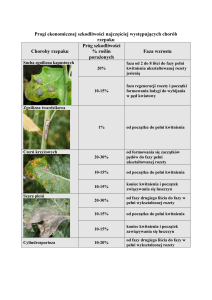
Regulacja kwitnienia
Wybór właściwego czasu
kwitnienia jest kluczowy dla
sukcesu reprodukcyjnego
roślin
Ewolucja doprowadziła
do powstania wielu
ścieżek regulujących
czas kwitnienia
Rośliny kwiatowe przechodzą fazę wzrostu wegetatywnego
(wytwarzanie pędów i liści) i fazę kwitnienia, w trakcie której wytwarzają
organy służące do rozmnażania płciowego
•
U roślin jednorocznych faza
wegetatywna zaczyna się w momencie
kiełkowania nasion. Następująca po
niej faza kwitnienia kończy się
starzeniem się i śmiercią rośliny.
•
U roślin dwuletnich, faza wegetatywna
trwa przez pierwszy rok, w drugim
roku następuje kwitnienie, które
kończy się śmiercią rośliny.
•
U roślin wieloletnich, kwitnienie
następuje co rok, przez wiele lat.
Wzrost wegetatywny pędu następuje w merystemie wierzchołkowym. Jest to masa
niezróżnicowanych komórek na szczycie pędu. Podziały mitotyczne tych komórek
wytwarzają komórki, które różnicują w części pędu, liście, wtórne merystemy (zwane też pączkami
bocznymi) – dają początek rozgałęzieniom pędu.
Kwitnienie jest regulowane przez wiele czynników
Kwitnienie wymaga
przekształcenia merystemu
wierzchołkowego w merystem
kwiatowy
Zależy to od:
•
•
Czynników wewnętrznych
Czynników zewnętrznych
Regulacja przez temperaturę
•
•
Stan uśpienia pąków jest zlokalizowany.
Wiele roślin jednorocznych (np.
pszenica ozima) i dwuletnich ma
opóźniony czas kwitnienia, jeśli nie
przejdzie w trakcie zimy okresu
zimowego przechłodzenia. Zmiany
powodowane przez ten okres
zimowego przechłodzenia noszą
nazwę wernalizacji.
U wielu drzewiastych roślin
kwiatowych rosnących w klimacie
umiarkowanym (jabłonie, bzy),
kwitnienie wymaga uprzedniej
ekspozycji na niską temperaturę.
Drzewa te nie kwitną w klimacie
ciepłym, w którym nie ma wyraźnych
zim.
Regulacja przez fotoperiod (stosunek długości dnia do
nocy)
•
•
•
Fotoperiod jest wykrywany w liściach
(np. roślina X potrzebuje dnia o
długości co najmniej 8,5 godziny, by
móc zakwitnąć). Jednak wystarczy, by
tylko jeden liść był eksponowany na
właściwy fotoperiod, aby kwiaty
pojawiały się na całej roślinie.
Liście produkują sygnał chemiczny florigen, który jest transportowany
do merystemów wierzchołkowych.
Chemiczna natura florigenu nie jest do
końca wyjaśniona, jednym z jego
składników może być białko kodowane
przez gen FT (Flowering locus T).
Florigen może się przemieszczać poprzez system naczyniowy
Budowa kwiatu
•
•
1
2
•
3
4
•
•
Merystem kwiatowy różnicuje w cztery
koncentryczne okółki (kręgi) komórek,
które tworzą następnie cztery części
kwiatu.
Komórki w okółku 1 rozwijają się w
działki kielicha, tworzące najniższy
poziom. Łącznie działki tworzą tzw.
kielich.
Okółek 2 daje początek umieszczonym
nad kielichem płatkom, tworzącym
razem koronę kwiatu. Korona kwiatu
jest jego najbardziej barwną częścią.
Okółek 3 rozwija się w pręciki, męskie
organy płciowe.
Okółek 4 (najbardziej wewnętrzny)
tworzy słupki, narządy płciowe
żeńskie. Często zlewają się w
pojedynczą strukturę.
Model ABC rozwoju kwiatu
•
•
•
Geny ABC:
Grupa A: Apetala1 (AP1)
•
Apetala2 (AP2)
Grupa B: Apetala3 (AP3)
Pistilata (PI)
Grupa C: Agamous (AG)
•
•
Wyniki analizy genetycznej mutantów
Arabidopsis i Petunii, sugerowały, że istnieje
grupa genów kodujących czynniki
transkrypcyjne (główne włączniki) niezbędne
do włączania genów warunkujących rozwój
działek kielicha, płatków korony, pręcików i
słupków.
Te główne włączniki należą do trzech klas:
A, B i C.
Komórki, w których wyrażane są tylko geny
klasy A tworzą działki kielicha.
Komórki, w których wyrażane są zarówno
geny klasy A jak i klasy B, tworzą płatki
korony.
Komórki, w których wyrażane są zarówno
geny klasy B jak i klasy C, tworzą pręciki.
