&ARM0RZEGL.AUK
COPYRIGHT'RUPADR!2+WIECIÊSKIEGO)33.
%KSPRESJAGENÌWZWIZANYCHZKASKADKASPAZ
WKOMÌRKACHBIAACZKIPROMIELOCYTARNEJ(,
EKSPONOWANYCHNACISPLATYNÃ
%XPRESSIONOFGENESASSOCIATEDWITHCASPASESCASCADE
INPROMYELOTICLEUKEMIACELLS(,EXPOSEDTOCISPLATIN
%WELINA!LEKSANDER'RZEGORZ(IBNER!LICJA:AJDEL-ONIKA0AUL3AMOJEDNY!DAM7ILCZOK
+ATEDRAI:AKAD"IOFARMACJI7YDZIA&ARMACEUTYCZNYZ/DDZIAEM-EDYCYNY,ABORATORYJNEJ
gLSKI5NIWERSYTET-EDYCZNYW+ATOWICACH
+ATEDRAI:AKAD'ENETYKI-EDYCZNEJ7YDZIA&ARMACEUTYCZNYZ/DDZIAEM-EDYCYNY,ABORATORYJNEJ
gLSKI5NIWERSYTET-EDYCZNYW+ATOWICACH
Streszczenie
Abstract
Cisplatyna (CPT) jest jednym z najpowszechniejszych
chemioterapeutyków stosowanych w terapii nowotworów.
Uszkodzenie genomowego DNA po wniknięciu CPT do
jądra komórkowego wyzwala cytotoksyczne procesy prowadzące do śmierci komórek nowotworowych. Aktywacja kaskady proteolitycznej z udziałem członków rodziny
kaspaz jest kluczowym elementem śmierci w komórkach
apoptotycznych. Stosując technikę mikromacierzy oligonukleotydowych (Affymetrix) porównano ekspresję
genów związanych z kaskadą kaspaz w komórkach białaczki promielocytarnej HL-60 eksponowanych na CPT
oraz w komórkach kontrolnych. Analiza sygnałów fluorescencji 1772 sond, wyselekcjonowanych z bazy NetAffx
Analysis Center, umożliwiła wykazanie wzrostu ekspresji
genów BIK, SAFB, BAX, SFRS2IP, FUT7, SIGMAR1,
RALBP1, USO1, CASP6 oraz spadku poziomu ekspresji
TNFAIP3, CD48 i PDE4B. Wyjaśnienie zmian ekspresji genów związanych z kaskadą kaspaz wywoływanych
przez CPT może mieć istotne znaczenie w ustalaniu schematu terapii przeciwnowotworowej z zastosowaniem tego
leku i przewidywaniu jej skuteczności.
Cisplatin (CPT) is one of the most popular chemiotherapeutics used in anticancer therapy. Damage followed
by CPT binding to DNA after its incorporation to a cell
nucleus triggers the cytotoxic processes leading to cancer
cell death. Activation of proteolytic cascades, involving
members of the family of caspases, is the key element
causing the apoptotic cells death. Using oligonucleotide
microarray technique (Affymetrix) we compared the expression of genes involved in the cascade of caspases in
promyelotic leukemia HL-60 cells exposed to CPT and
in control cells. Analysis of the fluorescence signals of
1772 probes, selected from the NetAffx Analysis Center
database, showed the increased expression of BIK, SAFB,
BAX, SFRS2IP, FUT7, SIGMAR1, RALBP1, USO1,
CASP6 and inhibition of TNFAIP3, CD48 and PDE4B.
Explaining changes in expression of genes associated with
caspases cascade induced by CPT may have a significant
role in determining the regimen of anticancer therapy and
predicting its effectiveness.
