Spis treści
1 Błony Biologiczne
1.1 Struktura i skład chemiczny
1.1.1 Szczegółowa budowa błony biologicznej
1.2 Właściwości i pełnione funkcje
1.2.1 Właściwości błon biologicznych
1.2.2 Funkcje błon biologicznych
1.3 Typy transportu przez błonę komórkową
1.3.1 Transport małych cząstek
1.3.2 Transport większych cząstek
2 Cytoplazma
2.1 Rola
2.2 Skład i właściwości
2.3 Ruchy cytoplazmy
3 Jądro komórkowe
3.1 Funkcje
3.2 Budowa
3.2.1 Otoczka i pory jądrowe
3.2.2 Blaszka jądrowa
3.2.3 Chromosomy
3.2.4 Jąderko
3.2.5 Macierz jądrowa
3.3 Stany jądra komórkowego
4 Mitochondrium
4.1 Budowa
4.2 Funkcje
5 Organelle otoczone pojedynczą błoną komórkową
5.1 Reticulum endoplazmatyczne
5.1.1 Typy
5.2 Rybosomy
5.2.1 Typy
5.3 Funkcje
5.4 Aparat Golgiego
5.5 Budowa
5.6 Funkcje
5.7 Lizosomy
5.7.1 Rodzaje lizosomów
6 Wakuole
6.1 Funkcje
6.2 Wakuolę wypełniają
6.3 Plazmoliza
6.4 Deplazmoliza
7 Ściana komórkowa
7.1 Funkcje
7.2 Budowa
7.3 Formowanie się ściany komórkowej
Błony Biologiczne
Błona biologiczna jest to otoczka rozdzielająca odrębne przedziały w komórkach — jest ona
podstawową strukturą budującą komórki wszystkich organizmów. Są to zarówno błony komórkowe
jak i błony organelli wewnętrznych, których ogólna budowa we wszystkich organizmach jest taka
sama. U eukariontów posiadających ścianę komórkową zawsze występuje po stronie wewnętrznej tej
ściany.
Struktura i skład chemiczny
Przekrój przez błony eukariontów
Błony biologiczne składają się zawsze z dwóch komponentów: fosfolipidów (do których należą
fosfolipidy, glikolipidy i steroidy; tworzących zrąb lipidowy) oraz białek.
Na przestrzeni lat powstało kilka teorii budowy błony komórkowej. Należą do nich:
1. Model lipidowy (1895, Overton) — opierając się na fakcie, iż cząsteczki polarne znacznie
wolniej przenikają do wnętrza komórki niż cząsteczki niepolarne uznano, że błona komórkowa
zbudowana jest z lipidów.
2. Model dwuwarstwy lipidowej (1925, Gortel i Grendel) — podsumowując wyniki badań
dotyczących zawartości lipidów w erytrocytach sformułowano hipotezę, że błona komórkowa
składa się z dwóch warstw lipidowych, a polarne główki cząsteczek lipidów muszą być
skierowane na zewnątrz, a niepolarne łańcuchy węglowodorowe do wnętrza podwójnej
warstwy lipidowej.
3. Model trójwarstwowej błony (1935, Dowson i Danielli) — błony komórkowe zbudowane są
symetrycznie z podwójnej warstwy lipidowej pokrytej po obu stronach warstwą białek
(wyposażającą błonę w pewien stopień elastyczności i odporności mechanicznej oraz nadającą
jej niskie napięcie powierzchniowe). Drobiny lipidowe są zorientowane równolegle do siebie i
pod kątem prostym do płaszczyzny błony (niepolarne — nie naładowane — końce cząsteczek
lipidów są kierowane do siebie, a polarne — naładowane — skierowane są na powierzchnię
błony). Model ten zdawały się potwierdzać zdjęcia z mikroskopu elektronowego.
4. Model płynnej mozaiki (1972, Singer i Nicolson) — "w morzu lipidów pływają góry lodowe
białek..." — w modelu tym błony biologiczne są dwuwymiarowymi roztworami przestrzennie
zorientowanych lipidów i sferycznych białek (białka nie tworzą warstwy na powierzchni
lipidów, lecz pływają zanurzone w dwuwarstwie lipidowej). Dwuwarstwa jest
rozpuszczalnikiem dla białek błonowych i stanowi barierę przepuszczalności. W
monowarstwach istnieją tzw. „rafty” czyli lokalne obszary o składzie odbiegającym od rozkładu
przypadkowego (bogatsze od sąsiednich obszarów monowarstwy w specyficzne lipidy,
cholesterol czy białka). Błona taka jest asymetryczna, płynna i dynamiczna. Model ten, z
kilkoma modyfikacjami, obowiązuje do dnia dzisiejszego.
Szczegółowa budowa błony biologicznej
Lipidy błonowe:
Fosfolipidy— związki polarne (hydrofilowe "główki" — dzie znajduje się fosforan —
skierowane na zewnątrz błony — mające powinowactwo do wody oraz hydrofobowy
"ogon" skierowany do wnętrza — nie mający powinowactwa do wody, złożony z dwóch
łańcuchów kwasu tłuszczowego). Cząsteczki fosfolipidów mają w przybliżeniu jednakową
szerokość, co sprzyja układaniu się ich w podwójne warstwy cylindrycznych struktur.
Fosfolipidy łatwo przemieszczają się w obrębie jednej warstwy lipidowej błony (tzw.
dyfuzja boczna, która zachodzi co około 10—6 s). Wymiana cząsteczek lipidów między
jedną i drugą warstwą jest bardzo wolna (zachodzi nawet raz na kilkaset godzin).
Charakterystyczną cechą fosfolipidów jest to, że oprócz reszt kwasów tłuszczowych
występują w nich również reszty kwasu fosforowego (V). Wyróżnia się:
fosfolipidy cholinowe: fosfatydylocholina (lecytyna),
fosfolipidy aminowe: fosfatydyloetanolamina, fosfatydyloinozytol,
fosfatydyloseryna.
Glikolipidy — zlokalizowane w zewnętrznej warstwie błony; zawierają w swoim składzie
glicerol lub sfingozynę oraz kwasy tłuszczowe oraz składnik cukrowy (galaktoza lub
laktoza). Domeny polarne glikolipidów wystają ponad powierzchnię błony komórkowej,
prezentując swoje grupy polarne do środowiska. Warstwa glikolipidów pokrywa
większość komórek zwierzęcych tworząc tzw. glikokaliks. Wyróżnia się:
sfingoglikolipidy,
glikolipidy obojętne,
glikolipidy kwaśne.
Sterole — w błonie lokalizują się pomiędzy łańcuchami węglowodorowymi fosfolipidów:
fitosterole — pochodzenia roślinnego; do tej grupy należą m. in. β-sitosterol,
stigmasterol, kampesterol ,
zoosterole — pochodzenia zwierzęcego; do nich należy m. in. cholesterol.
Białka — biorą czynny udział w procesach takich jak transport, aktywność enzymatyczna,
przyczep komórek i komunikacja międzykomórkowa. Wyróżnia się:
Białka integralne — na trwałe wbudowane w dwuwarstwę; posiadają w łańcuchu
polipeptydowym przynajmniej jedną sekwencję składającą się z co najmniej 22
aminokwasów hydrofobowych, które pozwalają na zakotwiczenie się w błonie; do
ekstrahowania używa się detergentów — solubilizacja detergentem — przeprowadzenie
do roztworu wodnego kompleksów detergentu i składników błony). Wyróżnia się:
białka transbłonowe — przebijają całą grubość dwuwarstwy,
białka nie przebijające błony.
