
C 18:2, n-6
C 18:3- γ
C 18:3- α
0,036
0,660
1,798
23,901
1,261
11,298
27,668
23,329
0,0378
0,4333
0,033
0,598
1,856
21,833
1,096
12,102
25,043
25,279
0,0884
0,664
0,165
0,090
0,576
1,490
20,941
0,846
12,011
22,976
27,674
0,118
0,541
0,018
0,077
0,609
1,540
20,951
0,850
12,524
21,822
28,291
0,092
0,631
0,033
0,028
0,141
0,253
1,031
0,085
0,338
1,446
1,176
0,020
0,045
C 20:0
CLA c-9-t 11
0,263
0,338
0,207
0,507
0,171
0,386
0,187
0,379
0,037
0,049
CLA t10-c12
CLA c-9-c 11
CLA t9-t11
C 22:0
C 20:4
C 22:1
C 20:5, n-3, EPA
C 22:6, n-3, DHA
SFA
UFA
MUFA
0,235
0,071
0,086
0,014
7,760
0,357
0,218
0,231
37,971
62,029
29,287
0,194
0,163
0,109
0,176
9,099
0,304
0,290
0,358
36,805
63,195
26,443
0,208
0,126
0,178
0,257
10,017
0,375
0,372
0,480
35,702
64,298
24,196
0,149
0,097
0,096
0,258
10,163
0,389
0,319
0,553
36,166
63,834
23,061
0,030
0,031
0,019
0,058
0,649
0,042
0,056
0,075
1,436
1,433
1,483
PUFA
n-6
32,742
31,127
36,753
34,467
40,102
37,809
40,772
38,546
1,869
1,754
n-3
DFA
OFA
UFA/SFA
0,884
73,327
26,673
1,699
1,312
75,297
24,703
1,790
1,393
76,309
23,690
1,855
1,503
76,357
23,643
1,845
0,145
1,330
1,335
0,094
DFA/OFA
MUFA/SFA
2,882
0,800
3,264
0,738
3,444
0,697
3,417
0,656
0,198
0,050
PUFA/SFA
n-6/n-3
0,899
47,839
1,052
29,061
1,158
37,881
1,188
30,835
0,081
4,905
CLA
0,731
0,973
0,899
0,723
0,065
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
52
Badania wykonane w Państwowym Instytucie Weterynaryjnym - PIB
Ocena wskaźników hematologicznych krwi i efektywności odpowiedzi immunologicznej
W doświadczeniu wykonano ocenę statusu immunologicznego cieląt żywionych paszą z
udziałem materiałów GM. Badania wykonano na 20 cielętach (5 z każdej grupy) biorących
udział w doświadczeniu żywieniowym. Z żyły szyjnej zewnętrznej pobrano krew na EDTAK 2
oraz do probówek bez dodatku antykoagulantu do badań w kierunku oceny:
a) tzw. ogólnych wskaźników zdrowia, tj. parametrów układu czerwonokrwinkowego i płytek
krwi (RBC – liczba czerwonych krwinek, HGB - ilość hemoglobiny, HCT - hematokryt, MCV
- średnia objętość erytrocytu, MCH i MCHC - średni ciężar i stężenie hemoglobiny w jednej
krwince, PTL - liczba płytek krwi, MVP – średnia objętość płytek krwi).
b) komórkowej odpowiedzi immunologicznej z wykorzystaniem cytometrii przepływowej
(FCM) na podstawie immunofenotypowej analizy subpopulacji leukocytów krwi obwodowej
(WBC - liczba białych ciałek krwi, PMNL – liczba granulocytów obojętnochłonnych, MID –
liczba komórek średniej wielkości, tj. sumy monocytów, bazofilów i eozynofilów, LYM –
liczba limfocytów, CD2+ - limfocyty T, CD4+ - limfocyty T pomocnicze, Th, CD8+ limfocyty cytotoksyczno-supresorowe, Tc/s i WC4 - limfocyty B).
c) humoralnej odpowiedzi immunologicznej poprzez analizę miana swoistych przeciwciał
poszczepiennych anty – BRSV (wirus syncytialnego układu oddechowego bydła), PI3V
(parainfluenza typu 3) i BVDV (wirus biegunki bydła i choroby błon śluzowych typu 1).
Badania odporności humoralnej cieląt poprzedzone były dwukrotnym szczepieniem
poliwalentną szczepionką Rispoval 3 (Pfizer) - pierwsze szczepienie w wieku 14 dni, kolejne po 2-3 tygodniach. Szczepionka ta zawierała żywe, atenuowane szczepy wirusa syncytialnego
układu oddechowego bydła (BRSV) i parainfluenzy typu 3 (PI3V) oraz inaktywowany wirus
biegunki bydła i choroby błon śluzowych typu 1 (BVDV 1). Szczepionkę podawano
domięśniowo (i.m.) w ilości 4 ml po uprzednim rozpuszczeniu liofilizatu, zwierającego żywe
komponenty wirusa BRS i PI3, płynnym rozcieńczalnikiem stanowiącym nośnik dla wirusa
BVD. Dodatkowo przed samym szczepieniem od dwóch wybranych losowo zwierząt w każdej
grupie pobrano krew do w/w badań immunologicznych w celu ustalenia wyjściowego poziomu
odporności, zarówno komórkowej jak i humoralnej (grupę tą jako zbiorczą n-8 oznaczono
symbolem K0 i traktowano jako kontrolę ujemną).
Badania ogólnych wskaźników zdrowia, tj. erytrocytarnych (liczba czerwonych krwinek
– RBC, ilość hemoglobiny – HGB, hematokryt – HCT, średnia objętość erytrocytu – MCV,
53
średni ciężar i stężenie hemoglobiny w jednej krwince – MCH i MCHC) i trombocytarnych
(liczba płytek krwi – PLT i ich średnia objętość – MVP) oraz wybranych parametrów
odporności komórkowej, takich jak liczba białych ciałek krwi (WBC) i leukogram, tj.
jakościowy skład poszczególnych subpopulacji leukocytów z podziałem na granulocyty
obojętnochłonne – PMNL, komórki średniej wielkości – MID (sumaryczna wartość
monocytów, bazofilów, eozynofilów w jednostce objętości krwi) i limfocytów – LYM,
wykonano przy użyciu analizatora typu AC 920 AutoCounter Swelab Instrument AB.
Immunofenotypowanie limfocytów, tj. szczegółowa ocena poszczególnych subpopulacji
obwodowych limfocytów krwi cieląt z uwzględnieniem frakcji komórek typu CD2+ (limfocyty
T), CD4+ (Limfocyty T pomocnicze – Th), CD8+ (limfocyty cytotoksyczno-supresorowe –
Tc/s) i WC4 (limfocyty B) przeprowadzona została z wykorzystaniem cytometru
przepływowego Epics 4C XL f-my Beckman Coulter.