Key words: cisplatin, gene expression, caspases, caspases
cascade, promyelotic leukemia cells, HL-60
Słowa kluczowe: cisplatyna, ekspresja genów, kaspazy, kaskada kaspaz, komórki białaczki promielocytarnej, HL-60
Wstęp
Większość aktualnie stosowanych terapii przeciwnowotworowych wywołuje śmierć komórek nowotworowych poprzez indukcję apoptozy [1]. Jednym z najczęściej stosowanych chemioterapeutyków jest cisplatyna (CPT), powszechnie stosowana w rakach płuc, głowy, szyi, przełyku, żołądka,
okrężnicy, pęcherza moczowego, jąder, jajników i szyjki macicy oraz w większości innych zaawansowanych raków, takich
jak rak piersi, trzustki, wątroby, nerek, prostaty, jak również
przeciwko glejakom, czerniakom przerzutowym oraz międzybłoniakowi otrzewnej i międzybłoniakowi opłucnej [2].
Przyłączenie CPT do genomowego DNA (gDNA) w jądrze komórkowym jest głównym wydarzeniem odpowiedzialnym za przeciwnowotworowe właściwości tego związku. W ten sposób, uszkodzenie powstałe po tym połączeniu
może hamować transkrypcję oraz mechanizmy replikacji.
Następnie zmiany te mogą wyzwalać cytotoksyczne procesy
prowadzące do śmierci komórek nowotworowych [3]. Jednakże, selektywne defekty wewnątrzkomórkowych białek
sygnalizacyjnych i kaspaz mogą znosić wrażliwość nowotworów na bodźce apoptotyczne, co w konsekwencji prowadzi do oporności komórek nowotworowych na różne formy
chemioterapii i immunoterapii, których skuteczność zależy
&ARM0RZEGL.AUK
od indukcji apoptozy. Aktywacja kaskady proteolitycznej
z udziałem członków rodziny kaspaz jest kluczowym elementem realizacji śmierci w komórkach apoptotycznych [1].
Kaspazy to wewnątrzkomórkowe enzymy z rodziny proteaz
cysteinowych (ang. cysteine-dependent aspartate-directed
proteases) [4], które odgrywają główną rolę w kontrolowaniu procesu apoptozy, a także związane są z funkcjonowaniem układu immunologicznego [5]. Pierwszą, zidentyfikowaną w 1993 roku kaspazą był enzym konwertujący IL-1β
– ICE (ang. interleukin 1β converting enzyme), posiadający
zdolność przekształcenia prekursora pre-IL-1β w aktywną
cytokinę [4]. Kaspazy reprezentują 3 podgrupy białek. Bazując na homologii ich sekwencji aminokwasowych, dzielone są na: inicjatorowe - aktywujące inne kaspazy, efektorowe - uczestniczące w dalszych etapach apoptozy oraz zapalne - związane z cytokinami [6, 7]. Poszczególne kaspazy
różnią się od siebie swoistością substratową oraz sposobem
aktywacji. Wszystkie poznane dotychczas kaspazy są syntetyzowane jako nieaktywne proenzymy [8].
Celem prezentowanej pracy było określenie, jakie zmiany w ekspresji genów związanych z kaskadą kaspaz, wywołuje CPT w ludzkich komórkach białaczki promielocytarnej
(HL-60). Do analizy ekspresji genów zastosowano technikę mikromacierzy oligonukleotydowych Human Genome
U133A Set (HG-U133A; Affymetrix).
Materiał i metody
Hodowle komórkowe
Materiałem, który został wykorzystany do badań zmiany
ekspresji genów z wykorzystaniem mikromacierzy oligonukleotydowych, były hodowane ludzkie komórki białaczki
promielocytarnej (HL-60). Komórki linii HL-60 hodowano
w mieszaninie RPMI 1640 (Gibco) zawierającej 10% płodowej surowicy bydlęcej (FBS; Sigma), 2 mM glutaminy
(Sigma), 1 mM pirogronianu sodu (Sigma), 4,5 g/l glukozy,
100 μg/ml streptomycyny i 100 U/ml penicyliny (Sigma).
Hodowle komórkowe prowadzono w warunkach stałej wilgotności powietrza (95%), stałej temperatury (37°C) oraz
atmosfery 5% CO2. Po inkubacji, trwającej 48 godzin, dodawano pożywkę zawierającą CPT (Sigma) rozpuszczoną
w DMSO (Sigma), tak aby w hodowlach uzyskać stężenie
0,22 μg/ml, zgodne z wcześniej wyznaczoną wartością ID30
(Inhibitory Dose 30%). Hodowle stanowiące układ odniesienia prowadzono w identyczny sposób, dodając po 48 godzinach pożywkę z dodatkiem DMSO, tak by jego stężenie
było identyczne jak w próbach z CPT. Po trwającej 6 godzin
inkubacji komórki zbierano i przygotowano do ekstrakcji
RNA.
