Egzamin
Egzamin pisemny z Neurobiologii odbędzie we wtorek 12 czerwca o godz. 10 w Sali
132.
Po egzaminie pisemnym, będzie można się zapisać na egzamin ustny, w terminie 13-15
czerwca.
Część wisceralna i somatyczna
Ciało składa się z dwóch części:
Część wisceralna (trzewia): organy wewnętrzne – narządy klatki piersiowej (serce,
płuca) i jamy brzusznej (żołądek, jelita).
Część somatyczna – aparat mięśnioszkieletowy
A. Hipotetyczny prymitywny strunowiec z rozdzieloną częścią wisceralną i somatyczną.B. Niższy
kręgowiec (ryba) wykazujący większą integrację dwóch składowych ciała. Z: Romer, A. S. 1964. The
Vertebrate Body. W. B. Saunders. Philadelphia.
Układ autonomiczny i somatyczny
Układ autonomiczny (wegetatywny) unerwia narządy wewnętrzne. Działanie u.a.
powoduje reakcje niezależnie od naszej woli (np. wydzielanie soków żołądkowych) i
utrzymuje podstawowe parametry fizjologiczne (temeperatura, ciśnienie krwi) na
poziomie dostosowanym do aktualnego zachowania i warunków środowiska.
Układ somatyczny – kieruje pracą mięśni szkieletowych, gruczołów skórnych i komórek
barwnikowych skóry. W dużym stopniu podlega kontroli świadomości.
Podział układu nerwowego
Aktywność ruchowa
System ruchowy generuje:
- odruchy
- ruchy rytmiczne
- ruchy zamierzone
Odruchy
Charles Sherrington – ok. 1890. Odruch - podstawowa jednostka
funkcjonalna rdzenia kręgowego oraz innych części układu nerwowego.
Łuk odruchowy – droga impulsu nerwowego od receptora do efektora
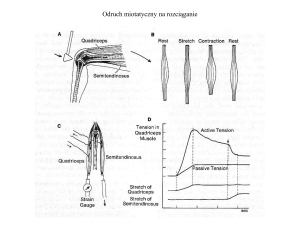
Odruchy rdzeniowe - odruch miotatyczny na rozciąganie
Odpowiedzi mięśnia
czworogłowego uda na rozciąganie
przed i po przecięciu
unerwiającego go motoneuronu.
Po przecięciu nerwu, odpowiedź
jest mniejsza, co wskazuje na
istnienie polaczen przechodzacych
przez rdzen kregowy, a nie na
wrodzoną reakcję mięśnia.
Obwody rdzenia powodują skurcz
męśnia, ktory byl rozciagniety.
Rozciaganie mięśnia
przeciwstawnego (mięsień
półścięgnisty), powoduje reakcje
hamującą (strzałka).
Obwody rdzenia kręgowego
Receptory w skórze i mięśniach wysyłają informacje do rdzenia kręgowego. Ciałka komórek receptorowych
znajdują się w zwojach korzeni grzbietowych (dorsal root ganglion). Włóka wchodzą do rdzenia kręgowego
poprzez korzenie grzbietowe (dorsal root). Komórki rogów brzusznych (ventral horn) wysyłają połączenia przez
korzenie brzuszne (ventral root) do mięśni, w celu kontroli ruchu.
Odruchy mięśniowe i skórne
Badanie obwodów odruchów. Porównanie odpowiedzi w aksonie motoneuronu w rdzeniu kręgowym na
stymulację elektryczną włókna wrzecionka mięśniowego (włókna przewodzące szybkie) i włókna
czuciowego skóry (włókna przewodzące wolne + droga polisynaptyczna).
Rodzaje włókien nerwowych
Obwody refleksów - połączenia monosynaptycze i
polisynaptyczne
neuron pobudzający
neuron hamujący
A. Badanie obwodów odruchów. B. Porównanie odpowiedzi motoneuronu w rdzeniu kręgowym na stymulację
wychodzących włókien mięśniowych typu Ia i II. Eksperymentalne wykazanie drogi monosynaptycznej i
dwusynaptycznej do motoneuronu u kota.