Białka powierzchniowe (peryferyjne) — leżą na powierzchni błony; są luźno związane z
powierzchniami błony; nie perforują one żadnej z monowarstw błony, a z błoną związane
są za pomocą słabych oddziaływań molekularnych, głównie wiązań jonowych,
wodorowych i Van der Waalsa; dają się łatwo usunąć z błony wodą lub roztworami soli.
Schemat organizacji białek błonowych. 1. Białko
transmembranowe 2. Białko monowarstwy
zewnętrznej 3. Białko monowarstwy wewnętrznej 4.
Białko wewnętrzne błony Niebieskie — białka
peryferyjne
W błonie komórkowej obserwuje się asymetrię dwuwarstwy. Wyróżnia się dwie warstwy:
1. lipidową zewnętrzną E (ang. exoplasmic) od strony środowiska,
2. lipidową cytoplazmatyczną P (ang. protoplasmic) od strony protoplazmy.
Na taką asymetrię wskazują:
różnice w budowie obu powierzchni błony, skierowanych na zewnątrz i ku wnętrzu komórki lub
organelli,
różnice w szybkości ruchów międzybłonowych pomiędzy poszczególnymi błonami,
różny skład fosfolipidów (w błonie erytrocytu człowieka warstwa E zbudowana jest głównie z
fosfolipidów cholinowych (fosfatydylocholin = lecytyn i sfingomielin), natomiast warstwa P
zbudowana jest z fosfolipidów aminowych tzw. kefalin: fosfatydyloseryny i
fosfatydyloetanoloaminy),
asymetryczne rozmieszczenie cholesterolu. Jest charakterystyczny dla zewnętrznej części
błony komórkowej (ta monowarstwa jest znacznie sztywniejsza),
duża ilość ujemnej fosfatydyloseryny w warstwie cytozolowej, wpływająca na ładunek ujemny
wewnątrz komórki.
Właściwości i pełnione funkcje
Właściwości błon biologicznych
Schematyczna kinematyka błony
lecytynowej
Dynamiczność — ruchy białek i lipidów (powodują m. in. zamykanie wszelkich wyrw i
ubytków):
fosfolipidy — okolice polarne mniej ruchliwe; końce łańcuchów węglowodorowych
wykonują szybkie ruchy,
białka — przemieszczane dyfuzyjnie w płaszczyźnie; wykonują ruchy obrotowe w osi
prostopadłej do powierzchni błony; także wynurzają się i zanurzają w dwuwarstwie
lipidowej.
Półprzepuszczalność — błony są w stanie przepuszczać niektóre rodzaje cząsteczek a
zatrzymywać inne; w szczególności woda przechodzi przez błonę swobodnie, a wybiórczo
substancje w niej zawarte. Przepuszczalność błony dla danej substancji zależy od rozmiaru i
ładunku jej cząsteczki. Cząsteczki większe takie jak na przykład glukoza i jony różnej wielkości
nie przedostają się z powodu zbyt dużych rozmiarów lub na skutek odpychania przez ujemnie
naładowaną powierzchnię błony.
Polarność — ładunki dodatnie na zewnątrz, ładunki ujemne od wewnątrz (istotne znaczenie
przy odbieraniu i przewodzeniu bodźców).
Funkcje błon biologicznych
Stanowią granicę pomiędzy światem zewnętrznym a światem wewnętrznym komórki lub
organellum — co jest podstawą do zachowania jego odrębności i integralności:
pozwalają na utrzymanie homeostazy komórki oraz utrzymanie odpowiedniego
środowiska wewnętrznego,
organizują komórkę i jej wnętrze (m. in. budują organella komórkowe); budują struktury
błoniaste: endoplazmatyczne retikulum, aparat Golgiego, pojedyncza błona otacza
wakuolę, lizosomy, peroksysomy a podwójna jądro komórkowe, mitochondria i plastydy.
Umożliwiają odbieranie i przewodzenie bodźców, pobieranie i wydalanie substancji i cząstek.
Umożliwiają oddziaływanie między komórką i podłożem oraz między komórkami.
Umożliwiają transport (na drodze dyfuzji, dyfuzji ułatwionej, transportu aktywnego oraz
endocytoz — pobieraniu makrocząsteczek do komórki i egzocytoz — wydzielaniu produktów
komórki do środowiska).
W błonach odbywają się niektóre procesy biochemiczne jak: fosforylacja w fotosyntezie,
łańcuch oddechowy w oddychaniu tlenowym.
Wytwarzają potencjał elektrochemiczny — różna koncentracja jonów.
U części protistów jak i niektórych komórek zwierzęcych (np. amebocyty gąbek) przelewanie
cytoplazmy powodujące uwypuklanie błony umożliwia przemieszczanie się tych komórek
(ruchem pełzakowatym — ameboidalny).
Typy transportu przez błonę komórkową
Zachowanie komórki roślinnej znajdującej się w
roztworze hiper-, izo- i hipotonicznym
Błąd przy generowaniu miniatury
(process:16563): Gtk-WARNING **: Locale not
supported by C library.
Using the fallback 'C' locale.
Background RRGGBBAA: ffffff00
Area 0:0:553.065:290.307 exported to 370 x 194 pixels
(60.2099 dpi)
Bitmap saved as:
/var/www/html/edu/images/tmp/transform_d61f2e71bf
21-1.png
terminate called after throwing an instance of
'std::bad_alloc'
what(): std::bad_alloc
Emergency save activated!
Emergency save completed. Inkscape will close now.
If you can reproduce this crash, please file a bug at
www.inkscape.org
with a detailed description of the steps leading to the
crash, so we can fix it.
GLib: Cannot convert message: Could not open
converter from 'UTF-8' to 'ANSI_X3.4-1968'
** Message: Error: Inkscape encountered an internal
error and will close now.
/var/www/html/edu/includes/limit.sh: line 101: 16562
Aborted /usr/bin/timeout $MW_WALL_CLOCK_LIMIT
/bin/bash -c "$1" 3>&Zachowanie komórki zwierzęcej (erytrocytu)
znajdującej się w roztworze hiper-, izo- i hipotonicznym
Błąd przy generowaniu miniatury
(process:16567): Gtk-WARNING **: Locale not
supported by C library.
Using the fallback 'C' locale.
Background RRGGBBAA: ffffff00
Area 0:0:581:254 exported to 370 x 162 pixels (57.315
dpi)
Bitmap saved as:
/var/www/html/edu/images/tmp/transform_1fb2e90a33
52-1.png
terminate called after throwing an instance of
'std::bad_alloc'
what(): std::bad_alloc
Emergency save activated!
Emergency save completed. Inkscape will close now.
If you can reproduce this crash, please file a bug at
www.inkscape.org
with a detailed description of the steps leading to the
crash, so we can fix it.
GLib: Cannot convert message: Could not open
converter from 'UTF-8' to 'ANSI_X3.4-1968'
** Message: Error: Inkscape encountered an internal
error and will close now.