Ekstrakcja całkowitego RNA
Ekstrakcję całkowitego RNA przeprowadzono
z wykorzystaniem odczynnika TRIZOL (Invitrogen Life
Technologies) zgodnie z zaleceniami producenta. Uzyskane ekstrakty oczyszczano przy użyciu RNeasy Mini
Kit (Qiagen). Następnie oceniano je jakościowo techniką
elektroforezy agarozowej w obecności bromku etydyny
oraz ilościowo w oparciu o pomiar spektrofotometryczny
z wykorzystaniem GeneQuant II (Pharmacia Biotech; λ
= 260 nm).
Analiza aktywności transkrypcyjnej genów techniką mikromacierzy oligonukleotydowych HG-U133A (Affymetrix)
Całkowity RNA, w ilości 8μg, został następnie wykorzystany do syntezy dwuniciowego cDNA (Gibco BRL SuperScript Choice system), który stanowił matrycę do syntezy
biotynylowanego cRNA (reakcja transkrypcji in vitro, Enzo
kit). W dalszym etapie, po fragmentacji biotynylowanego
cRNA, przeprowadzono hybrydyzację z mikromacierzą
Test3, a następnie, po pozytywnej weryfikacji, hybrydyzację z mikromacierzą HG-U133A (Affymetrix). Produkty hybrydyzacji znakowano kilkustopniowo kompleksem streptawidyna – fikoerytryna, znakowanymi przeciwciałami
przeciw biotynie oraz przeciwciałami przeciw kompleksowi streptawidyna – fikoerytryna zgodnie z protokołem
EukGEWS2v4 zalecanym dla mikromacierzy oligonukleotydowej HG–U133A w przewodniku Gene Expression
Analysis Technical Manual (Affymetrix). W kolejnym
etapie płytki mikromacierzy odczytywano używając skanera Agilent GeneArray Scanner G2500A (Agilent Technologies). Otrzymane wyniki intensywności fluorescencji zapisano i zarchiwizowano w programie Microarray
Suite oraz specjalnie do tego celu przygotowanej bazie
danych.
Analizę porównawczą transkryptomów przeprowadzono, po normalizacji wyników w programie RMA Express,
polegającej na przekształceniu logarytmicznym wartości
sygnału fluorescencji dla każdego transkryptu (log2), wykorzystując programy komputerowe: Statistica v.7,0, Gnumeric, oraz Microsoft Excel. Spośród znajdujących się na
mikromacierzy HG-U133A sond, wykorzystując dane zaczerpnięte z bazy Affymetrix NetAffxTM Analysis Center
(http://www.affymetrix.com/analysis/index.affx) po wpisaniu kwerend: „caspases” i „caspase”, wyselekcjonowano te
określające ekspresję genów związanych z kaskadą kaspaz.
Sumaryczny zbiór 1772 sond stanowił podstawę do poszukiwania genów, których sygnał ekspresji różnił się w próbach
badanych i kontrolnych, co najmniej, ±2STD (dwukrotna
wartość odchylenia standardowego).
Wyniki
W niniejszej pracy porównywano ekspresję genów
związanych z kaskadą kaspaz komórek eksponowanych
na CPT względem hodowli kontrolnych. Posługując się
programem komputerowym Gnumeric wyfiltrowano
1772 sygnały fluorescencji, charakteryzujące ekspresję
wybranych transkryptów związanych z kaskadą kaspaz,
z grupy 22283 sond mRNA, obecnych na mikromacierzy
HG-U133A (Affymetrix). Porównując wartości parametru SLR (stosunek zlogarytmowanych uśrednionych wartości fluorescencji transkryptu w porównywanych próbach, ang. signal log ratio), wyznaczono geny związane
z kaskadą kaspaz, których ekspresja pod wpływem CPT
w największym stopniu uległa zmianie, a SLR różnił się,
co najmniej ±2STD w odniesieniu do komórek hodowli kontrolnych. W komórkach HL-60 eksponowanych na
CPT w największym stopniu stymulowana była ekspresja genów BIK, SAFB, BAX, SFRS2IP, FUT7, SIGMAR1,
RALBP1, USO1, CASP6. Genami, których ekspresja
została najbardziej zahamowana, okazały się TNFAIP3,
COPYRIGHT'RUPADR!2+WIECIÊSKIEGO)33.