Wnioski:
- aferenty Ia tworzą drogę pobudzającą monosynaptyczną z własnymi motoneuronami i
dwusynaptyczną hamującą droge z motoneuronami antagonistycznymi
- aferenty II tworzą dwusynaptyczną pobudzającą drogę z własnymi motorneuronami.
Odruchy rdzeniowe
Odruchy rdzeniowe i wyższe
Bodźce czuciowe wyzwalają refleksy poprzez obwody rdzenia i dłuższe obwody wyższe.
Krótkie napięcie mięśnia kciuka powoduje szybką odpowiedź M1 w rozciąganym mięśniu i
wolniejszą odpowiedź M2 biorącą się z obwodu zawierającego korę motoryczną.
Koordynacja odruchów
Hamujące interneurony koordynują odruchy.
A. Ia interneuron umożliwia koordynację mięśnia przeciwstawnego pojedynczym sygnałem. Dodatkowo, otrzymuje
on wyjścia z obwodów wyższych dzięki czemu sygnał zstępujący aktywujący jedną grupę mięśni, automatycznie
rozluźnia grupę przeciwstawną. Inne wejścia zstępujące tworzą połączenia hamujące i pobudzające, zwiększając
możliwości kontroli.
B. Komórki Renshawa tworzą hamowanie zwrotne z motoneuronami. Umożliwia to regulację pobudzenia
motoneronów i stabilizację ich częstości odpalania. Wejścia zstępujące modulujące pobudzenie komórek Renshawa
modulują pobudliwość wszystkich motoneuronów w stawie.
Lokomocja
Eadweard Muybridge i zoopraxiskop (1879)
Neuronalne składowe systemów motorycznych
Główne komponenty większości systemów motorycznych: mięśnie, oscylator (aktywność rytmiczna) i ośrodki kontroli ruchu
Centralne generatory wzorca
Centralny generator wzorca – mechanizm neuronalny generacji rytmicznych i
skoordynowanych impulsów do motoneuronów
Podstawowe typu generatorów rytmu. Oznaczenia: D – driver, E – extensor motoneuron, F – flexor motoneuron, I –
interneuron, P – pacemaker (generator rytmu).
Pływanie u kręgowców - minóg
Minóg w trakcie pływania wykonuje falujące ruchy. Skurcz
po prawej stronie powoduje wygięcie ciała w prawo, po
lewej – w lewo. Fala skurczu rozchodzi się od głowy do
ogona po obu stronach ciała. Fale po każdej stronie są
względem siebie przesunięte w fazie.
Układ pływania u minoga składa się z wyższych ośrodków w pniu mózgu i segmentów w części rdzeniowej. Każdy segment tworzy
CGW. Podczas pływania sieci CGW są aktywowane przez system siatkowo-rdzeniowy (reticulospinal system). Najbardziej pobudliwe
generatory wzorca znajdują się w przednim odcinku, co powoduje rozchodzenie się skurczu od przodu do tyłu. Połączenie pomiędzy
segmentami zapewniają koordynację i opóźnienia (lag) pomiędzy kolejnymi CGW. Opóźnienie determinuje prędkość ruchu.
Segmentowy obwód CGW u minoga
MN – motoneurony, interneurony pobudzające (E), interneurony hamujące (L, I), pień mózgu pobudza
wszystkie te elementy. Aktywność w każdym segmencie jest inicjowana poprzez pobudzenie z systemu
siatkowo-rdzeniowego. Pobudzenie receptora NMDA powoduje aktywacja prądów Na+/Ca2+. Napływ
Ca2+ wywołuje repolaryzacje poprzez IK(Ca). Pobudzenie receptora kainate/AMPA przyśpiesza cykl.
Neurony pobudzające (E) pobudzają motoneurony oraz aktywują interneurony hamujące (L, I). Aksony
interneuronów I hamują aktywność wszystkich komórek w kontralateralnej stronie segmentu, zapewniając
przesuniecie w fazie aktywności po obu stronach o 180 o.