W celu oceny swoistej odporności humoralnej analizowano efekty immunizacji cieląt
szczepionką Rispoval 3 badając poziom odpowiedzi poszczepiennej w odniesieniu do trzech
antygenów szczepionkowych, tj. BRSV, PI3V i BVDV testem Elisa o nazwie Pentakit
produkcji BIO-X-Diagnostics (Belgia). Test ten umożliwia badanie obecności przeciwciał
przeciwko pięciu różnym wirusom występującym u bydła (BHV1, BVDV, BRSV, PI3V,
Adenowirus 3). W obecnych badaniach skoncentrowano się przede wszystkim na analizie
rezultatów swoistej odpowiedzi humoralnej w odniesieniu do trzech wirusów BVDV, BRSV,
PI3V, tj. tych których antygeny zawierała stosowana szczepionka. Pozostałe antygeny
wirusowe były również badane, jednak ich znaczenie nie było w realizowanym doświadczeniu
tak istotne.
W badaniach wskaźników erytrocytarnych i trombocytarnych (Tabele 52 i 53) nie
stwierdzono różnic pomiędzy grupami cieląt, a ich średnie wartości utrzymywały się na
poziomie uznanym za fizjologiczny dla tego gatunku zwierząt i badanej grupy wiekowej.
Nieznacznie wyższe średnie wartości zaobserwowano w odniesieniu do liczby erytrocytów
(RBC) i hemoglobiny (HGB) we krwi cieląt grupy 2 w porównaniu do pozostałych zwierząt,
jednak nie były to różnice statystycznie istotne. Z kolei nieco niższe wartości notowano w tej
grupie cieląt, a także w grupie 1 w zakresie ogólnej liczby płytek krwi (PLT) we krwi
obwodowej tych zwierząt, przy czym i w tym wypadku nie były to różnice istotne statystycznie
(Tabela 53).
54
Tabela 52. Średnie wartości wskaźników układu erytrocytarnego cieląt.
RBC
HCT
MCV
HGB
MCH
MCHC
(l/l)
(fl)
(mmol/l
(fmol)
(mmol/l)
12
(10 /l
K0
6,28±1,07
0,21±0,05
33,43±2,15
7,40±1,01
1,13±0,08
35,43±4,41
Grupa 1
7,87±1,24
0,28±0,06
35,25±2,63
8,30±3,01
0,98±0,25
28,78±6,69
Grupa 2
8,54±0,66
0,31±0,03
36,50±2,08
10,23±1,02
1,13±0,05
32,83±0,92
Grupa 3
7,55±1,26
0,26±0,06
34,80±1,79
9,10±1,34
1,16±0,05
35,02±2,83
Grupa 4
7,83±1,63
0,27±0,07
42,75±17,17
7,95±2,07
1,13±0,10
34,93±3,73
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Tabela 53. Średnie wartości wskaźników układu trombocytarnego cieląt.
PLT
MPV
9
(10 /L)
(fl)
K0
623,29±237,06
6,49±0,50
Grupa 1
417,50±221,75
6,48±0,15
Grupa 2
445,75±251,39
6,78±0,60
Grupa 3
629,00±201,60
6,78±0,55
Grupa 4
504,00±139,30
8,20±3,00
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Podobnie kształtowały się u cieląt grupy 2 niektóre parametry odporności komórkowej
(Tabela 54). Ogólna liczba leukocytów we krwi obwodowej tych zwierząt była bowiem
podwyższona w porównaniu do pozostałych grup cieląt, w głównej mierze jako następstwo
wyższej zdecydowanie liczby granulocytów obojętnochłonnych - PMNL i tzw. komórek MID,
których dominującą frakcją są monocyty. Względne wartości tych subpopulacji leukocytów, tj.
PMNL i MID zachowywały się analogicznie i wynosiły odpowiednio: 41,75±6,24 i
8,50±1,73%. W tym miejscu należy jednak pokreślić, że ogólna liczba leukocytów oraz
liczebność omawianych wyżej subpopulacji komórkowych była wyraźnie wyższa we
wszystkich 4 grupach doświadczalnych, jeśli porówna się ich wartości z tymi notowanymi w
kontroli ujemnej (K0), tj. uzyskanymi przed immunizacją cieląt szczepionką Rispoval 3.
Zmiany te były najwyraźniej skutkiem stymulującego wpływu szczepienia na kształtowanie się
poziomu analizowanych komórek.
55
Tabela 54. Wybrane wskaźniki odporności komórkowej cieląt.
WBC
LYM
MID
PMNL
LYM
MID
PMNL
9
9
9
9
(%)
(%)
(%)
(10 /L)
(10 /L)
(10 /L)
(10 /L)
K0
0,64±2,44
7,10±2,06
0,73±0,25 2, 81±0,89
67,43±9,02
6,29±1,60
26,29±7,59
Grupa 1
2,23±2,08
7,73±2,44
0,93±0,28 3, 58±1,48
64,00±13,7
7,00±2,00
29,00±11,8
Grupa 2
3,98±5,12
6,60±1,78
1,23±0,46 6, 15±3,05
49,75±5,56
8,50±1,73
41,75±6,24
Grupa 3
2,46±4,24
6,24±1,84
0,94±0,67 5, 28±2,00
52,00±5,10
6,60±3,05
41,40±5,13
Grupa 4
1,43±1,76
7,25±1,30
0,73±0,15 3, 45±0,75
53,95±32,4
9,00±6,38
37,50±7,85
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Z kolei zmiany w poszczególnych subpopulacjach limfocytów obwodowych krwi cieląt
zachowywały się różnie w zależności od badanego parametru i były one bardziej zbliżone do
siebie (Tabela 55). Najbardziej jednak widoczne różnice, choć nieistotne statystycznie,
zaobserwowano w wartościach odsetka limfocytów T i subpopulacji Tc/s. W odniesieniu do
pierwszej z tych subpopulacji najwyższe wartości odsetka limfocytów T we krwi obwodowej
cieląt zaobserwowano w grupie 3. Z kolei w subpopulacji limfocytów Tc/s zmiany te były
zbliżone do poprzednich, bowiem i w tym wypadku wyższe wartości niż w grupie Ko, 1 i 4
notowano w grupie 3. Jednak najwyższy odsetek tych komórek wykazany został w grupie 2.
Trzeba jednak pamiętać, że w badaniach tych wykazane różnice nie były istotne statystycznie, a
efekt
zwiększonej
liczebności
badanych
frakcji
komórek
we
wszystkich
grupach
doświadczalnych mógł być wynikiem niespecyficznego działania na odporność komórkową
zastosowanej immunizacji.
Tabela 55. Odsetek poszczególnych subpopulacji limfocytów (%).
Limfocyty T (CD2)
Limfocyty Th (CD4)
Limfocyty Tc/s (CD8)
Limfocyty B (W C4)
K0
57,26
31,26
16,87
20,37
Grupa 1
59,58
32,00
19,04
20,00
Grupa 2
59,54
31,28
21,05
20,80
Grupa 3
61,32
31,62
20,76
19,86
Grupa 4
59,60
32,34
19,92
20,12
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
56
Tabela 56.
Średnie wartości wybranych wskaźników aktywności fagocytarnej cieląt.