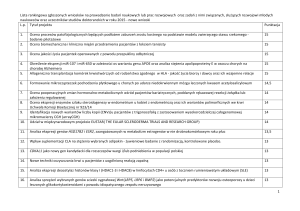
Tab. I. Ekspresja genów różnicujących związanych z kaskadą kaspaz w komórkach białaczki promielocytarnej HL-60 eksponowanych na CPT względem prób odniesienia (komórki eksponowane na DMSO)
Nazwa sondy
Symbol genu
Nazwa kodowanego białka
205780_at
BIK
201747_s_at
SAFB
211833_s_at
BAX
213850_s_at
SFRS2IP
210506_at
FUT7
214484_s_at
SIGMAR1
nieopioidowy wewnątrzkomórkowy receptor sigma 1
(sigma non-opioid intracellular receptor 1)
+ 1,55 ↑
202844_s_at
RALBP1
białko wiążące ralA
(ralA binding protein 1)
+ 1,51 ↑
201831_s_at
USO1
homolog białka drożdży dokujący do pęcherzyka USO1
(USO1 vesicle docking protein homolog (yeast))
+ 1,34 ↑
209790_s_at
CASP6
203708_at
PDE4B
204118_at
CD48
202643_s_at
TNFAIP3
białko 3 indukowane czynnikiem martwicy nowotworów alfa
(tumor necrosis factor, alpha-induced protein 3)
- 1,79 ↓
202644_s_at
TNFAIP3
białko 3 indukowane czynnikiem martwicy nowotworów alfa
(tumor necrosis factor, alpha-induced protein 3)
- 2,07 ↓
białko z rodziny BCL-2,
promotor śmierci komórkowej
(BCL-2-interacting killer)
czynnik B wiążący chromatynę
(scaffold attachment factor B)
białko X związane z BCL-2
(BCL-2-associated X protein)
białko oddziałujące z bogatym w serynę/argininę czynnikiem
splicingowym 2
(serine/arginine-rich splicing factor 2, interacting protein)
fukozylotransferaza 7
(fucosyltransferase 7)
kaspaza 6
(caspase 6, apoptosis-related cysteine peptidase)
fosfodiesteraza cAMP 4B
(phosphodiesterase 4B, cAMP-specific)
antygen różnicowania komórkowego 48
(cluster of differentiation 48)
Wartość SLR
+ 1,72 ↑
+ 1,70 ↑
+ 1,66 ↑
+ 1,66 ↑
+ 1,58 ↑
+ 1,34 ↑
- 1,74 ↓
- 1,79 ↓
SLR – ang. signal log ratio – stosunek zlogarytmowanych uśrednionych wartości fluorescencji transkryptu w porównywanych próbach, strzałka ↑ – nadekspresja, strzałka ↓ – spadek ekspresji genu w komórkach HL-60 eksponowanych na CPT
CD48 i PDE4B. Szczegółowe dane uwzględniające nazwy sond, nazwy genów, funkcje tych genów oraz wartości liczbowe SLR zestawiono w tabeli I.