/var/www/html/edu/includes/limit.sh: line 101: 16566
Aborted /usr/bin/timeout $MW_WALL_CLOCK_LIMIT
/bin/bash -c "$1" 3>&Schemat dyfuzji ułatwionej
Sposób transportu cząstki z i do komórki lub organellum zależy od jej rozmiaru i właściwości.
Transport małych cząstek
Dyfuzja — swobodne przenikanie cząsteczek przez podwójną warstwę lipidową z obszaru o
stężeniu wyższym do obszaru o stężeniu niższym (zgodnie z gradientem stężeń):
osmoza — transport wody lub innego rozpuszczalnika przez błonę; spontanicznie
zachodzi od roztworu o niższym stężeniu substancji rozpuszczonej do roztworu o
wyższym, czyli prowadzi do wyrównania stężeń obu roztworów; roztwór z którego ubywa
rozpuszczalnika nazywa się hipotonicznym, tego w którym przybywa nazywa się
hipertonicznym (gdy roztwory pozostają w równowadze osmotycznej, mówi się że są
wzajemnie izotoniczne względem siebie),
dializa — transport substancji rozpuszczonych przez błonę.
Dyfuzja ułatwiona — niektóre cząsteczki, np. glukoza, potrzebują nośników białkowych
(wiążących czasowo transportowaną cząstkę), co zwiększa tempo ich przedostawania się przez
błony; ruch cząsteczek odbywa się tylko w kierunku zgodnym ze spadkiem gradientu stężenia;
białko przenośnikowe po odłączeniu jednej cząsteczki może natychmiast wiązać się z drugą
(nie ulega przemianie). Transport nośnikowy może być związany z ruchem kompleksów
cząsteczka-nośnik w poprzek błony. Możliwa jest także sytuacja, w której nośnik wiąże
substancję transportowaną po jednej stronie błony, zmienia konformację i następnie uwalnia
przeniesione cząsteczki po drugiej stronie błony.
<videoflash>JShwXBWGMyY&feature=related</videoflash>
Transport aktywny — do zachowa¬nia wewnątrzkomórkowego składu jonowego komórek i do
wprowadza¬nia cząsteczek, których stężenie na zewnątrz jest mniejsze niż w komórce,
niezbędny jest aktywny transport cząsteczek i jonów wbrew ich gradiento¬wi
elektrochemicznemu; transportowanie cząsteczek wbrew gradientowi stężeń z udziałem
nośników białkowych i nakładzie energii (często z ATP), np. transport jonów Na+ i K+ za
pomocą mechanizmu pompy jonowej: sodowo — potasowej (zlokalizowana w błonach
plazmatycznych grupa specyficznych białek; transportuje ona jony sodu z wnętrza komórki na
zewnątrz, jednocześnie przenosząc jony potasu w kierunku odwrotnym. Na jedną rozłożoną
przez tę pompę cząsteczkę ATP przypada transport trzech jonów sodu i dwóch jonów potasu).
W tym wypadku wytwarzany gradient stężenia dotyczy cząstek obdarzonych ładunkiem, zatem
w poprzek błony tworzy się nie tylko gradient stężenia, lecz i także gradient potencjału
elektrycznego. Schemat działania pompy sodowo — potasowej znajdziesz tutaj.Wyróżnia się:
translokację grupową — energia do transportu danej cząsteczki równa jest energii
potrzebnej do wytworzenia nowych wiązań kowalencyjnych w transportowanej
cząsteczce,
transport aktywny pierwotny — energia do transportu danej cząsteczki równa jest
energii potrzebnej do wytworzenia nowych wiązań kowalencyjnych w nośniku (źródło:
hydroliza cząsteczki ATP),
transport aktywny wtórny — aktywnie transportowana pierwsza substancja tworzy
gradient potencjału elektrochemicznego, który warunkuje transport innej substancji.
Transport większych cząstek
Schemat endocytozy z podziałem na jej rodzaje
Endocytoza — pobranie do wnętrza komórki cząstek poprzez wytworzenie z błony komórkowej
wodniczki, która oderwawszy się od plazmallemy przeniesie pobraną cząstkę do cytoplazmy.
Przebieg endocytozy: wpuklenie błony → zamknięcie pęcherzyka (wodniczki) → transport
pęcherzyka w głąb cytoplazmy. Wyróżniamy:
fagocytozę — transport bez ubytków błony; polega na otoczeniu pochłanianych
cząsteczek przez mikrofałdy błony komórkowej i utworzeniu wokół nich wakuoli; cząstka
pokarmu zostaje strawiona i wchłonięta do cytoplazmy, a niestrawione resztki są
wyrzucane na zewnątrz, gdy wodniczka z powrotem łączy się z błoną komórkową; na
drodze fagocytozy komórka pochłania duże cząstki pokarmowe np. bakteri,
pinocytozę — transport z ubytkami błony biologicznej; małe drobinki płynu zostają
uwięzione w mikrofałdach błony komórkowej, z której odrywają się po stronie
cytoplazmy drobne pęcherzyki; płynna zawartość pęcherzyków przenika powoli do
cytoplazmy, zaś pęcherzyki powoli zmniejszają się, aż w końcu zanikają (pęcherzyki
zostają w całości rozłożone enzymatycznie, przy udziale lizosomów, a następnie
rozproszone w cytoplazmie).
Egzocytoza (cytopempsja) — proces przeciwstawny do endocytozy; to proces uwalniania
metabolitów powstających wewnątrz komórki (np. hormonów czy enzymów), a także
produktów ubocznych metabolizmu do przestrzeni pozakomórkowej. To również podstawowy
mechanizm powiększania się błon. Przebieg egzocytozy: transport pęcherzyka w kierunku
błony komórkowej → łączenie pęcherzyka z błoną komórkową → otwarcie pęcherzyka.
Istnieją dwa główne typy pęcherzyków uczestniczących w transporcie substancji do komórki:
dołeczki okryte — zagłębienia otoczone przez specjalne białka związane z błoną; służą do
przenoszenia substancji w głąb komórki,
kaweole — zagłębienia otoczone przez białko kaweolinę; mogą skupiać substancje z
przestrzeni zewnątrzkomórkowej, które dalej przedostają się do cytozolu (cytoplazmy) —
potocytoza — a kaweole pozostaja dalej w formie zagłębień i nie tworzą pęcherzyków; w
przypadku gdy kaweole tworzą pęcherzyki mamy do czynienia z trans cytozą; kaweole
umożliwiają również uruchamianie systemu wtórnych przekaźników komórkowych przez
zjawiska działające na komórkę z zewnątrz (wewnątrzkomórkowa sygnalizacja).
Cytoplazma
Cytozol, substancja o pozornie jednolitej strukturze, mogącą przyjmować różną postać od cieczy do
galaretki. Cytoplazmę podstawową przenikają układy błonowe. Od zewnątrz ogranicza ją błona
komórkowa — plazmolemma.
Rola
Wypełnia komórki substancją o pozornie jednolitej strukturze i nadaje jej kształt.
Utrzymuje organelle.
Uczestniczy w podziale komórkowym; umożliwia ruchy chromosomów w czasie mitozy i
mejozy.
Dzięki jej ruchom i elementom cytoszkieletu możliwe jest przemieszczanie organelli i transport
substancji w komórce.