Czas
obserwacji
Czas
obserwacji
0
1
NBT (%)
Gr. 1
Gr. 2
16,060,80
18,190,68
18,990,87
19,980,73
Gr. 4
12,063,01
13,034,10
Parametry
kf (%)
Gr. 3
Gr. 4
60,774,51
63,015,01
61,092,19
65,914,91
NBT (%)
Gr. 3
Gr. 4
17,080,52
18,910,55
19,790,96
19,010,53
IF
Gr. 1
12,193,60
13,823,21
0
1
Gr. 2
13,024,33
13,992,25
Parametry
kf (%)
Gr. 1
Gr. 2
65,025,08
66,115,02
66,955,01
66,924,98
IF
Gr. 3
12,533,01
14,073,10
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
0 – wartości wyjściowe przed rozpoczęciem podawania GMO, 1 – po żywieniu
Podobny efekt immunologiczny, który należy wiązać, wyłącznie, z niespecyficzną
stymulacją poszczepienną, stwierdzono w odniesieniu do badanej aktywności fagocytarnej
PMNL (Tabela 56). Średnie wartości wskaźników tej aktywności wzrosły wyraźnie, choć
różnice te nie były istotne statystycznie i mieściły się w porównywalnym zakresie we
wszystkich grupach zwierząt wynoszącym dla IF od 13,03±4,10 do 14,07±3,10, dla kf od
61,09±2,19 do 66,95±5,01 % oraz odnośnie odsetka granulocytów NBT-dodatnich zakres jego
wartości średnich kształtował się w granicach od 18,99±0,87 do 19,98±0,73 %.
W badaniach odporności humoralnej zanotowano istotne podwyższenie średnich
wartości wskazujących na wzrost miana swoistych przeciwciał po szczepieniu we wszystkich
badanych grupach cieląt w porównaniu do kontroli ujemnej (Ko). Otrzymane rezultaty
jednoznacznie wskazują na pełną efektywność tej odpowiedzi i związaną z nią syntezą
specyficznych przeciwciał anty BRSV, PI3V i BVDV (Tabela 57). Wzrost ten był najbardziej
nasilony (P≤0,01) w odniesieniu do wirusa BRS. Dla pozostałych antygenów wirusowych, tj.
BVDV i PI3 uzyskane wartości również były wyższe niż w grupie wyjściowej (Ko), a różnice
te utrzymywały się na poziomie istotności wynoszącym P≤0,05. Obserwowane, istotne
podwyższenie wartości po immunizacji cieląt w porównaniu do wyjściowych notowanych
przed podaniem im szczepionki (Ko), dotyczyło wszystkich czterech grup zwierząt, a ich skala
była zbliżona i nie miało to, jak się wydaje bezpośredniego związku z podawaną paszą.
W odpowiedzi ostrej fazy (APR) prowadzone szczepienie pobudziło również, choć nie
istotnie zmiany w koncentracjach najbardziej czułych dla bydła białek ostrej fazy, tj. SAA i Hp,
przy czym zmiany te, z porównywalną intensywnością, były notowane we wszystkich grupach
zwierząt (Tabela 58). Maksymalne wartości dla SAA i Hp notowano po szczepieniu w grupie 1
i wynosiły one odpowiednio: 21,98±5,11 i 0,69±0,07 mg/ml. W pozostałych grupach cieląt
kształtowały się one w zakresie od 15,95±5,12 do 18,96±4,19 mg/ml w odniesieniu do SAA
oraz od 0,39±0,07 do 0,60±0,09 mg/ml dla Hp.
57
Tabela 57.
Zmiany w swoistej odpowiedzi humoralnej cieląt po immunizacji w kierunku BVDV,
BRSV, PI3V.
BHV-1 (%)
BVDV (%)
BRSV (%)
PI 3 (%)
Adeno3 (%)
K0
9,71±4,94
18,94± 9,72
2,36±1,0
43,87±16,28
14,99±7,27
Grupa 1
7,47±1,59
49,59±15,35* 6
7,21±21,14**
78,57±32,37*
20,07±11,47
Grupa 2
8,77±2,59
23,50±11,49* 8
6,92±24,58**
81,98±32,03*
3,06±1,95
Grupa 3
3,84±1,60
33,67±16,99* 8
4,82±35,25**
90,27±29,94*
12,88±6,48
Grupa 4
10,17±5,91
50,19±19,65* 8
6,25±35,07**
83,89±39,85*
24,73±10,30
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Objaśnienia: *P<0.05 i **P<0.01 - różnica statystycznie istotna w porównaniu do grupy Ko
Tabela 58.
Czas
obserwacji
0
1
Średnie stężenia wybranych białek ostrej fazy (APP) w surowicy krwi cieląt.
Parametry
SAA (mg/ml)
Gr. 1
18,163,61
21,985,11
Czas
obserwacji
0
1
Hp (mg/ml)
Gr. 2
16,782,45
18,905,13
Gr. 1
0,510,08
0,690,07
Gr. 2
0,490,10
0,580,15
Parametry
SAA (mg/ml)
Gr. 3
15,955,12
18,964,19
Hp (mg/ml)
Gr. 4
16,873,75
17,214,10
Gr. 3
0,530,11
0,600,09
Gr. 4
0,390,07
0,460,06
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
0 – wartości wyjściowe przed rozpoczęciem podawania GMO, 1 – po żywieniu
Objaśnienia: *P<0.05 i **P<0.01 - różnica statystycznie istotna w porównaniu do grupy Ko
Uogólniając wyniki badań immunologicznych na cielętach, można stwierdzić, że
brak jest wpływu u bydła stosowanych materiałów paszowych GM na procesy odporności
komórkowej, jak również swoistą odpowiedź humoralną po szczepieniu. Podawanie pasz
GM nie upośledzało reaktywności immunologicznej zwierząt na stosowane szczepienie, co
znalazło odzwierciedlenie zarówno w pełni efektywnej odpowiedzi komórkowej, jak i
humoralnej (porównywalnej z cielętami żywionymi paszami niemodyfikowanymi).
Wyniki badań histopatologicznych
W ramach doświadczenia na cielętach przeprowadzono badania histologiczne, w ramach
których od 5 zwierząt z każdej grupy pobierano wycinki wątroby, nerek, śledziony, trzustki,
dwunastnicy, jelita czczego oraz mięśni szkieletowych (mięsień smukły). Z utrwalonego w
10% roztworze formaliny materiału wykonywano skrawki parafinowe, które następnie
barwiono metodą hematoksylina-eozyna (H-E).
58
Badanie histopatologiczne wątroby, nerek, śledziony, trzustki, dwunastnicy, jelita
czczego oraz mięśni szkieletowych nie wykazało znaczących różnic pomiędzy grupą
kontrolną, w której stosowano dietę bez materiałów paszowych a grupami doświadczalnymi.