Dyskusja
Zastosowanie metody mikromacierzy oligonukleotydowych umożliwiło określenie wpływu CPT na profil ekspresji genów kodujących białka związane z kaskadą kaspaz
w komórkach białaczki promielocytarnej HL-60. Zaobserwowano, że genem o najbardziej stymulowanej ekspresji
był BIK, który koduje proapoptotyczne białko BIK. Białko
to zlokalizowane jest głównie w siateczce śródplazmatycznej (ER) i indukuje apoptozę za pośrednictwem szlaku mitochondrialnego poprzez mobilizację wapnia z ER do mitochondriów oraz przebudowę grzebieni wewnętrznej błony
mitochondrialnej. Białko BIK jest proapoptotyczym supresorem nowotworów w różnych ludzkich tkankach i jego
ekspresja w nowotworach jest zablokowana przez delecje
chromosomalne obejmujące locus Bik lub poprzez wyciszanie epigenetyczne. Białko BIK wydaje się być kluczowym
efektorem w apoptozie indukowanej toksynami, cytokinami
i infekcjami wirusowymi. Kilka leków przeciwnowotworowych aktywuje ekspresję genu BIK poprzez szlaki transkrypcyjne zależne od takich czynników transkrypcyjnych
jak E2F i białka P53 lub poprzez usunięcie zmian epigenetycznych w chromatynie [9]. Białko BIK, wraz z innymi
białkami proapoptotycznymi takimi jak BAD i BID, uczestniczy w modulowaniu funkcji apoptosomu. Białka te poprzez związanie się z białkami BCL-2/xL umożliwiają i ułatwiają aktywację kaspazy 9 przez Apaf-1 [10]. Kolejnym
genem różnicującym, którego ekspresja była stymulowana
w komórkach HL-60 jest SAFB. Jego produktem jest białko SAFB, którego plejotropowość wiąże się z obecnością
w jego strukturze licznych domen. Domena SAP obecna
na N-końcu jest charakterystyczna dla białek, które zaangażowane są w procesy takie jak: transkrypcja, przetwarzanie RNA, apoptotyczna degradacja chromatyny i naprawa
DNA. SAFB odgrywa niekorzystną rolę w rozwoju nowotworów piersi. Białko to funkcjonuje jako korepresor dla
receptorów estrogenowych. Nadekspresja SAFB powoduje
zahamowanie aktywności tych receptorów. Ponadto nade-
&ARM0RZEGL.AUK
kspresja SAFB w hodowlach komórkowych doprowadza do
zahamowania wzrostu i generuje komórki o wielu jądrach
[11]. Gen BAX, którego ekspresja również znacząco wzrosła, koduje białko należące do rodziny białek BCL-2, która
stanowi grupę onkogenów komórkowych regulujących proces śmierci komórki. Białka z rodziny BCL-2 mają wspólny co najmniej jeden z czterech regionów określanych jako
domeny homologii z BCL-2 (BCL-2 homology domainsBH): BH1, BH2, BH3 i BH4. Rodzinę białek BCL-2 można podzielić na trzy grupy, biorąc pod uwagę ich strukturę
oraz pełnione funkcje. Do I grupy należą białka antyapoptotyczne, takie jak BCL-2, MCL-1, BCL-xL, które praktycznie zawsze posiadają wszystkie 4 domeny BH. Do grupy II
klasyfikuje się białka proapoptotyczne, jak na przykład Bak,
Bax, i inne (tzw. „multidomain” proapoptotic proteins). Do
grupy III należą białka proapoptotyczne posiadające „tylko
domenę BH3” (tzw. „BH3-only” proteins), m.in. o białka
BIK, BAD, NIX, BLK, NIP3. Białko BAX w żywotnych
komórkach obecne jest w formie monomeru w cytozolu
albo luźno przylega do zewnętrznej błony mitochondrialnej.
Jednak w formie monomeru nie jest zdolne do tworzenia
kanałów w błonach lipidowych, ani do uwalniania cytochromu C z mitochondriów. Dopiero z chwilą wystąpienia
sygnałów indukujących przepuszczalność błony mitochondrialnej cytozolowy BAX oraz BAX znajdujący się przy
błonie mitochondrialnej ulegają homooligomeryzacji i jako
multimer są wprowadzane w zewnętrzną błonę mitochondrialną. Oligomeryzacja BAX jest możliwa dzięki działaniu
innego białka proapoptycznego, a mianowicie białka BID,
które wcześniej musi być rozszczepione przez kaspazę 8/10
do postaci tBID [12]. Wykazano, że zwiększona ekspresja
BAX w warunkach in vivo jest skorelowana w wielu typach
komórek z wrażliwością na chemioterapię [13]. Zaobserwowano również, że komórki linii komórkowej jelita DLD-1
transfekowanej genem BAX w większym stopniu ulegają
apoptozie pod wpływem cytostatyków – CPT i doksorubicyny w porównaniu z komórkami nietransfekowanymi [14].