Komórki, w których wyrażane są tylko geny
klasy C tworzą słupki.
Efekty mutacji w genach ABC
Relacje między genami ABC
•
Geny klasy A i C są w stosunku do
siebie represorami. W
nieobecności A, C są aktywne w
całym kwiecie. W nieobecności C,
A są aktywne w całym kwiecie.
Efekty mutacji w AP1
Geny klasy E (SEP) uzupełniają geny modelu ABC
•
Geny SEP (SEPALLATA), obok
genów ABC, są niezbędne do
prawidłowego określania
tożsamości organów kwiatowych
Większość genów ABCE koduje czynniki
transkrypcyjne z domeną MADS
MADS
Domena K
Domena C
•
Domena MADS występuje na N-końcu białka i warunkuje: wiązanie do DNA,
zdolność do dimeryzacji i lokalizację jądrową.
•
•
(tylko białko AP2 nie należy do rodziny białek MADS)
Arabidopsis zawiera ponad 100 genów kodujących różne białka z domeną MADS.
Niezależna ewolucja genetycznych narzędzi kontroli
rozwoju u zwierząt i roślin
ANIMALS
PLANTS
Pattern formation in development
Embryo development
Flower development
Master regulatory genes contain
homeobox (Hox genes)
Master regulatory genes
contain MADS box
Dorsal-ventral specification controlled
by TGF-related proteins (GURKEN),
receptor tyrosine kinases, Ras
activation, transcription factors
of kappa B, Rel, basic HLH families
No relatives of GURKEN
No receptor tyrosine kinases
No Ras proteins
No kappa B or Rel –type TFs
Weak similarity of the bHLH
domain
Cell-cell signaling
Critical role of receptor tyrosine kinases, Critical role of serine/threonine
Ras activation
kinases of the type not found in
animals
Chromatin
Histones, histone modifying proteins, Swi/Snf-type
ATPases, Trx proteins, Polycomb proteins, HP1-type proteins.
Enahncer of zeste (Polycomb-type
maintains repression of
the Hox genes (Ultrabithorax)
CURLY LEAF (Polycomb type)
maintains repression of the MADS
genes (AGAMOUS)
Meyerowitz, EM. Science (2002)
Centralna rola genu LFY (LEAFY)
•
•
•
1.
2.
Ortologi LFY występują u wszystkich gatunków roślin (także nie
kwiatowych).
Aktywność LFY jest konieczna i wystarczająca do determinacji
merystemu kwiatowego.
Niezależnie od determinacji typu merystemu, LFY pełni dwie kluczowe
funkcje w rozwoju kwiatu:
Jest głównym integratorem sygnałów prowadzących do indukcji
kwitnienia.
Jest głównym aktywatorem genów ABCE.
Cztery fazy rozwoju kwiatu
1.
W odpowiedzi na sygnały ze środowiska i sygnały wewnętrzne roślina
przestawia się z wzrostu wegetatywnego na wzrost reprodukcyjny – ten
proces kontrolują geny regulujące czas kwitnienia (flowering time
genes).
2.
Sygnały z różnych ścieżek wpływających na czas kwitnienia są
integrowane, co prowadzi do aktywacji niewielkiej grupy genów
tożsamości merystemu (meristem identity genes), które warunkują
powstanie kwiatu (Geny TFL1, LFY, AP1).
3.
Geny tożsamości merystemu aktywują geny tożsamości organów
kwiatowych (Geny ABCE).
4.
Geny tożsamości organów aktywują zależne od nich geny „budujące
organy”, które determinują różne typy komórek, z których składają się
poszczególne organy kwiatu.
Genetyczna kontrola czasu kwitnienia
Geny kontrolujące czas kwitnienia działają
w czterech ścieżkach indukcji:
A
D
C
B
A.
Zależnej od fotoperiodu (rytm dobowy,
długi dzień);
B.
Zależnej od giberelin (GA);
C.
Autonomicznej;
D.
Wernalizacyjnej
U Arabidopsis liczba liści w rozecie jest miarą czasu kwitnienia
Analiza czasu kwitnienia w Arabidopsis, efekt mutacji w genach SWI3
Leaf number at flowering during short day
Leaf number at flowering during long day
%
%
days leaf No
C-65-75 /27
D-60-75 /34
Wt-58-67 /54
20
60,0
days leaf No
C-24-25 /9-10
D-24-25 /11
Wt-20-21 /11
18
50,0
16
14
40,0
12
10
30,0
8
20,0
6
4
10,0
2
atswi3c
atswi3d
wt
atswi3c
60
57
56
54
52
50
46
44
42
48
wt
%
60
60
50
50
40
40
30
30
20
20
10
10
C-3-4
D-6-7
Wt-8
0
0
C
atswi3d
Number of secondary inflorescences
Number of cauline leaves
%
C-3-4
D-6-7
Wt-8
40
B
38
16
36
15
34
14
32
13
30
12
28
11
26
A
10
24
9
22
0
0,0
1
2
3
4
5
atswi3c
6
7
8
atswi3d
wt
9
10
11
D
0
1
2
3
4
atswi3c
5
6
atswi3d
7
8
9
10
11
wt
Sarnowski et al.,Plant Cell 2005
atswi3c: early flowering in SD, slightly early flowering in LD
atswi3d: early flowering in SD
Kontrola czasu kwitnienia – fotoperiod (long day pathway)
•
•
•
AP1
LFY
Wiele genów indukujących kwitnienie w długim
dniu koduje białka zaangażowane w percepcję
światła (PHYTOCHROME A,
CRYPTOCHROM2) lub składniki regulujące
zegar okołodobowy (GIGANTEA, ELF3).