Środowisko wielu reakcji chemicznych (zachodzi w niej wiele reakcji metabolicznych, np.
glikoliza — pierwszy etap oddychania komórkowego).
Skład i właściwości
W przypadku komórki eukariotycznej jest to część protoplazmy, która pozostaje poza jądrem
komórkowym. W przypadku komórki prokariotycznej jest to cała protoplazma (ponieważ komórka ta
nie posiada jądra komórkowego). U protistów (głównie u pierwotniaków) można zaobserwować
wyraźnie rozróżnienie na dwa obszary: ektoplazmę (stałą, zewnętrzną warstwę cytoplazmy komórki,
przylegającą do błony komórkowej) i endoplazmę (płynną, zlokalizowaną w części bardziej
wewnętrznej). Cytoplazma jest to płynny koloid wodny zawierający:
związki organiczne takie jak: białka, tłuszczowce, węglowodany, RNA,
związki nieorganiczne pierwiastków takich jak wapń, magnez, potas, cynk, miedź, mangan,
fosfor, tlen, chlor, siarka, węgiel, azot czy bor,
wodę (faza rozpraszająca) oraz
organelle — „twory” wyspecjalizowane do pełnienia określonej funkcji (mitochondria, plastydy,
lizosomy, peroksysomy, wakuole, cytoszkielet, retikulum endoplazmatyczne, Aparat Golgiego),
które omówione zostaną w dalszej części skryptu.
Zasadniczym elementem strukturalnym cytoplazmy podstawowej są białka mające postać globularną
lub fibrylarną:
Model filamentu
aktynowego
Struktura
mikrotubuli
Model
filamentów
pośrednich
Mikrofilamenty — średnica 5-9 nm; cienkie włókna białkowe zbudowane z białka aktyny
znajdujące się w cytoplazmie; odpowiedzialne za ruch cytoplazmy, zmianę kształtu komórki i
jej ruch pełzakowaty dzięki pseudopodiom oraz endocytozę; stanowią mechaniczną podporę
dla komórki i różnych elementów komórkowych; zlokalizowane są tuż pod błoną komórkową.
Mikrotubule — średnica 20-27 nm; utworzone są ze spiralnie zwiniętych łańcuchów cząsteczek
białka; ełnią funkcję cytoszkieletu nadając komórce kształt a nawet przyczyniając się do jego
zmiany; mogą również tworzyć stałe struktury takie jak: rzęski lub wici, umożliwiające ruch
komórki; biorą udział w podziałach komórkowych.
Filamenty pośrednie — średnica 10 nm; zbudowane są z wielu skręconych razem białek
włóknistych; bardzo wytrzymałe, przez co chronią komórkę przed mechanicznymi
uszkodzeniami na skutek rozciągania; pomagają utrzymać określony kształt komórki; pary
filamentów tworzą tzw. dimery → dimery tworzą tetrametr, który poprzez wiązanie
kowalencyjne łączy się z drugim tetrametrem, tworząc ostatecznie filament pośredni.
Ruchy cytoplazmy
Ruchy cytoplazmy są najlepiej widoczne w komórkach roślinnych. Ich przejawem jest
przemieszczanie się zawieszonych w cytoplazmie chloroplastów (patrz film). Ruchy cytoplazmy
spowodowane są skurczami włókienek kurczliwych.
Rodzaje ruchów:
1. Cyrkulacyjny — ruch cytoplazmy po mostkach cytoplazmatycznych → ruch fontannowy —
szczególna odmiana ruchu cyrkulacyjnego, w którym cytoplazma płynie wokół dwóch wakuoli
w przeciwnych kierunkach.
2. Rotacyjny — ruch cytoplazmy wokół centralnie ułożonej wakuoli.
3. Pulsacyjny — cytoplazma płynie raz w jednym, raz w drugim kierunku (wokół wakuoli).
Ruch rotacyjny
cytozolu wokół
wakuoli
Ruch pulsacyjny
cytozolu wokół
wakuoli
Ruch
cyrkulacyjny
cytozolu wokół
wakuoli
Jądro komórkowe
Jest to największe pojedyncze organellum w komórce otoczone błonę komórkową, które występuje u
organizmów eukariotycznych. Stanowi ono centrum kontroli komórki, ponieważ jego główną funkcją
jest przechowywanie i powielanie informacji genetycznej oraz kontrolowanie czynności komórki.
Znajduje się tu ponad 99% materiału genetycznego w postaci DNA (pozostała część DNA znajduje się
w macierzy mitochondriów). W zdecydowanej większości komórki posiadaja tylko jedno jądro — są to
monokariocyty. Są jednak komórki posiadające dwa (bikariocyty, no. hepatocyty) lub kilka jąder
(polikariocyty, np. komórki mięśni poprzecznie prążkowanych).
Jądro (komórka cebuli)
Funkcje
Ośrodek decyzyjny komórki.
Przechowanie informacji na temat budowy i funkcjonowania komórki.
Synteza DNA (replikacja informacji genetycznej przed podziałem jądra, samopowielanie).
Synteza RNA (transkrypcja).
Synteza białek (rybosomy powstające w jąderku).
Budowa
Schemat budowy jądra komórkowego u
człowieka.
Jądro komórkowe jest to najbardziej widoczna organellą w komórce. Zwykle jest kuliste lub owalne,
o wielkości około 5-10 μm (ssaków średni rozmiar jądra wynosi około 6 μm i stanowi około 10%
objętości ich komórki). Jądra barwią się barwnikami zasadowymi (takimi jak hematoksylina), a więc
są zasadochłonne.
Otoczka i pory jądrowe
Otoczka (błona) jądrowa, jest to podwójna błona białkowo-lipidowa, która odgranicza wnętrze jądra
komórkowego od cytoplazmy. Pełni ona rolę bariery, chroniącej wnętrze jądra.
Występuje tu:
Błona zewnętrzna (pokryta rybosomami) i wewnętrzna — oddzielone są od siebie o 20-40 nm
(mogą też się stykać).
Przestrzeń okołojądrowa — wystepuje pomiędzy błonami.
Pory jądrowe (zbudowane z wielu białek) — miejsca, w których błony otoczki jądrowej łączą się
ze sobą, tworząc kanały; umożliwiają bierną i czynną wymianę makromolekuł z cytoplazmą (do
jądra wnikają np. białka, zaś do cytoplazmy przenikają kompleksy rybonukleoproteinowe
mRNA oraz dojrzałe rybosomy); przeciętnie w komórkach eukariotycznych jest 10-20
porów/mm²; swobodna dyfuzja cząsteczek wynosi jedynie 9 nm, zaś duże cząstki muszą być
aktywnie transportowane między jądrem a cytoplazmą.
Blaszka jądrowa
Jest to cienka warstwa filamentów pośrednich typu V (grubość 10-100 nm). Zbudowane są one z
białka (laminy i białek globularnych) przylegającego do wewnętrznej błony otoczki. Pojedynczy
monomer lamin zawiera helikalną domenę, dzięki której dwie cząsteczki białka mogą owinąć się
wokół siebie, tworząc dimer. Z kolei dwa dimery łączą się tworząc tetramer (protofilament). Osiem
protofilamentów łączy się bokami i zwija, tworząc filament, podobny w swej strukturze do liny.