Od pływania do chodzenia
Pływanie ryby
Chodzenie salamandry.
Nogi u płazów wyewoluowały z płetw u ryb. Poruszanie po lądzie odbywa się poprzez wyciągnięcie,
podparcie i nacisk kończyn na podłoże w koordynacji z pływającymi ruchami ciała.
..i biegania u jaszczurki
U płazów i gadów nogi zamocowane są bocznie (stabilność). U ptaków i ssaków – od dołu (prędkość). Niektóre
gady mogą poruszać się bardzo szybko. Bazyliszek (basilisk lizard) osiąga prędkość 20 km/h (krok dwunożny)
Kroki
Porównanie kroków karalucha i kota.
Stopa uniesiona
Stopa na ziemi
Kroki cd
Cykl kroku kota składa się z dwóch faz – zamachu i podparcia. Dolne przebiegi pokazują
aktywność mięśni.
Rdzeniowe mechanizmy chodzenia
Zapis elektromiogramu (EMG) z mięśni kota w stanie normalnym (A) i po przecięciu połączeń z mózgiem
(B). Wzorzec podstawowy jest zachowany lecz staje się mniej stabilny i łatwo ustaje.
Rdzeniowe mechanizmy chodzenia
Centralny generator wzorca generujący chodzenie u kota jest typu ‘half-center’ i zlokalizowany jest w rdzeniu. Długie,
opóźnione serie wyładowań są widoczne w interneuronach w rdzeniu kręgowym w wyniku stymulacja ipsilateralnej
nerwów FRA (flexor reflex afferents).
Hierarchiczna struktura organizacji kontroli ruchu - początki
Piętra kontroli ruchu wg. Jacksona:
Płat czołowy
John Hughlings Jackson (1835 - 1911)
Na podstawie obserwacji pacjentów
epileptycznych opracował koncepcję
poziomów kontroli ruchów. Niższe
poziomy kontrolują ruchy automatyczne,
wyższe – ruchy zamierzone. Wyższe
poziomy kontrolują też niższe poziomy
poprzez hamowanie lub pobudzanie.
Gdy wyższa funkcja jest uszkodzona,
niższe poziomy ulegają ‘wyzwoleniu’
spod kontroli, co prowadzi do
hyperaktywności.
Kora mózgowa wzdłuż bruzdy Rolanda
Rdzeń kręgowy i pień mózgu
Hierarchiczna struktura organizacji kontroli ruchu
Układy motoryczne mają trzy poziomy
kontroli – rdzeń kręgowy, pień mózgu i
kora mózgowa. Wszystkie trzy poziomy
układu motorycznego dostają wejścia
sensoryczne i są dodatkowo pod
wpływem dwóch struktur podkorowych –
zwojów podstawy i móżdżku.
Hierarchiczna struktura organizacji kontroli ruchu
1.
Aparat rdzeniowy kontrolowany
poprzez motoryczne obszary kory i
pnia mózgu (poziom środkowy
Jacksona).
2.
Kora i pień mózgu kontrolowane
poprzez różne obszary (np.
móżdżek, zwoje podstawy), a nie
tylko przez korę przednią.
3.
Kontrola nie jest stricte
hierarchiczna (np. drogi równoległe,
nie tylko szeregowe).
4.
Połączenia zwrotne, również do
najwyższego poziomu.
Dwa rodzaje kontroli
A.
W układzie z kontrolą zwrotną,
sygnał z sensora jest porównywany
z sygnałem odniesienia. Różnica jest
przekazywana do kontrolera i
wywołuje odpowiednią reakcje
mięśni.
B.
Kontrola torująca opiera się na
informacji otrzymanej przed
aktywacją sensora kontroli zwrotnej.
Mechanizm ma zasadnicze
znaczenie dla ruchów szybkich.
Dwa rodzaje kontroli
A.