W niektórych przypadkach, niezależnie od stosowania pasz GM, obserwowano zmiany w
wątrobie i w nerkach (Tabela 59). Stwierdzone zmiany histopatologiczne występowały w
podobnym zakresie we wszystkich grupach, zarówno w grupie kontrolnej, jak i grupach
doświadczalnych, niezależnie od udziału w diecie materiałów paszowych GM. Stwierdzane
zmiany stanowią prawdopodobnie efekt intensywnego żywienia, w tym dużej zawartości białka
i energii metabolicznej w dietach, oraz szybkiego tempa metabolizmu u wysokoprodukcyjnych
zwierząt gospodarskich, natomiast piankowata struktura hepatocytów była prawdopodobnie
artefaktem charakterystycznym dla wątroby, z której wyługowany został zhydrolizowany w
czasie obróbki histologicznej glikogen. Uogólniając, należy stwierdzić, że skarmianie
mieszanek paszowych z udziałem badanych materiałów paszowych genetycznie
zmodyfikowanych nie ma wpływu na obraz histopatologiczny narządów i tkanek cieląt.
Tabela 59.
Wyniki badań histopatologicznych narządów wewnętrznych cieląt (dotyczy wszystkich
grup doświadczalnych).
Grupa doświadczalna*
1
2
Wątroba
3
4
Przekrwienie miąższu
Przekrwienie miąższu
Przekrwienie miąższu
Przekrwienie miąższu
(3/5), piankowata
(4/5), piankowata
(4/5), piankowata
(4/5), obecność
struktura hepatocytów
struktura hepatocytów
struktura hepatocytów
ogniskowych nacieków
nieznacznego stopnia
nieznacznego stopnia
nieznacznego stopnia
komórek limfoidalnych
(1/5), obecność
(2/5), obecność
(1/5), obecność
nieznacznego stopnia
ogniskowych nacieków
ogniskowych nacieków
ogniskowych nacieków
(1/5).
komórek limfoidalnych
komórek limfoidalnych
komórek limfoidalnych
nieznacznego stopnia
nieznacznego stopnia
nieznacznego stopnia
(1/5).
(1/5).
(1/5).
Nerki
Przekrwienie miąższu
Przekrwienie miąższu
Przekrwienie miąższu
Przekrwienie miąższu
nieznacznego stopnia
nieznacznego stopnia
nieznacznego stopnia
nieznacznego stopnia
(2/5),
(1/5),
(2/5),
(2/5),
nacieki komórek
nacieki komórek
nacieki komórek
nacieki komórek
limfoidalnych
limfoidalnych
limfoidalnych
limfoidalnych
nieznacznego stopnia
nieznacznego stopnia
nieznacznego stopnia
nieznacznego stopnia
(1/5).
(2/5).
(1/5).
(1/5).
Śledziona
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego
narządu.
Trzustka:
Nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego narządu. W pojedynczym
przypadku (1/20, grupa II) odnotowano zapalenie ropne.
Dwunastnica
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego
narządu.
Jelito czcze
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego
narządu.
Mięśnie
W żadnym przypadku nie stwierdzono istotnych odstępstw od prawidłowego obrazu histologicznego
szkieletowe
narządu.
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
59
Analiza obecności transgenicznego DNA w narządach, tkankach i przewodzie pokarmowym
tcieląt
W ramach wykonanego doświadczenia analizowano także pasaż transgenicznego DNA
w organizmie cieląt. W Krajowym Laboratorium Pasz Instytutu Zootechniki PIB materiał
biologiczny do badań transferu transgenicznego DNA stanowiły:
-krew,
-płuca,
-wątroba,
-śledziona,
-treść żwacza, treść dwunastnicy, treść jelita cienkiego i grubego oraz mięśnie.
W treści przedżołądka (żwacza) wszystkich zwierząt otrzymujących materiał GM
stwierdzono DNA pochodzące z soi RR i/lub kukurydzy MON (Tabela 60), co wynikało
obecności w treści żwacza niestrawionych fragmentów paszy. W treści jelita cienkiego oraz
grubego nie potwierdzono obecności DNA specyficznego dla soi RR i/lub kukurydzy MON810
oraz nie zaobserwowano niestrawionych cząstek paszy. Nie stwierdzono obecności
transgenicznego DNA w badanych narządach i tkankach, tj. w płucach, wątrobie, śledzionie i w
mięśniach. W pełnej krwi również nie stwierdzono obecności sekwencji DNA specyficznych
dla soi i kukurydzy GM.
Tabela 60.
Wyniki jakościowej analizy DNA w kierunku obecności specyficznych dla GMO
transgenów – materiał biologiczny z doświadczenia na cielętach (analizy wykonane w
KLP IZ-PIB).
Grupa doświadczalna *
2
3
1
Płuca
Wątroba
Śledziona
Tkanka
mięśniowa
Treść żwacza
Treść
dwunastnicy
Treść jelita
cienkiego
Treść jelita
grubego
Krew
4
MON810
RR
MON810
RR
MON810
RR
MON810
RR
-
-
n/a**
n/a
n/a
n/a
-
-
n/a
n/a
n/a
n/a
-
-
-
-
n/a
n/a
+
-
+
-
n/a
n/a
+
-
+
-
-
-
n/a
-
-
n/a
-
-
-
-
n/a
-
-
n/a
-
-
-
-
n/a
-
-
n/a
-
-
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
**/ nie analizowano
60
Niezależnie od badań wykonanych w Krajowym Laboratorium Pasz Instytutu
Zootechniki, analizy transgenicznego DNA w materiale biologicznym przeprowadzono także w
Państwowym Instytucie Weterynaryjnym - PIB w Puławach. Materiał genetyczny
ekstrahowano z krwi, tkanki mięśniowej, nerki, wątroby, śledziony, płuc i trzustki oraz z
wybranych odcinków przewodu pokarmowego: żołądka, dwunastnicy, jelita czczego i jelita
grubego cieląt. Wyniki analiz wykazały, że poza treścią żołądka nie stwierdzono w tkankach,
organach
oraz
pozostałych
odcinkach
przewodu
pokarmowego
transgenicznych
i
referencyjnych fragmentów DNA. Treść żołądka z grupy zwierząt żywionej mieszankami
paszowymi na bazie odmian konwencjonalnych zawierała jedynie referencyjne geny lektyny i
inwertazy. W pozostałych grupach zwierząt (2, 4) wykryte zostały elementy transgeniczne:
terminator NOS i promotor 35S charakterystyczne dla stosowanych modyfikacji. W grupie 3
zidentyfikowano promotor 35 S występujący w kukurydzy MON810 (Tabela 61).
Uogólniając uzyskane wyniki badań, można stwierdzić, że transgeniczny DNA
badanych odmian soi i kukurydzy GM jest efektywnie rozkładany w przewodzie
pokarmowym i nie zachodzi pasaż jego wykrywalnych fragmentów do krwi i narządów
organizmu.
Tabela 61.
Wyniki analizy obecności transgenicznego DNA – materiał biologiczny z doświadczenia
na cielętach (analizy wykonane w PIWet-PIB).