Blaszka nadaje odpowiedni kształt jądru komórkowemu, zapewniaj jądru wytrzymałość mechaniczną
oraz uczestniczy w organizacji strukturalnej chromatyny.
Chromosomy
Chromosom
submetacentryczny: 1 —
chromatyda 2 — centromer —
miejsce złączenia dwóch
chromatyd 3 — ramię krótkie
4 — ramię długie
To forma organizacji materiału genetycznego wewnątrz komórki, w postaci wielu liniowych
cząsteczek DNA zorganizowanych w struktury. Przyjmują postać chromatyny — kompleksu DNA oraz
białek. Są zbudowane z dwóch chromatyd siostrzanych (podłużnych jego części) połączonych w
jednym punkcie centromerem. Każda cząsteczka DNA buduje jedną chromatydę. Struktura
chromosomu nie jest niezmienna, podlega on bowiem zmianom zwanym mutacjami. Mutacje
dotyczące bezpośrednio chromosomów to aberracje chromosomowe lub mutacje genomowe.
Dwie postaci chromatyny:
euchromatyna — zawiera geny, które są częściej transkrybowane przez komórkę; jest mniej
skondensowana; dostrzegalna jako jasnobarwliwe obszary,
heterochromatyna — zawiera geny transkrybowane rzadziej; bardziej skondensowana (nić
DNA jest szczególnie mocno upakowana); są to ciemno barwiące się obszary, często
przylegające do błony jądrowej:
fakultatywna — zawiera geny zorganizowane w postaci heterochromatyny jedynie w
pewnych typach komórek,
konstruktywna — tworzy elementy strukturalne chromosomu, jak centromery lub
telomery.
Jąderko
Mikrofotografia elektronowa
jądra komórkowego z ciemno
zabarwionym jąderkiem.
Jest to zagęszczenie chromatyny; najlepiej widoczna pod mikroskopem część jądra.
Luźno zawieszony w nukleoplazmie obszar o średnicy 1-3 mikrometra.
Odpowiada za syntezę RNA (głównie tRNA) i tworzenie rybosomów (złożone podjednostki
rybosomów są największymi strukturami przechodzącymi przez pory jądrowe).
Komórki nieaktywne maja niewyraźne jąderka, zaś komórki aktywne metabolicznie mają
jąderka duże i mnogie.
Zanika podczas podziału komórkowego.
Nie zawiera błony.
Składa się z trzech regionów:
wewnętrznych center fibrylarnych (zlokalizowane są w nich geny kodujące rRNA )
otoczonych przez:
gęsty komponent fibrylarny (utworzony z włókien o średnicy 4–5 nm i długości do 50
nm), który otoczony jest z kolei przez
komponent granularny (ziarna o średnicy 15–20 nm w postaci pól wymieszanych z
gęstym składnikiem włóknistym).
Transkrypcja rDNA zachodzi zarówno w centrach fibrylarnych jak i na granicy między centrami
fibrylarnymi oraz gęstym komponentem fibrylarnym.
Komórki HeLa z dobrze widocznymi jądrami,
wybarwionymi za pomocą barwnika
Hoechst. Komórki w środku i z prawej
znajdują się w interfazie. Komórka z lewej
przechodzi proces mitozy, stąd jej DNA jest
skupione i gotowe do podziału.
Macierz jądrowa
Sieć włókien białkowych tworzących wewnętrzny szkielet jądra komórkowego.
Zbudowana z filamentów (o średnicy 3-5 nm) i ziarenek (o średnicy 15-30 nm) zanurzonych w
kariolimfie.
W jej skład wchodzi co najmniej 13 rodzajów białek (w tym białak funkcjonalne i strukturalne).
Około 98% macierzy jądrowej stanowią białka, reszta to kwasy nukleinowe oraz fosfolipidy.
Odpowiada za utrzymanie struktury przestrzennej chromatyny.
Odgrywa rolę w regulacji syntezy i transkrypcji DNA.
Stany jądra komórkowego
Jądro komórkowe może znajdować się w trzech różnych stanach:
Jądro interfazowe — między dwoma następującymi po sobie podziałami.
Jądro mitotyczne — w czasie podziału komórki — występuje w komórkach dzielących się.
Jądro metaboliczne — występuje w komórkach wyrośniętych, kieruje procesami przemiany
materii.
Mitochondrium
To cylindryczne organelle otoczone błoną; wydłużone struktury cytoplazmatyczne
przybierające kształt nitek lub lasek.
Występują w komórkach eukariotycznych.
Są odpowiedzialne za proces oddychania tlenowego, czyli proces przekształcania energii
chemicznej związków organicznych w energię wiązań ATP.
W warunkach typowych mają długość od 0,5 do 2 mikrometrów.
Każde mitochondriom ma własny DNA oraz system syntezy białek niezależny od jądra
komórkowego (dlatego uważa się, że mitochondria rozwinęły się w komórkach ludzkich jako
symbiotyczne organizmy protokariotyczne, podobne do bakterii).
Liczba mitochondriów w komórce zależy od zapotrzebowania komórki na energię (najliczniej
występują np. w komórkach mięśni poprzecznie prążkowanych).
Mogą zmieniać swój kształt i rozmiary.
Powstają przez wzrost i podział już istniejących mitochondriów.
Całkowita objętość jednej komórki wynosi około 20% jej objętości całkowitej.
Pojedyncza komórka zawiera od kilku sztuk do kilku tysięcy mitochondriów (zwykle kilkaset).
W wyniku ruchów cytoplazmy lub dzięki związaniu się z elementami cytoszkieletu,
mitochondria mają zdolność do przemieszczania się w kierunku miejsca o zwiększonym
zapotrzebowaniu na energię.
Mikrofotografia elektronowa dwóch
mitochondriów pochodzących z płuca ssaka,
pokazująca ich matriks oraz błony.
Budowa
Błąd przy generowaniu miniatury
(process:16595): Gtk-WARNING **: Locale not supported
by C library.
Using the fallback 'C' locale.
Background RRGGBBAA: ffffff00
Area 0:0:561.577:358.559 exported to 400 x 255 pixels
(64.1052 dpi)
Bitmap saved as:
/var/www/html/edu/images/tmp/transform_7fbb464a1ed4-1
.png
GLib: Cannot convert message: Could not open converter
from 'UTF-8' to 'ANSI_X3.4-1968'
(inkscape:16595): GLib-ERROR **:
/build/glib2.0-y6934K/glib2.0-2.42.1/./glib/gmem.c:103:
failed to allocate 16384 bytes
/var/www/html/edu/includes/limit.sh: line 101: 16594
Trace/breakpoint trap /usr/bin/timeout
$MW_WALL_CLOCK_LIMIT /bin/bash -c "$1" 3>&Schemat mitochondrium zwierzęcego.