Układ eksperymentalny łapania
piłki.
B.
Średnia odpowiedź człowieka
łapiącego piłkę puszczoną z 0.8 m.
Zapisy od góry do dołu pokazują:
kąt łokcia, kąt nadgarstka, zapis
EMG z bicepsa, tricepsa, flexor
carpi radialis (FCR)i extensor carpi
radialis (ECR). Widoczna aktywacja
mięśni zarówno przed, jak i po,
złapaniu piłki.
Kontrola ruchu – ośrodki pnia mózgu
Przyśrodkowe i boczne drogi zstępujące z pnia mózgu kontrolują różne grupy neuronów i mięśni. A. Drogi przyśrodkowe
dochodzą do części brzusznej rdzenia kręgowego i kontrolują mięśnie tułowia (postura). B. Droga boczna zaczyna się w
wielkokomórkowej części jądra czerwiennego i opada w kontralateralnej kolumnie grzbietowej a następnie dochodzi do
części grzbietowej rdzenia kręgowego unerwiając motoneurony dystalnych (dalszych) części kończyn (manipulacja).
Ośrodki pnia mózgu inicjują lokomocję i kontrolują jej
prędkość
Wzrost pobudzenia elektrycznego przyłożonego do
mesencephalic locomotor region (MLR, część pnia mózgu) u
kotów z przeciętym połączeniem do kory mózgowej, zmienia
krok i prędkość chodu. W miarę wzrostu prędkości, tylne łapy
zmieniają krok z naprzemiennego na równoczesne zgięcia i
wyprosty.
Kontrola chodzenia - trzy ośrodki pnia mózgu
Aktywność neuronów w trzech ośrodkach pnia mózgu podczas chodzenia. A.
Komórki siatkowato – rdzeniowe zwiększają odpalanie w fazie zginania.
Usunięcie móżdżku znacznie redukuje tę aktywność – spada napięcie mięśni
koordynacja kończyn. B. Aktywność neuronów przedsionkowo-rdzeniowych
osiąga maksimum w fazie podparcia. C. Aktywność neuronów czerwienno rdzeniowych osiąga maksimum w fazie zgięcia. Wniosek: ośrodki pnia mózgu
kontrolują generatory rdzenia kręgowego poprzez i) ogólną aktywację i ii)
modulowaną aktywność torującą poszczególne fazy kroku
Móżdżek
- ma 10-100 * 109 neuronów (więcej niż
wszystkie pozostałe części mózgu razem
wzięte)
- liczba połączeń z korą mózgową – 40*106
(trakt optyczny - 1* 106)
- budowa modułowa (wykonywanie tych
samych operacji na różnych wejściach)
Móżdżek kontroluje układ ruchowy porównując polecenia ruchowe z ich rzeczywistym wykonaniem i
korygując komendy w korze mózgowej i pniu mózgu podczas wykonywania oraz powtarzania ruchu.
Trzy aspekty organizacji służą tym funkcjom: 1. Duża liczba wejść (40 razy więcej niż wyjść). 2.
Wyjścia do kory motorycznej i pnia mózgu – struktur kontrolujących ruch. 3. Transmisja synaptyczna w
obwodach móżdżku może być modyfikowana, co umożliwia adaptacje motoryczną i uczenie.
Neurony kory móżdżku
Kora móżdżku ma 3 warstwy i 5
rodzajów komórek. Warstwa
drobinowa (molecular layer),
zwojowa (Purkinje cell body
layer), ziarnista (granule layer).
Komórki Purkinjego, ziarniste
(Granule), gwiaździste (Stellate),
koszyczkowe (Basket), Golgiego.
Móżdżek otrzymuje dwa rodzaje
wejść: mossy fibers i climbing
fibers. Oba wejścia są
pobudzające lecz tworzą różne
wzorce wyładowań w komórkach
Purkiniego.