Grupa doświadczalna *
inwertaza
NOS
35 S
lektyna
inwertaza
NOS
35 S
lektyna
inwertaza
NOS
35 S
lektyna
inwertaza
4
lektyna
3
35 S
2
NOS
1
Krew
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Nerka
Wątroba
Śledziona
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Tkanka
mięśniowa
Płuca
Trzustka
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Zawartość
żołądka
Dwunastnica
-
-
+
+
+
+
+
+
-
+
+
+
+
+
+
+
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Jelito czcze
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
Jelito grube
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
61
Dodatkowo przeprowadzono badania mikroflory przewodu pokarmowego, które
dotyczyły przede wszystkim oceny możliwości transferu transgenicznego DNA do wybranych
grup mikroorganizmów. Analizie poddano wybrane gatunki bakterii takie jak: Escherichia coli
oraz Enterococcus faecalis i Enterococcus faecium. Bakterie izolowano z treści jelita cienkiego
i grubego. Do hodowli mikroorganizmów wykorzystywano podłoża selektywne. Dla E. coli
stosowano podłoże TBX tryptono-żółciowo glukuronidynową, a dla Enterococcus podłoże
Slanetz’a z azydkiem sodu. Do namnażania drobnoustrojów wybrano podłoże płynne BHI –
wyciąg mózgowo-sercowy. Nie wykazano obecności w bakteryjnym DNA elementów
transgenicznych: promotora 35S i terminatora NOS. Badania dotyczącego składu ilościowego
wybranych mikroorganizmów nie wykazały różnic ilościowych pomiędzy grupami żywionymi
odpowiednimi dla nich mieszankami paszowymi.
Podsumowując można stwierdzić, że stosowanie w żywieniu cieląt pasz
zawierających badane materiały paszowe GM nie wpłynęło na skład ilościowy mikroflory
przewodu pokarmowego, jak również nie stwierdzono transferu transgenicznego DNA do
bakterii zasiedlających wybrane odcinki przewodu pokarmowego.
BADANIA NA KROWACH MLECZNYCH
Doświadczenie żywieniowe na 40 krowach rasy Holsztyno-Fryzyjskiej wykonano w
Zakładzie Doświadczalnym IZ PIB Rudawa Sp. z o.o. Krowy utrzymywane były w oborze
wolno-wybiegowej i żywione grupowo TMR zbilansowanym w oparciu o normy IZ-INRA
(2001). Układ doświadczeń był następujący: 4 grupy krów, po 10 zwierząt, żywionych
mieszankami paszowymi, zawierającymi lub niezawierającymi genetycznie zmodyfikowane
materiały paszowe:
Grupa 1 – śruta kukurydziana niemodyfikowana + poekstrakcyjna śruta sojowa
niemodyfikowana.
Grupa 2 – śruta kukurydziana niemodyfikowana + poekstrakcyjna śruta sojowa GM.
Grupa 3 – śruta kukurydziana GM + poekstrakcyjna śruta sojowa niemodyfikowana.
Grupa 4 – śruta kukurydziana GM + poekstrakcyjna śruta sojowa GM.
W doświadczenia określano wpływ śruty z ziarna kukurydzy Bt oraz poekstrakcyjnej
śruty sojowej RR na produkcyjność, podstawowy skład mleka, poziom wybranych metabolitów
w surowicy krwi oraz transfer transgenicznego DNA do mleka krów.
62
Wyniki
Badania przeprowadzone w Instytucie Zootechniki PIB
Wskaźniki produkcyjne i jakość mleka
W doświadczeniu obejmującym okres od 3 tygodnia przed wycieleniem do 305 dnia
laktacji nie stwierdzono różnic pomiędzy grupą kontrolną a grupami doświadczalnymi, w
których stosowano materiały paszowe GM, w wydajności krów i składzie mleka oraz w
poziomie metabolitów (BHBA, FFA, glukoza, insulina i progesteron) w surowicy krwi (Tabela
62, Wykresy 1-3). Zestawienie uzyskanych danych eksperymentalnych wskazuje, że średnie
pobranie dawki paszowej TMR i składników pokarmowych w poszczególnych grupach w
całym okresie doświadczenia było podobne. Podsumowując, stosowanie w żywieniu krów
mlecznych badanych materiałów paszowych GM, tj. poekstrakcyjnej śruty sojowej i śruty
z ziarna kukurydzy, nie miało wpływu na wydajność mleczną, jakość mleka oraz poziom
wybranych metabolitów w surowicy krwi.
Tabela 62.
Wydajność krów oraz skład mleka.
Grupa doświadczalna*
SEM
1
2
3
4
Wydajność całkowita, kg
7916
8004
8055
8079
206,2
Wydajność dzienna, kg/dzień
27,78
26,66
26,90
27,52
0,68
Dni laktacji, dni (od 6 dnia laktacji)
285,3a
300,0 b
299,7 b
293,3 ab
1,85
tłuszcz
3,68
3,42
3,58
3,91
0,08
białko
3,26
3,24
3,20
3,42
0,05
tłuszcz/białko
1,13
1,00
1,12
1,15
0,02
laktoza
4,74
4,80
4,79
4,80
0,05
Skład mleka, kg/tydzień:
a, b – P ≤ 0.05
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
63
Wykres 1. Zawartość FFA (wolnych kwasów tłuszczowych) w surowicy krwi krów.
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Wykres 2. Zawartość kwasu β-hydroksymasłowego w surowicy krwi krów.
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
64
Wykres 3. Zawartość glukozy w surowicy krwi krów.
*/ 1 – grupa kontrolna (śruta sojowa i śruta kukurydziana konwencjonalne), 2 – śruta sojowa GM + śruta kukurydziana konwencjonalna,
3 - śruta sojowa konwencjonalna + śruta kukurydziana GM, 4 - śruta sojowa GM + śruta kukurydziana GM
Badania na krowach przetokowanych
W ramach badań na bydle wykonano także doświadczenie na trzech krowach z
przetokami do żwacza. Określono, w żwaczu, współczynniki rozkładu białka i suchej masy
badanych materiałów paszowych, tj. kukurydzy i poekstrakcyjnej śruty sojowej (Tabela 63).
Uogólniając otrzymane wyniki można stwierdzić, że rozkład białka i suchej masy w
żwaczu, w materiałach konwencjonalnych oraz genetycznie zmodyfikowanych, przebiegał
podobnie.
Tabela 63. Efektywny rozkład w żwaczu suchej masy oraz białka ogólnego pasz konwencjonalnych
oraz zmodyfikowanych genetycznie.