Składa się z dwóch błon zbudowanych z dwuwarstwy lipidowej oraz rozmieszczonych w niej
białek:
zewnętrznej — błona, która otacza całe organellum, oddzielając je od środowiska
zewnętrznego; współczynnik białek do lipidów wynosi około 1:1 wagowo; zawiera duże
ilości wyspecjalizowanych białek transportowych (poryny — średnica około 2-3 nm),
które umożliwiają swobodne przechodzenie cząsteczek o ciężarze do ok. 10 kD z cytozolu
do przestrzeni międzybłonowej; białka o większych cząsteczkach mogą pokonać błonę
zewnętrzną tylko przy pomocy transportu aktywnego,
wewnętrznej — znajduje się w środku organellum; zachodzą tu reakcje chemiczne,
przekształcające energię cząsteczek substancji pokarmowych w energię wiązań ATP;
wysoki współczynnik białek do lipidów (ponad 3:1 wagowo); powierzchnia błony
wewnętrznej mitochondrium, jest kilkukrotnie większa od powierzchni zewnętrznej błony
mitochondrialnej; jest wysoce nieprzepuszczalna dla małych jonów z powodu dużej
zawartości fosfolipidu kardiolipiny (umożliwia to wytworzenie gradientu
elektrochemicznego podczas produkcji wysokoenergetycznych metabolitów
komórkowych). Występuje w niej kilka typów białek o różnych funkcjach:
białka biorące udział w reakcjach redoks fosforylacji oksydacyjnej,
syntaza ATP wytwarzająca ATP w macierzy mitochondrialnej,
specjalne białka kontrolujące przechodzenie metabolitów do oraz z macierzy,
białka zajmujące się importem innych białek,
białka, które zajmują się syntezą oraz rozkładem innych białek.
Grzebienie mitochondrialne (tworzone przez fałdy błony wewnętrznej) — mocno
pofałdowana błona wewnętrzna mitochondriom (powoduje to zwiększenie powierzchni), w
którą wbudowane są przenośniki łańcucha oddechowego oraz syntezy ATP; to miejsce w
którym zachodzi fosforylacja oksydacyjna.
Przestrzeń międzybłonowa (pomiędzy błoną zewnętrzną a wewnętrzną) — zawiera:
substraty metaboliczne, dyfundujące przez błonę zewnętrzną,
ATP wytworzone przez mitochondrium,
jony wypompowane z przestrzeni macierzy podczas oksydatywnej fosforylacji (proces
polegający na przyłączeniu reszty kwasu ortofosforowego do adenozynodifosforanu —
ADP — przy udziale energii — powstaje wtedy adenozynotrifosforanu — ATP); ponieważ
błona zewnętrzna jest przepuszczalna dla małych cząsteczek, stężenie substancji takich
jak jony lub cukry w przestrzeni międzybłonowej jest takie same jak w cytozolu.
Macierz mitochondrialna (wewnętrzna przestrzeń mitochondriom, matriks) —
bezpostaciowa substancja wypełniająca wnętrze mitochondrium; przestrzeń wewnątrz
mitochondrium, ograniczona błoną wewnętrzną wypełnioną wodnym roztworem białek (np.
enzymy β-oksydacji kwasów tłuszczowych, cyklu Krebsa czy syntezy steroidów) i metabolitów
zużywanych na potrzeby mitochondriom; zawiera materiał genetyczny w postaci kilku kopii
mitochondrialnego DNA (mtDNA), rybosomy mitochondrialne i tRNA mitochondrialne;
Funkcje
1. Przemiany energetyczne — wytwarzanie ATP — zachodzi dzięki utlenianiu głównych
produktów rozkładu glukozy — pirogronianu i NADH, wytwarzanych w cytozolu; jest to proces
oddychania komórkowego (oddychanie tlenowe), zależny jest od obecności tlenu (gdy ilość
tlenu dostarczanego mitochondriom jest ograniczona, produkty glikolizy przetwarzane są w
ramach oddychania beztlenowego, procesu który nie zachodzi w mitochondriach). Należy
zwrócić uwagę na fakt, iż energia w komórce nie może istnieć w stanie wolnym, gdyż ulegałaby
rozproszeniu. Z tego względu musi zostać zmagazynowana w wiązaniach chemicznych.
Okazuje się, że najlepsze do tego jest adenozynotrifosforan (ATP) — w jego budowie występują
dwa wysokoenergetyczne wiązania. ATP wytwarzane jest z ADP i Pi (reszty ortofosforanowej)
w wyniku działania syntazy ATP. Rotacja jej odpowiedniego segmentu umożliwia syntezę ATP.
Energia niezbędna do syntezy dostarczana jest przez gradient elektrochemiczny. Gdy stężenie
protonów w przestrzeni międzybłonowej wzrasta, po przeciwnych stronach błony wewnętrznej
wytwarza się silny gradient elektrochemiczny (jest on powodowany parciem protonów do
powrotu do macierzy mitochondrialnej). Jedyną mozlwością powrotu jest kompleks białkowy
syntazy ATP — energia przechodzących przez niego protonów używana jest do syntezy ATP z
ADP oraz anionu fosforanowego (Pi). Każda Komórki produkują ATP tylko na własne potrzeby,
ponieważ ATP jest związkiem nietrwałym (nie może być transportowane z komórki do komórki)
→ ADP + Pi + energia → ATP. UWAGA: Produkcja ciepła — wyciekanie protonów; w pewnych
warunkach protony mogą przedostawać się do macierzy mitochondrialnej nie wytwarzając
ATP, co powoduje rozproszenie energii potencjalnej gradientu elektrochemicznego protonów w
postaci ciepła.
2. Magazynowanie jonów wapnia — mitochondria mogą przejściowo magazynować wapń;
wapń może regulować szereg reakcji i jest istotne dla przewodnictwa sygnałów w komórce.
3. Regulowanie potencjału błonowego.
4. Regulacja stanu redoks komórki (potencjał redoks jest w pewnym sensie miarą
koncentracji elektronów, które zdolne są do przechodzenia na inne substancje i utleniania ich).
5. Synteza hemu (niebiałkowa część wielu enzymów).
6. Synteza sterydów.
Organelle otoczone pojedynczą błoną komórkową
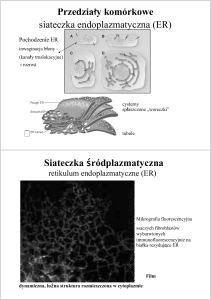
Reticulum endoplazmatyczne
Elektronowy obraz tkanki płuca. Widoczne
retikulum endoplazmatyczne i mitochondria
Retikulim endoplazmatyczne (siateczka śródplazmatyczna) i aparat Golgiego (patrz dalej) stanowią
dwa odrębne obszary wzajemnie połączonego przedziału błoniastego uczestnicząc w biosyntezie
białek i lipidów.
Typy
Schemat: Obraz jądra komórkowego,
siateczki śródplazmatycznej i aparatu
Golgiego. 1. Jądro komórkowe 2. Por
jądrowy 3. Szorstka siateczka
śródplazmatyczna (Rough endoplasmic
reticulum — rER) 4. Gładka siateczka
śródplazmatyczna(sER) 5. Rybosom na rER
6. Białka, które są transportowane 7.
Pęcherzyk transportowy 8. Aparat Golgiego
9. Biegun cis aparatu Golgiego 10. Biegun
trans aparat Golgiego 11. Cysterna aparatu
Golgiego
Szorstkie (granularne, ER-g) — rozbudowywane w komórkach szybko rosnących oraz w
komórkach w których zachodzi biosynteza białek; charakteryzujące się obecnością licznych
rybosomów, osadzonych na jego zewnętrznej powierzchni (rybosomy produkujące peptydy z
sekwencja sygnałowa dla białek błonowych lub wydzielniczych zostają przyczepione do
powierzchni siateczki śródplazmatycznej, na której dalsza część peptydu podlega translacji);
synteza białek przez ER-g prowadzi bądź do włączenia białek do błony siateczki bądź do
zatrzymania w świetle siateczki białek przeznaczonych do wydzielenia → nowo wytworzone
białka docieraja do retikulum endoplazmatycznego gładkiego a następnie do aparatu Golgiego.