Innym genem, którego ekspresja była znacząco stymulowana był gen CASP6, kodujący kaspazę 6, która obok kaspazy
3 i 7 jest jedną z trzech kaspaz efektorowych - końcowych
enzymów kaskady kaspaz. Aktywacja kaskady kaspaz doprowadza w efekcie do śmierci komórki przez rozkład jej
struktur, fragmentację DNA oraz inaktywację mechanizmów
naprawczych DNA [15]. Aktywna kaspaza 6 trawi laminę,
wchodzącą w skład strukturalnych blaszek jądrowych, co
powoduje rozszczelnienie błony jądrowej [6]. A zatem można stwierdzić, że zwiększona ekspresja genu CASP6 może
być odpowiedzialna za promowanie apoptozy w komórkach
nowotworowych linii HL-60. Na uwagę zasługuje również
gen RALBP1, którego ekspresja także była stymulowana
po ekspozycji na CPT. Gen ten koduje białko 1 wiążące
RAL (RLIP76), które jest białkiem błonowym transportującym na zewnątrz komórki koniugaty glutationu, związki
elektrofilowe i inne ksenobiotyki w tym chemioterapeutyki
[16]. Białko to często wykazuje nadekspresję w komórkach
nowotworowych i odgrywa w nich znaczącą rolę antyapoptotyczną [17]. Drake i wsp. [18] wykazali, że nadekspresja
RALBP1 w ludzkiej linii komórek nowotworowych białaczki szpikowej K562, jest odpowiedzialna za oporność min.
na CPT, doksorubicynę i mitomycynę-C. A zatem zwiększo-
na ekspresja RALBP1 w linii HL-60 prawdopodobnie jest
powiązana z opornością na CPT.
Wśród genów, których ekspresja została znacząco zahamowana po ekspozycji na CPT występuje TNFAIP3, określany również jako A20. Początkowo został on uznany za
gen, który jest indukowany przez czynnik martwicy nowotworu (TNF) w ludzkich komórkach śródbłonka żyły pępowinowej. Późniejsze badania wykazały, że A20 jest również
indukowany w wielu innych typach komórek przez szeroką gamę bodźców [19]. Produktem tego genu jest białko
cytoplazmatyczne zawierające motyw palca cynkowego,
który hamuje aktywność jądrowego czynnika kappa-B (NFκB), a także programowaną śmierć komórki przebiegającą
z udziałem TNF [20]. A zatem można założyć, że obniżona
ekspresja tego genu sprzyja apoptozie indukowanej przez
CPT.
Przedstawione wyniki badań sugerują, że CPT w znaczny sposób modyfikuje ekspresję genów związanych z kaskadą kaspaz w komórkach HL-60. Poznanie efektów towarzyszących zmianie ekspresji tych genów wydaje się być
niezwykle istotne do określenia mechanizmów działania
leków, przeciwdziałania oporności a także do wdrażania
innych niż chemioterapia podejść terapeutycznych jak np.
przeciwnowotworowa terapia genowa, która dzięki modulowaniu aktywności poszczególnych genów może stanowić
uzupełnienie standardowych metod leczenia.
Wnioski
1. CPT powoduje zmiany ekspresji genów związanych
z kaskadą kaspaz w komórkach nowotworowych białaczki
promielocytarnej HL-60.
2. Wśród genów związanych z kaskadą kaspaz różnicujących odpowiedź komórek HL-60 hodowli eksponowanej
na CPT w porównaniu do hodowli kontrolnej najbardziej
stymulowana była ekspresja genów BIK, SAFB, BAX, a najbardziej zahamowana TNFAIP3, CD48 i PDE4B.
3. Wyjaśnienie zmian ekspresji genów związanych z kaskadą kaspaz wywoływanych przez CPT może mieć istotne
znaczenie w ustalaniu schematu terapii przeciwnowotworowej z zastosowaniem tego leku i przewidywaniu jej skuteczności.