Wyszczególnienie
Śruta kukurydziana
GM
Śruta kukurydziana
konwencjonalna
Poekstrakcyjna śruta
sojowa GM
Poekstrakcyjna śruta
soja
konwencjonalna
a
27,7
Sucha masa
b
c
69,8
0,046
a
17,1
Białko ogólne
b
c
94,4
0,024
ERŻ
58,0
ERŻ
44,0
29,5
71,6
0,04
58,1
20,1
127,2
0,012
41,3
21,6
80,5
0,08
67,6
2,7
103,3
0,072
59,0
21,7
81,0
0,089
70,1
3,0
101,7
0,085
62,6
a- frakcja natychmiast ulegająca rozkładowi w żwaczu, b- frakcja wypływająca ze żwacza w tempie c,
c- tempo wypływu frakcji b, ERŻ- efektywny rozkład w żwaczu; k=0,06
65
Po kolejnych godzinach inkubacji badanych materiałów paszowych GM w żwaczu
oznaczono również, w Krajowym Laboratorium Pasz Instytutu Zootechniki PIB (Pracowania w
Szczecinie) obecność transgenicznego DNA. W próbkach poekstrakcyjnej śruty sojowej
genetycznie zmodyfikowanej stwierdzono u trzech krów obecność sekwencji specyficznej dla
soi Roundup Ready - po 2 i 4 godzinach inkubacji. W śrucie kukurydzianej GM wykryto
obecność specyficznej sekwencji dla kukurydzy MON 810 po 2 godzinach inkubacji u trzech
krów, natomiast po 4 godzinach inkubacji - jedynie u dwóch krów. Po 8 godzinach inkubacji
próbek w żwaczu, zarówno w przypadku poekstrakcyjnej śruty sojowej jak i ziarna
kukurydzy GM, nie stwierdzono obecności transgenicznego DNA. Powyższe wyniki
wskazują, że przy powolnym przepływie żwaczowym treści pokarmowej, transgeniczne
DNA ulega całkowitemu rozkładowi w żwaczu.
Od krów żywionych przez 2 tygodnie dietą z udziałem badanych materiałów paszowych
GM pobrano przez przetokę dwukrotnie po dwie próbki treści żwacza. Z każdej próbki
wyizolowano DNA bezpośrednio po pobraniu. Do izolacji DNA zastosowano zestaw
odczynników, pozwalający na wyizolowanie wyłącznie plazmidowego DNA specyficznego dla
mikroorganizmów. Wyizolowane DNA zamrażano w temp. około -20ºC. W celu oceny
poziomego transferu DNA pochodzącego z roślin zmodyfikowanych genetycznie (soja RR i
kukurydza MON810) zastosowano badanie jakościowe metodą PCR zgodnie z procedurą
badawczą PB-34/PS. Badania wykonano w Krajowym Laboratorium Pasz Instytutu
Zootechniki PIB w Szczecinie. Nie stwierdzono poziomego pasażu DNA pochodzącego z
materiałów paszowych GM do DNA plazmidowego mikroorganizmów bytujących w
żwaczu krów.
Analiza obecności transgenicznego DNA w mleku
Wyniki analizy próbek mleka krów żywionych z udziałem materiałów paszowych GM
nie wykazały obecności transgenicznego DNA o sekwencji specyficznej dla kukurydzy MON
810 oraz soi RR. Świadczy to o braku pasażu analizowanych transgenów z przewodu
pokarmowego do mleka.
66
Wymierne rezultaty realizacji zadań.
Wyniki uzyskane w omawianych badaniach pozwalają na sformułowanie
następujących uogólnieniem i wniosków dotyczących bezpieczeństwa stosowania
poekstrakcyjnej śruty sojowej produkowanej z roślin GM (Roundup Ready) oraz śruty z
ziarna kukurydzy GM (Bt, MON 810) w żywieniu zwierząt gospodarskich:
1. Nie stwierdzono negatywnego wpływu poekstrakcyjnej śruty sojowej z soi RR i śruty z
ziarna kukurydzy MON 810 na wskaźniki produkcyjne, jakość uzyskiwanych produktów
odzwierzęcych oraz status metaboliczny i zdrowotny organizmu, w tym efektywność
odpowiedzi immunologicznej - co wskazuje, że badane, genetycznie zmodyfikowane
materiały paszowe są równoważne pokarmowo, w żywieniu drobiu, trzody chlewnej i
bydła, z odpowiednimi paszami konwencjonalnymi (ziarnem kukurydzy i poekstrakcyjną
śrutą sojową).
2. Brak obecności transgenicznego DNA w dalszych częściach przewodu pokarmowego
(począwszy od treści jelit cienkich) świadczy o wysokiej efektywności jego trawienia u
badanych grup zwierząt gospodarskich oraz ogranicza możliwość przechodzenia
aktywnych fragmentów łańcucha kwasu nukleinowego do organizmu. Nie wykazano
obecności transgenicznego DNA w narządach wewnętrznych, krwi, tkance mięśniowej,
mleku i jajach, co wskazuje na brak pasażu wykrywalnych fragmentów transgenów z
przewodu pokarmowego do organizmu badanych gatunków oraz bezpieczeństwo
konsumpcji przez człowieka produktów pochodzenia zwierzęcego.
3. Wnioski wynikające z wykonanego zadania dotyczą wyłącznie badanych genetycznie
zmodyfikowanych materiałów paszowych, tj. poekstrakcyjnej śruty sojowej MON-40-3-2
oraz ziarna kukurydzy MON 810 (DKC 3421YG), tak więc nie powinny być uogólniane i
przenoszone na inne niż wymienione materiały paszowe uzyskiwane z roślin GM.
Wymiernym efektem wykonania zadania są publikacje naukowe, dotyczące
problematyki stosowania genetycznie zmodyfikowanych materiałów paszowych w żywieniu
zwierząt gospodarskich. Prace te były publikowane w formie twórczych artykułów
oryginalnych,
przeglądowych,
popularno-naukowych
i
doniesień
konferencyjnych,
przyczyniając się do upowszechniania wiedzy na temat organizmów GM. Ważną formą
popularyzacji uzyskanych wyników były także referaty, prezentowane w ramach konferencji
naukowych i naukowo technicznych. Referat dotyczący tego zagadnienia wygłoszono między
67
innymi na konferencji nt. „GMO jednym z czynników prorozwojowych polskiego rolnictwa i
obszarów wiejskich – prawda o szansach i zagrożeniach genetycznej modyfikacji roślin i
zwierząt”, która odbyła się 22 marca 2010 w Sejmie RP. Ze względu na wciąż
niewystarczającą wiedzę i emocje, jakie nadal wzbudza w społeczeństwie problem GMO,
stosowanie pasz GM w żywieniu zwierząt gospodarskich, szerokie upowszechnianie wyników
uzyskiwanych w badaniach stanowiło ważny element niniejszego zadania.
Wykaz publikacji przygotowanych w ramach zadania
1. Oryginalne prace twórcze w recenzowanych czasopismach naukowych:
Świątkiewicz S., Świątkiewicz M., Koreleski J., Kwiatek K. (2010). Nutritional efficiency of
genetically modified, insect resistant corn (MON810) and glyphosate tolerant soybean meal
(Roundup Ready) for broilers. Bulletin of the Veterinary Institute in Pulawy, 54, 43-48.
Świątkiewicz S., Twardowska M., Markowski J., Mazur M., Sieradzki Z., Kwiatek K. (2010). Fate
of transgenic DNA from Bt corn and Roundup Ready soybean meal in broilers fed GMO
feed. Bulletin of the Veterinary Institute in Pulawy, 54, 237-242.
Świątkiewicz S., Koreleski J., Arczewska-Włosek A., Twardowska M. (2010). Poekstrakcyjna
śruta sojowa i ziarna kukurydzy z genetycznie zmodyfikowanych roślin w żywieniu kur
nieśnych. Zeszyty Problemowe Postępów Nauk Rolniczych, 544, 11-18.