Gładkie (agranularne, ER-a) — rozwinięte w komórkach syntezujących niebiałkowe produkty
organiczne niezwiązane z rybosomami; to miejsce syntezy lipidów błonowych, przetwarzania
białek, oraz detoksykacji czyli usunięcia z organizmu trujących substancji; od błon siateczki
śródplazmatycznej gładkiej mogą oddzielać się pęcherzyki, które przekształcają się w wakuole.
Rybosomy
Rybosomy — drobne, elektronowo gęste cząstki; zbudowane z rRNA i białek; to struktury
występujące w każdej komórce, odpowiadają za proces biosyntezy białek.
Atomowa struktura dużej podjednostki 50S
rybosomu. Kolorem niebieskim zaznaczone
są białka, pomarańczowym RNA a
czerwonym adenina 2486 w centrum
aktywnym
Typy
Rybosomy małe, wolne — występują w komórkach Procaryota oraz w mitochondriach i
plastydach Eucaryota; stała sedymentacja (70S); nie są związane z błonami biologicznymi;
służą one do syntezy białek nieeksportowanych poza komórkę, takich jak enzymy
wewnątrzkomórkowe, białkowe elementy błon komórkowych, białka cytoplazmy czy białka
cytoszkieletu.
Rybosomy duże — występują w cytoplazmie komórek Eucaryota; stała sedymentacji (80S);
związane są z błonami biologicznymi (zwykle to błony reticulum endoplazmatycznego w
których następuje synteza białek eksportowanych transportowanych przez siateczkę
śródplazmatyczną także poza błony komórki — hormony białkowe, kolagen, białka
wydzielnicze, enzymy lizosomalne, białka wchodzące w skład błon, nici elastynowe dla tkanki
łącznej).
Funkcje
Biosynteza białek i lipidów.
Udział w transporcie komórkowym — pozwala na szybkie transporty wewnątrzkomórkowe, co
jest związane z faktem, iż cytoplazma jest w nim rzadsza.
Udział w przemianach węglowodanów.
Detoksykacja (szczególnie w komórkach wątroby), np. rozkładanie związków rakotwórczych.
Udział w przekazywaniu informacji w komórce, np. impulsu nerwowego do wnętrza komórek
mięśniowych.
Tworzy przedziały subkomórkowe, co umożliwia zachodzenie w bliskim sąsiedztwie
przeciwstawnych reakcji, które nie przeszkadzają sobie wzajemnie.
Z jego błon mogą powstawać aparaty Golgiego, lizosomy i wodniczki.
<videoflash>vJ6ygMhzQEo</videoflash>
Aparat Golgiego
Mikrofotografia aparatu Golgiego
widocznego jako stos półkolistych czarnych
pierścieni w dolnej części obrazu. Liczne
koliste pęcherzyki są widoczne w pobliżu
organellum
Błoniasta struktura komórki (zazwyczaj znajduje się w pobliżu jądra); występuje niemal we
wszystkich komórkach eukariotycznych. Po przejściu przez cysterny gładkiej siateczki
śródplazmatycznej dalsze syntetyzowanie przetwarzanych makrocząstek zachodzi własnie w
aparacie Golgiego. Tu są sortowane, opakowywane i transportowane produkty komórkowe.
Budowa
powstaje z szorstkiej siateczki śródplazmatycznej
podstawową jednostką strukturalną aparatu Golgiego jest diktiosom (średnica ok. 1
mikrometr) — układ 6-30 spłaszczonych woreczków, zbiorników oraz połączonych z nimi rurek
i pęcherzyków, których ściany są zbudowane z błony; w obrębie diktiosomu wyróżnia się dwa
bieguny (biegun cis-formowania — gdzie znajdują się enzymy: transferaza Nacetyloglukozoaminy oraz transferazy: galaktozylowa, fukozylowa, sialowa oraz biegun transdojrzewania — który stanowi stację rozdzielczą i sortująca, w której produkty z wnętrza
diktiosomu zostają rozsortowane zależnie od przeznaczenia i zapakowane do odpowiedniego
typu pęcherzyków)
Funkcje
Synteza różnych związków (np. w komórkach roślinnych są to wielocukry do budowy ściany
komórkowej, synteza polisacharydów oraz mukopolisacharydów: glikozoaminoglikanów,
hemicelulozy, pektyny).
Modyfikacje reszt cukrowych glikoprotein i glikolipidów.
Kieruje przepływem makrocząstek.
Segreguje makrocząsteczki (grupuje wg budowy chemicznej).
Posiadają zdolność redukcji azotanu(V) srebra(I).
Cząsteczki o różnym pochodzeniu są pakowane w pęcherzyki odrywające się od cystern i
transportowane do innych organelli lub po fuzji z błoną komórkową wydalane na zewnątrz
komórki.
Lizosomy
Lizosomy (w komórce zwierzęcej) niewielkie (0,02-0,8 μm) struktury kuliste lub owalne, otoczone
pojedynczą błoną wypełnione enzymami trawiennymi. Znajdują się tam enzymy służące do trawienia
lipidów (lipazy), cukrowców (amylazy), białek (proteazy) oraz kwasów nukleinowych (nukleazy).
Lizosom działa jako wewnątrzkomórkowy system trawienny (substancje, które mają ulec strawieniu,
docierają do lizosomu w pęcherzykach). Wewnątrz lizosomu utrzymywane jest pH na poziomie ok. 5.
To niskie pH aktywuje silnie działające enzymy o charakterze kwaśnych hydrolaz, które pochodzą z
pęcherzyków powstających w aparacie Golgiego. Łącznie w lizosomach jest obecnych ok. 40 różnych
hydrolaz, czyli enzymów hydrolitycznych. Po strawieniu substancji przez kwaśne hydrolazy nie
dający się strawić materiał bezpostaciowy i resztki błonowe mogą być widoczne w dużych
obłonionych pęcherzykach (ciałkach resztkowych). Po obumarciu komórki błony lizosomów pękają
uwalniając enzymy trawienne do cytoplazmy. To zjawisko jest przyczyną rozpadu wielu komórek po
śmierci organizmu.
Rodzaje lizosomów
Trawienne,
magazynujące ,
grabarze — rozkład obumarłych składników cytoplazmy.
Wakuole
Wodniczki (zabarwione) wewnątrz komórek. Pole
widzenia — ok. 450 μm
Wodniczka to wydzielona przestrzeń w cytoplazmie, otoczona jest pojedynczą błoną białkowolipidową tzn. tonoplastem, wypełniona sokiem komórkowym zawierającym wodę, substancje
zapasowe oraz wydaliny komórki. W komórkach zwierzęcych występuje wiele małych wodniczek
(wakuol), natomiast w roślinnych jedna lub kilka dużych; zajmują do 90% komórki.