Geny te ostatecznie aktywują gen CO
(CONSTANS) (mutany co są późnokwitnące,
nadekspresja CO powoduje wczesne kwitnienie).
CO koduje białko jądrowe zawierające dwie
domeny palców cynkowych.
Kontrola czasu kwitnienia – gibereliny
•
AP1
LFY
Mutanty z defektem w biosyntezie giberelin (np.
ga1) są bardzo późno kwitnące w krótkim dniu,
ale nie w długim, co wskazuje, że szlak GA ma
kluczowe znaczenie w indukcji kwitnienia w
sytuacji braku sygnału indukującego długiego
dnia (fotoperiodycznego)
Kontrola czasu kwitnienia – szlak autonomiczny
i szlak wernalizacyjny
•
•
•
AP1
LFY
Geny szlaku autonomicznego kontrolują
kwitnienie niezależne od długiego dnia
(Arabidopsis jest normalnie rośliną kwitnącą w
długim dniu, ale po dłuższym okresie zakwita
także w krótkim dniu). Kwitniecie w krótkim
dniu jest indukowane przez geny szlaku
autonomicznego.
Geny szlaku wernalizacyjnego są związane z
indukcją kwitnienia zależną od przejścia
zimowego przechłodzenia.
Geny FLC, SOC1, FT i LFY pełnią funkcję
integratorów sygnałów z różnych szlaków
kontroli czasu kwitnienia.
Ekspresja okołodobowa głównych elementów
ścieżki związanej z fotoperiodem (długością dnia)
a. GI i CDF1 są kontrolowanymi przez zegar okołodobowy
odpowiednio, pozytywnymi i negatywnymi regulatorami CO.
FKF1 jest niezbędny do degradacji CDF1 w środku dnia, co
bezpośrednio umożliwia wzrost ilości mRNA CO.
b. Profil ekspresji CO w krótkim i długim dniu jest inny. Dwufazowa krzywa występuje tylko w długim dniu.
c. Akumulacja białka CO silnie zależy od koincydencji światła i
ekspresji mRNA. CO jest degradowane w ścieżce proteasomowej,
ale w sposób zależny od receptorów światła i białek SPA.
d. Wytwarzanie mRNA FT jest bezpośrednim rezultatem
akumulacji białka CO.
Sieć regulatorowa kontrolująca ekspresje mRNA genu CO
I stabilność białka CO
Regulacja transkrypcyjna
Symbol zegara oznacza, że
transkrypcja genu jest
regulowana przez zegar
okołodobowy
Zygzakowana strzałka oznacza,
że białka jest fotoreceptorem
Regulacja
potranskrypcyjna
Akumulacja białka CO
powoduje aktywację genu FT
Koincydencja akumulacji mRNA CO i światła i pod koniec długiego dnia stabilizuje
białko CO. Białko CO umożliwia następnie transkrypcję FT i TSF (Twin Sister of
FT, bardzo podobne białko) w komórkach towarzyszących rurkom sitowym (floem),
w odległej części liścia. Białko FT i prawdopodobnie TSF są ładowane do
elementów sitowych poprzez dyfuzje przez plazmodesmy albo inny
niezidentyfikowany mechanizm aktywnego transportu.
Następuje długodystansowy transport FT poprzez floem wraz innymi metabolitami
transportowanymi z liścia do tkaenk docelowych
W wyniku transportu FT i TSF akumulują się w merystemie wierzchołkowym pędu (SAM),
gdzie prawdopodobnie wytwarzają gradient
FT i czynnik transkrypcyjny bZIP FD oddziałują i aktywują transkrypcję AP, który
rozpoczyna przekształcenie w merystem kwiatowy i hamuje ekspresję SOC1 (Suppressor of
overexpresion of CO).
Białko FT stanowi sygnał
systemiczny
TSF – Twin sister of FT
SOC1 – Suppresor of overexpresion of CO
Dynamika ekspresji genów w merystemie wierzchołkowym pędu (SAM)
w trakcie przejścia do fazy kwitnienia
Indukcja
Wegetatywny
Kwiatowy
Czerwony – mRNA
Niebieski - białko
SWI/SNF and control of flowering time
BRM
SWI3BSWI3C
BSH
AP1
LFY
Kompleks SWI/SNF pośredniczy
w sygnalizacji uruchamianej
przez gibereliny