Kora móżdżku – wejścia i wyjścia
Mossy fibers pobudzają komórki
ziarniste, które włóknami
równoległymi pobudzają kilkaset
komórek Purkinjego. Climbing fibers
tworzą połączenia w kierunku
prostopadłym z ok. 10 komórkami
Purkinjego. Połączenia wejściowe
tworzą więc macierz ortogonalną.
Obwód móżdżku
Podstawowe cechy obwodu:
1.
Oba wejścia, bezpośrednie
(climbing fibers CF) i pośrednie
(mossy fibers MF - k. ziarniste –
włókna równoległe) są
pobudzające.
2.
Jądra głębokie również dostają
wejścia z CF i MF.
3.
Wszystkie inne połączenia są
hamujące.
4.
Wyjściowa pętla pobudzająca przez
jądra głębokie jest modulowana
przez pętlę hamującą kory
móżdżku (regulacja ruchów w
czasie rzeczywistym).
Aktywność w komórkach Purkinjego
Oba wejścia pobudzające tworzą różne
wzorce wyładowań w komórkach
Purkinjego. Wyładowania złożone
(prawa klamra) są wywołane poprzez
synapsy climbing fiber, a pojedyncze
spiki (lewa klamra) są wywołane
wejściem z mossy fiber. Mossy i
climbing fiber różnie kodują sygnały
sensoryczne. Częstość pojedynczych
spików w komórkach Purkinjego zależy
od aktywności we włóknach
sensorycznych oraz od aktywności
ruchowej. Częstość spików koduje więc
czas trwania i natężenie ruchu.
Wyładowania w climbing fibers są
rzadkie i kodują prawdopodobnie
czasowe zależności w sygnałach
przychodzących oraz mogą być
‘wyzwalaczem’ aktywności.
Plastyczność w móżdżku
Uczenie w móżdżku – długotrwałe osłabienie synaptyczne (long-term depression, LTD) w
synapsach włókien równoległych w wyniku pobudzania komórek Purkinjego poprzez włókna pnące
(climbing fibers). Osłabienie może się utrzymywać minuty lub godziny i zależy od depolaryzacji
wywołanej aktywnością włókien pnących.
Mechanizm: Aktywacja włókien
pnących prowadzi do zwiększenia
stężenia Ca2+ w komórkach Purkinjego.
Wapń uruchamia mechanizm
przekaźnictwa wtórnego, który obniża
czułość receptorów glutaminergicznych
AMPA w synapsach włókien
równoległych
Uczenie się ruchów
Włókna pnące przekazują informację o błędzie motorycznym z jądra oliwki do komórek Purkinjego.
Ponowne pojawienie się tej samej informacji wejściowej powoduje mniejsze pobudzenie (LTD) komórek
Purkinjego przez włókna równoległe . Kolejne próby wykonania danego zadania zmieniają wyjście z
komórek Purkinjego tak, że wykonanie ruchu ulega poprawie.
Uczenie w móżdżku na przykładzie adaptacji koordynacji
wzrokowo-ruchowej
A, B. W wyniku okularów
zakrzywiających widzenie, wzrok widzi
cel z lewej strony. Początkowo ręka
kieruje lotkę w stronę widzianego celu.
Następnie następuje korekta rzutu w
prawo wg doświadczenia a nie patrzenia,
tak, że wykonanie zadania ulega
poprawie.
C. Adaptacja nie występuje u pacjenta z
jednostronnym uszkodzeniem móżdżku.
Defekty wywołane uszkodzeniami móżdżku
Uszkodzenia móżdżku prowadzą do zaburzeń
koordynacji ruchowej.
•
Uszkodzenie prawej półkuli móżdżku
opóźnia wykonanie ruchu (zaciśnięcie
obu dłoni na sygnał ‘go’). Ruch lewą
ręką jest opóźniony.
•
Pacjent ma kłopoty w sterowaniu
ruchem, by dotknąć czubek nosa. Tremor
zwiększa się wraz ze zbliżaniem się do
nosa.
•
Niemożność wykonania regularnych
ruchów - zginanie i prostowanie
przedramienia w łokciu.