Funkcje
Utrzymanie komórki w stanie turgoru (stanu napięcia komórki).
Magazyn wody w komórce.
Magazynowanie zbędnych produktów przemiany materii (u roślin).
Wakuola może gromadzić rozmaite substancje, np:
barwniki — antocyjany, flawony i in. (np. betalainy korzenia buraka),
jony nieorganiczne (m. in. Na+, K+, Ca2+, Cl-, SO42-, NO3-, PO43- ),
ciałka zapasowe (ciała białkowe),
inne związki organiczne:
cukrowce — m. in. glukoza, fruktoza, sacharoza, inulina,
kwasy organiczne,
aminokwasy i białka.
W wakuolach mogą być magazynowane liczne substancje o charakterze obronnym (trujące,
gorzkie, itp. które mogłyby działać szkodliwie np. na cytoplazmę):
białka obronne (np. inhibitory proteaz, chitynaza, lektyny),
żywice fenolowe (np. antocyjaniny, kwas galusowy, eskulina),
terpenoidy (np. kwas oleanolowy, saponiny),
oligosacharydy (np. gentianoza, gentiobioza, stachyoza),
alkaloidy (np. atropina, nikotyna, ajmalicyna),
inne związki azotowe (np. glukozylany, glikozydy),
metabolity wtórne (np. papaweryna, kodeina, dopamina, morfina),
poliamina (np. skopolamina, serpentyna, solanidyna).
Niektóre substancje chronią także przed UV, np. metabolity wtórne (flawonoidy, kumaryny).
Utrzymywanie niskiego stężenia jonów sodowych w cytozolu.
Aktywnie gromadzą jony nieorganiczne (np. azotany, jony wapnia) i w razie potrzeby
dostarczają je do cytozolu.
Wodniczki tętniące — (u pierwotniaków słodkowodnych) usuwające nadmiar wody z komórki.
Wodniczki trawiące — u pierwotniaków reguluje poziom płynów w organizmie i wypuszcza soki
trawienne w celu trawienia pokarmu.
Wakuolę wypełniają
Związki nieorganiczne: woda (ok. 90% całości składu), jony potasowe, sodowe, wapniowe,
magnezowe, cynkowe, siarczanowe, fosforowe, chlorkowe, kryształy szczawianu wapnia
(rafidy, druzy, styloidy) oraz węglanu wapnia (cystolity).
Związki organiczne: wolne aminokwasy, białka, cukry, glikozydy (alkohol+cukier), antocyjany
(barwnik o barwie czerwonej bądź niebieskiej, zależnie od pH), flawony (barwnik o barwie
żółtej), alkaloidy — zwykle trucizny (np. nikotyna, kofeina, kokaina, morfina, teofilina,
teobromina, chinina).
Plazmoliza
Zachodzi w roztworze hipertonicznym; jest to proces tracenia wody w komórce.
Deplazmoliza
Zachodzi w roztworze hipotonicznym; proces odwrotny, polegający na pobieraniu wody przez
wodniczki z roztworu zewnętrznego o mniejszym stężeniu.
Przed plazmolizą
cytoplazma
(różowa)
przylega do
ścian
komórkowych
Cytoplazma w
trakcie
plazmolizy
zmniejszyła
swoją objętość
Ściana komórkowa
Jest to martwy składnik komórki, otoczka komórki leżąca na zewnątrz błony komórkowej.
Funkcje
Stanowi granicę między światem zewnętrznym a wewnętrznym komórki.
Ogranicza wzrost komórki, nadaje jej kształt i sztywność.
Chroni przed urazami mechanicznymi, infekcjami bakteryjnymi i wirusowymi, nadmierną
utratą wody, nadmiernym parowaniem.
Przepuszcza substancje.
Budowa
60% stanowią składniki podłoża; białka, pektyny (polisacharydy i oligosacharydy o zmiennym
składzie), hemicelulozy (niejednorodna grupa polisacharydów i ich pochodnych, połączonych
wiązaniami β-glikozydowymi tworzących rozgałęzione łańcuchy), woda
40% stanowią składniki szkieletowe:
U eukariota: rośliny — celuloza, która tworzy regularne łańcuchy celulozowe. Ich pęczki
to miofibryle. Kilka miofibryli to fibryla celulozowa. Jest pogrążona w macierzy i pełni
funkcję głównego rusztowania ściany komórkowej; grzyby — chityna, glony — koloza i
mannoza (hemicelulozy). To mieszanina wielocukrów amorficznych (lżejsza od celulozy).
U prokariota: sinice, bakterie — kwasy pileminowy, murawinowy, stanowiące razem
mureinę.
Formowanie się ściany komórkowej
Kutykula utrudnia gromadzenie wody na
powierzchni pędu
1. Blaszka środkowa — budowana wspólnie przez dwie komórki z wielocukrów, głównie związków
pektynowych; powstaje podczas podziału i jest pierwszą strukturą, która oddziela
nowopowstałe komórki.
2. Ściana pierwotna — kolejna warstwa oddzielającą; budowana jest przez dwie komórki po obu
stronach blaszki środkowej; jest elastyczna i cienka; zbudowana z różnych wielocukrów o
krótkich łańcuchach (pektyny i hemicelulozy) oraz z długich łańcuchów celulozy.
3. Ściana wtórna — od strony protoplastu odkładane są na powierzchnie ściany pierwotnej nowe
warstwy celulozy; jest grubsza niż ściana pierwotna; zbudowana jest z różnych wielocukrów, w
tym celulozy; podlega przekształceniom:
Inkrustracji — odkładaniu się substancji mineralnych między wiązkami celulozy.
Wyróżniamy:
Mineralizację — wnikanie związków mineralnych jak węglanu wapnia CaCO3 lub
krzemionki SiO2 . Ściany są twarde, sztywne, ale łamliwe.
Drewnienie (lignifikacja, ligninizacja) — wnikanie ligniny (drzewnika); ściana
twardnieje, staje się odporna na zgniatanie i przerwanie, a zawartość w niej wody
bardzo spada;
Adkrustacji — odkładaniu się substancji mineralnych na powierzchni pierwotnej ściany
komórkowej. Adkrustacja jest ochroną przed nadmiernym przepuszczaniem wody przez
ścianę. Wyróżniamy:
Kutynizację i woskowacenie — powleczenie zewnętrznej strony ściany kutyną i/lub
woskiem, substancjami pochodzenia tłuszczowego; proces odkładania kutyny
między warstwami celulozy w obrębie ściany komórkowej (kutyna uważana jest za
substancję najbardziej odporną w przyrodzie na działanie czynników chemicznych).
Suberynizację (korkowacenie) — powleczenie wewnętrznej strony ściany suberyną
(substancja złożona przede wszystkim z długołańcuchowych kwasów tłuszczowych
oraz ich estrów z alkoholami alifatycznymi), a potem jeszcze jedną warstwą
celulozy; tak zmodyfikowana ściana staje się nieprzepuszczalna dla wody; suberyna
odkłada się na pierwotnej ścianie komórkowej, na przemian z warstwami kutyny i
wosku; ściany są wtedy warstwą izolacyjną chroniącą roślinę przed wpływem
niekorzystnych warunków środowiska.