Piśmiennictwo
1. Kolenko VM i wsp. Caspase-dependent and – independent death pathways in cancer therapy. Apoptosis 2005;
1: 17-20.
2. Boulikas T. Molecular mechanisms of cisplatin and
its liposomally encapsulated form, LipoplatinTM. LipoplatinTM as a chemotherapy and antiangiogenesis drug.
Cancer Ther 2007; 5: 351-376.
3. Cepeda V i wsp. Biochemical mechanisms of cisplatin
cytotoxicity. Anti-Cancer Agents Med Chem 2007; 7:
3-18.
4. Korzeniewska-Dyl I. Kaspazy – struktura i funkcja. Pol
Merk Lek 2007; 23: 403-407.
5. Korzeniewska-Dyl I. Kaspazy – nowy cel w terapii
przeciwzapalnej i przeciwnowotworowej? Pol Merk
Lek 2008; 24: 5-7.
COPYRIGHT'RUPADR!2+WIECIÊSKIEGO)33.
6. Fang TJ i wsp. Caspase family proteases and apoptosis.
Acta Biochem Biophys Sin 2005; 37: 719-727.
7. Ślesak B, Harłozińska A, Knast W. Ekspresja kaspazy
3 (CPP32), Ki-67 w relacji do wczesnych faz apoptozy
w złośliwych i zapalnych guzach trzustki. Adv Clin Exp
Med 2005; 14: 225-229.
8. Hordyjewska A, Pasternak K. Apoptotyczna śmierć komórki. Adv Clin Exp Med 2005; 14: 545-554.
9. Chinnadurai G, Vijayalingam S, Rashmi. BIK, the founding member of the BH3-only family proteins: mechanisms of cell death and role in cancer and pathogenic
processes. Oncogene 2009; 27: 20-29.
10. Maślińska D. Programowana śmierć komórki (apoptoza) w procesie zapalnym. Nowa Med 1999; 12:
34-41.
11. Towson S i wsp. SAFB2, a new scaffold attachment factor homolog and estrogen receptor corepressor. J Biol
Chem 2003; 278: 20059-20068.
12. Rupniewska Z, Bojarska-Junak A. Apoptoza: Przepuszczalność błony mitochondrialnej i rola pełniona przez
białka z rodziny BCL-2. Postepy Hig Med Dosw 2004;
58: 538-547.
13. Øyan AM i wsp. Genes of cell-cell interactions, chemotherapy detoxification and apoptosis are induced during chemotherapy of acute myeloid leukemia. BMC Cancer 2009; 9: 77.
14. Rzońca S, Małecki M. Proapoptotyczna terapia genowa
a wrażliwość nowotworów na chemioterapię. Współcz
Onkol 2009; 13: 61-65.
15. Malinowska I. Rola apoptozy w patogenezie i leczeniu
nowotworów układu hematopoetycznego. Postepy Hig
Med Dosw 2004; 58: 548-559.
16. Awasthi S i wsp. RLIP76 and Cancer. Clin Cancer Res
2008; 14: 4372-4377.
17. Singhal SS i wsp. Regession of lung and colon cancer
xenografts by depleting or inhibiting RLIP76 (Ral-Binding Protein 1). Cancer Res 2007; 67: 4382-4389.
18. Drake KJ i wsp. Multidrug-resistance conferred by
RalBP1/RLIP76. Int J Oncol 2007; 30: 139-144.
19. Coornaert B, Carpentier I, Beyaert R. A20: Central
gatekeeper in inflammation and immunity. Biol Chem
2009; 284: 8217-8221.
20. Lee EG i wsp. Failure to regulate TNF-induced NFkappaB and cell death responses in A20-deficient mice.
Science 2000; 289: 2350-2354.
data otrzymania pracy: 27.07.2010 r.
data akceptacji do druku: 27.08.2010 r.
Adres do korespondencji:
mgr Ewelina Aleksander
Katedra i Zakład Biofarmacji
Śląski Uniwersytet Medyczny w Katowicach
ul. Narcyzów 1
41-200 Sosnowiec
e-mail: [email protected]