Świątkiewicz M., Hanczakowska E., Twardowska M., Mazur M., Kwiatek K., Kozaczyński W.,
Sieradzki Z., Świątkiewicz S. (2011). The effect of genetically modified feeds on fattening
results and transgenic DNA transfer to swine tissues. Bulletin of the Veterinary Institute in
Pulawy, 55, 121-125.
Stadnik J., Karwowska M., Dolatowski Z.J., Świątkiewicz S, Kwiatek K. (2011). Effect of
genetically modified, insect resistant corn (MON810) and glyphosate tolerant soybean meal
(Roundup Ready) on physico-chemical properties of broilers breast and thigh muscles.
Bulletin of the Veterinary Institute in Pulawy, 55, 541-546.
Świątkiewicz S., Koreleski J., Arczewska-Włosek A., Świątkiewicz M., Twardowska M.,
Markowski J., Mazur M., Sieradzki Z., Kwiatek K. (2011). Detection of transgenic DNA
from Bt maize and herbicide tolerant soybean soybean meal in tissues, eggs and contents of
segments of digestive tract in laying hens fed diets containing genetically modified plants.
Annals of Animal Science, 11, 413-424.
Stadnik J., Karwowska M., Dolatowski Z.J., Świątkiewicz M., Kwiatek K. (2011). Effect of
genetically modified feeds on physico-chemical properties of pork. Annals of Animal
Science, 11, 597-606.
68
2. Prace przeglądowe w recenzowanych czasopismach naukowych:
Świątkiewicz S., Koreleski J. (2008). Rośliny genetycznie zmodyfikowane w żywieniu drobiu.
Medycyna Weterynaryjna, 64, 1379-1383.
Świątkiewicz S., Świątkiewicz M. (2009): Rośliny genetycznie zmodyfikowane drugiej generacji
w żywieniu zwierząt gospodarskich. Medycyna Weterynaryjna, 65, 460-465.
Świątkiewicz S., Koreleski J., Twardowska M., Markowski J., Świątkiewicz M., Kwiatek K.,
Mazur M., Sieradzki Z., Pejsak Z., Tomczyk G., Bednarek D., Kozaczyński W., Reichert M.
(2010). Pasze genetycznie zmodyfikowane w żywieniu zwierząt – ocena zagrożeń na
podstawie wyników badań krajowych. Pasze Przemysłowe, 4/5/6, 18-31.
Świątkiewicz S., Arczewska-Włosek A. (2011). Prospects for the use of genetically modified
crops with improved nutritional properties as feed materials in poultry nutrition. World’s
Poultry Science Journal, 67, 631-642.
3. Artykuły popularno-naukowe:
Świątkiewicz S., Koreleski J. (2008). Rośliny genetycznie modyfikowane II i III generacji w
żywieniu drobiu. Polskie Drobiarstwo, 5, 8-11.
Świątkiewicz S., Koreleski J., Arczewska A., Twardowska M., Kwiatek K., Tomczyk G., Reichert
M., Mazur M., Bednarek D. (2010). Bezpieczeństwo transgenicznych materiałów paszowych
w żywieniu drobiu – wyniki doświadczeń krajowych. Życie Weterynaryjne, 2: 161-165.
Świątkiewicz S., Koreleski J., Arczewska A., Twardowska M., Kwiatek K., Tomczyk G., Reichert
M., Mazur M., Bednarek D. (2010). Krajowe badania nad stosowaniem pasz genetycznie
zmodyfikowanych w żywieniu drobiu. Cz. 1. Polskie Drobiarstwo, 1:10-13.
Świątkiewicz S., Koreleski J., Arczewska A., Twardowska M., Kwiatek K., Tomczyk G., Reichert
M., Mazur M., Bednarek D. (2010). Krajowe badania nad stosowaniem pasz genetycznie
zmodyfikowanych w żywieniu drobiu. Cz. 2. Polskie Drobiarstwo, 2: 2-5.
Świątkiewicz S., Koreleski J., Arczewska A. (2010) Pasze GMO w żywieniu zwierząt
gospodarskich. Nowa Wieś Europejska, 1-2: 6-8.
4. Doniesienia na konferencje naukowe i naukowo-techniczne:
Świątkiewicz S., Koreleski J., Arczewska A. (2008). Wstępne wyniki badań nad stosowaniem pasz
z roślin genetycznie modyfikowanych w żywieniu kurcząt brojlerów. Materiały V
Konferencji Młodych Badaczy: “Fizjologia i biochemia w żywieniu zwierząt, Warszawa, 2223.09.2008, str. 146-148.
69
Świątkiewicz S., Koreleski J., Arczewska A. (2008). Performance of broilers fed diets containing
genetically modified components. Materiały XX International Poultry Symposium PB
WPSA, Bydgoszcz, Wenecja, 15-17.09.2008, str.147.
Świątkiewicz M. (2009). Rośliny modyfikowane genetycznie w żywieniu świń. Materiały I
Kongresu Nauk Rolniczych „Nauka Praktyce”. Puławy, 14-15.05.2009, str. 79-80.
Świątkiewicz S., Koreleski J., Arczewska A. (2009). Zastosowanie surowców paszowych z roślin
genetycznie modyfikowanych w żywieniu drobiu. Materiały I Kongresu Nauk Rolniczych
„Nauka Praktyce”. Puławy, 14-15.05.2009, str. 83-84.
Świątkiewicz M., Hanczakowska E. (2009). Wpływ pasz genetycznie modyfikowanych na wyniki
tuczu świń i jakość mięsa. Materiały Konferencyjne XXXVIII Sesji Naukowej Komisji
Żywienia Zwierząt KNZ PAN “Pasze zmodyfikowane genetycznie (GMO) i pasze tradycyjne
w żywieniu zwierząt”. Balice, 28-29.05.2009, str. 91-92.
Świątkiewicz S., Koreleski J., Arczewska A. (2009). Wskaźniki produkcyjne i jakość jaj u kur
oraz wyniki odchowu brojlerów żywionych dietą z udziałem surowców genetycznie
modyfikowanych. Materiały Konferencyjne XXXVIII Sesji Naukowej Komisji Żywienia
Zwierząt KNZ PAN “Pasze zmodyfikowane genetycznie (GMO) i pasze tradycyjne w
żywieniu zwierząt”. Balice, 28-29.05.2009., str. 101-102.
Furgał-Dierżuk I., Strzetelski J. (2009). Rozkład w żwaczu genetycznie modyfikowanej soi oraz
kukurydzy. Materiały Konferencyjne XXXVIII Sesji Naukowej Komisji Żywienia Zwierząt
KNZ PAN “Pasze zmodyfikowane genetycznie (GMO) i pasze tradycyjne w żywieniu
zwierząt”. Balice, 28-29.05.2009, str. 171-172.
Świątkiewicz S., Koreleski J., Arczewska-Włosek A., Twardowska M. (2010). Soybean meal and
maize grain from genetically modified plants in laying hens nutrition. Materiały XXXIX Sesji
Naukowej Komisji Żywienia Zwierząt KNZ PAN: “Żywienie w regulacji rozwoju i
produkcyjności zwierząt”. Rynia k/Warszawy, 26-28.05.2010, str. 63.
Świątkiewicz M., Hanczakowska E., Twardowska M., Kwiatek K., Mazur M., Kozaczyński W.,
Świątkiewicz S., (2010). The effect of genetically modified feeds on fattening results and
transfer of transgenic DNA to swine tissues. Materiały XXXIX Sesji Naukowej Komisji
Żywienia Zwierząt KNZ PAN: “Żywienie w regulacji rozwoju i produkcyjności zwierząt”.
Rynia k/Warszawy, 26-28.05.2010, str. 97.
Furgał-Dierżuk I., Strzetelski J., Kwiatek K., Twardowska M., Mazur M. (2010). The effect of
genetically modified feeds on rearing performance and tDNA transfer in the digestive tract
and into tissues of calves. Materiały XXXIX Sesji Naukowej Komisji Żywienia Zwierząt
KNZ PAN: “Żywienie w regulacji rozwoju i produkcyjności zwierząt”. Rynia k/Warszawy,
70
26-28.05.2010, str. 19.
Furgał-Dierżuk I., Strzetelski J., Kwiatek K., Twardowska M., Mazur M. (2010). The effect of
genetically modified feeds on productivity, milk composition, serum metabolite profiles and
transfer of tDNA into milk cows. Materiały XXXIX Sesji Naukowej Komisji Żywienia
Zwierząt KNZ PAN: “Żywienie w regulacji rozwoju i produkcyjności zwierząt”. Rynia
k/Warszawy, 26-28.05.2010, str. 159.
Świątkiewicz S., Koreleski J., Twardowska M., Markowski J., Świątkiewicz M., Kwiatek K.,
Mazur M., Sieradzki Z., Pejsak Z., Tomczyk G., Bednarek D., Kozaczyński W., Reichert M.
(2010). Pasze genetycznie zmodyfikowane w żywieniu zwierząt gospodarskich – ocena
zagrożeń na podstawie wyników badań IZ PIB. Materiały XXVII Międzynarodowa
Konferencji Naukowo-Technicznej: „Analiza zagrożeń i analiza ryzyka w łańcuchu
paszowym”, Zwierzyniec, 23.04.2010, str. 13-15.
Świątkiewicz M., Hanczakowska E., Twardowska M., Markowski J. (2010). Stosowanie pasz
genetycznie zmodyfikowanych a jakość produktu zwierzęcego.
Materiały XXVII
Międzynarodowej Konferencji Naukowo-Technicznej: „Analiza zagrożeń i analiza ryzyka w
łańcuchu paszowym”, Zwierzyniec, 23.04.2010, str. 43-44.
5. Referaty:
Świątkiewicz S. (2009). Rośliny genetycznie modyfikowane w żywieniu zwierząt gospodarskich.
Konferencja Naukowa Lubelskiego Towarzystwa Naukowego i Katedra Higieny Żywności
Zwierzęcego Pochodzenia „Organizmy Modyfikowane Genetycznie – Szanse i Zagrożenia”.
Lublin, 22.05.2009. Str. 9-20.
Świątkiewicz S. (2009) Aktualna sytuacja w uprawie roślin GM na świecie. Pasze z roślin
genetycznie modyfikowanych w żywieniu zwierząt. Wykłady dla studentów studium
doktoranckiego Instytutu Zootechniki-PIB.
Kwiatek K., Świątkiewicz S. (2010). Pasze zmodyfikowane genetycznie (GMO) w żywieniu
zwierząt gospodarskich, a zdrowie zwierząt i bezpieczeństwo żywności pochodzenia
zwierzęcego. Konferencja naukowa: „GMO jednym z czynników prorozwojowych polskiego
rolnictwa i obszarów wiejskich – prawda o szansach i zagrożeniach genetycznej modyfikacji
roślin i zwierząt”, Sejm RP, Warszawa, 22.03.2010.
Świątkiewicz S., Koreleski J., Twardowska M., Markowski J., Świątkiewicz M., Kwiatek K.,
Mazur M., Sieradzki Z., Pejsak Z., Tomczyk G., Bednarek D., Kozaczyński W., Reichert M.
(2010). Pasze genetycznie zmodyfikowane w żywieniu zwierząt gospodarskich – ocena
zagrożeń na podstawie wyników badań IZ PIB. XXVII Międzynarodowa Konferencja
Naukowo-Techniczna: „Analiza zagrożeń i analiza ryzyka w łańcuchu paszowym”,
71
Zwierzyniec, 23.04.2010.
Świątkiewicz S., Koreleski J., Świątkiewicz M., Kwiatek K. (2010). Bezpieczeństwo stosowania
pasz
genetycznie
zmodyfikowanych
–
wyniki
badań
krajowych.
Seminarium:
„Wykorzystanie biotechnologii w rolnictwie amerykańskim i polskim”, Kraków, 14.04.2010.
Świątkiewicz S., Brzóska F. (2011). Wyniki badań krajowych nad bezpieczeństwem stosowania
pasz genetycznie zmodyfikowanych w żywieniu zwierząt. Konferencja XIX Szkoła Zimowa
Hodowców Bydła „Praktyka nauce – nauka praktyce”, 04-08 kwietnia 2011, Zakopane.
Brzóska F., Świątkiewicz S. (2011). Możliwość wykorzystania pasz GMO w żywieniu zwierząt w
oparciu o badania krajowe. II Kongres Nauko Rolniczych, Konferencja Naukowo-Techniczna
„Innowacyjność w produkcji zwierzęcej – oczekiwanie czy konieczność”, 13-14 czerwca
2011, IZ-PIB, Balice.
Tomczyk G., Minta Z., Kwiatek K., Świątkiewicz S. Sieradzki Z. (2011). Surowce GMO w
żywieniu drobiu a jego status zdrowotny. XXIII Międzynarodowe Sympozjum Drobiarskie
PO WPSA „Nauka praktyce drobiarskiej – praktyka drobiarska nauce”, 13-15 września 2011,
Poznań.
Rola partnerów w realizacji zadań (ze szczególnym uwzględnieniem organów
administracji publicznej)
Zadanie
zostało
zrealizowane
we
współpracy
z
Państwowym
Instytutem
Weterynaryjnym - PIB w Puławach. Rola PIWet-PIB w zadaniu polegała na ocenie
parametrów charakteryzujących status zdrowotny zwierząt żywionych bez udziału lub z
udziałem materiałów paszowych GM, a także na analizie transferu transgenicznego DNA w
organizmie.
Prace
PIWet-PIB
immunologicznej
po
obejmowały
wykonanych
między
innymi
szczepieniach
analizę
ochronnych,
stanu
odpowiedzi
analizę
parametrów
morfologicznych i immunologicznych krwi, badania makroskopowe oraz histologiczne
narządów wewnętrznych i błony śluzowej przewodu pokarmowego, analizy obecności
transgenicznego DNA w poszczególnych odcinkach przewodu pokarmowego oraz w
wybranych narządach i tkankach.
72