Uploaded by
common.user4525
Fizjologia człowieka w zarysie - Podręcznik

WYDAWNICTWO LEKARSKIE PZWL
Prof. dr hab. med
Władysław Z.Traczyk
fizjologia
człowieka
co zarysie
Wydanie VII
WARSZAWA
WYDAWNICTWO LEKARSKIE PZWL
© Copyright by Władysław Z. Traczyk, Warszawa 1976, 1978, 1982, 1989, 1992, 1997,
2000, 2002
Wszystkie prawa zastrzeżone. Przedruk i reprodukcja
wjakiejkolwiek postaci całości bądź części książki bez
pisemnej zgody autora i wydawcy są zabronione.
Redaktor Elżbieta Michalska
Redaktor techniczny Franciszka Wyszomirska
Korekta Zofia Kalina
Projekt okładki i strony tytułowej Małgorzata Różańska
ISBN 83-200-2694-6
Wydanie VII (dodruk)
Wydawnictwo Lekarskie PZWL
00-238 Warszawa, ul. Długa 38/40
tel. (+22) 831-21-81, http://www.pzwl.pl
Drukarnia Wydawnicza
im. W. L. Anczyca S.A. - Kraków
Słowo wstępne
do szóstego wydania
Niezbędne zmiany i uzupełnienia zostały wprowadzone do obecnego
szóstego wydania tej książki. W ostatnim dziesięcioleciu zaznaczył się
znaczny postęp wiedzy w zakresie komunikacji humoralnej pomiędzy
komórkami organizmu zarówno sąsiadującymi ze sobą, jak i odległymi.
Aby zachować zwięzłość tekstu, substancje i czynniki chemiczne biorące
udział w tej komunikacji zostały tylko wyliczone, nie omawiano natomiast
molekularnego mechanizmu ich działania. Z tych względów podręcznik
ten, który ma charakter repetytorium, nie może służyć do systematycznych i gruntowych studiów przedmiotu.
W obecnym wydaniu wymieniono większość rycin i dokonano zmian
w tabelach i schematach. Wielkości są wyrażone w jednostkach międzynarodowego układu SI zgodnie z uchwałą Zgromadzenia Ogólnego
Światowej Organizacji Zdrowia (WHO) z maja 1977 roku. W tekście
i w tabelach niektóre wielkości podane są jednocześnie w jednostkach
układu SI i w jednostkach tradycyjnych.
Serdecznie dziękuję tym wszystkim, którzy podzielili się ze mną swoimi
uwagami o wydaniach poprzednich, jak i wydaniu obecnym. Szczególnie
dziękuję Panu Profesorowi Franciszkowi Kokotowi, który zapoznał się
z całym czwartym i piątym wydaniem i podzielił się ze mną swoimi
propozycjami dotyczącymi zmian i poprawek. Serdecznie dziękuję za
uwagi dotyczące poszczególnych rozdziałów obecnego wydania, które
przekazali mi: Pan Doktor hab. Andrzej Godlewski — czynność komórek nerwowych, Pan Profesor Janusz Greger — prawa fizjologiczne
organizmu człowieka, Pan Profesor Franciszek Kokot — płyny ustrojowe
i czynność nerek, Pan Profesor Stanisław Konturek — układ trawienny,
Pan Profesor Bohdan Lewartowski — czynność komórek mięśniowych
i czynność serca, Pan Profesor Andrzej Niechaj — czucie i ruch,
Pan Profesor Marek Pawlikowski — kontrola środowiska wewnętrznego,
6
S/owo wstępne do szóstego wydania
Pani Profesor Teresa Pajszczyk-Kieszkiewicz — rozród, Pan Profesor
Jerzy Rożniecki — oddychanie, Pani Docent Zdzisława Traczyk — czynność tkanek krwiotwórczych i krwi i Pan Profesor Andrzej Trzebski —
krążenie.
Składam również najserdeczniejsze podziękowania współpracownikom
z Zakładu Fizjologii Akademii Medycznej w Łodzi, a szczególnie Pani
Ewie Zając, za pomoc w przygotowaniu książki do druku.
Łódź, dnia 26 maja 1997 roku
Autor
Od Wydawcy
do siódmego wydania
Podstawę obecnego wydania książki stanowi szóste wydanie, znacznie
zmienione w stosunku do wydań poprzednich, uwzględniono w nim
bowiem postęp, jaki dokonał się w ostatnich latach w nauce o fizjologii
człowieka. W siódmym wydaniu wprowadzono tylko niezbędne
zmiany i uzupełnienia.
Słowo wstępne
do pierwszego wydania
W okresie kilkuletnich wykładów zetknąłem się z opinią moich słuchaczy
o potrzebie zwięzłego podręcznika fizjologii człowieka napisanego w sposób ułatwiający powtórzenie przed egzaminem najważniejszych wiadomości. Wówczas zrodziła się myśl napisania takiego podręcznika i obecna
próba jest spełnieniem życzeń moich słuchaczy. Podręcznik powstał
częściowo na podstawie wykładów, jakie miałem dla studentów drugiego
roku Wydziału Lekarskiego Akademii Medycznej w Łodzi w latach
1963—1972. Ze względu na swój charakter nie może on służyć do
systematycznych i gruntownych studiów przedmiotu. Rolę tę spełniają
obszerne polskie podręczniki fizjologii, jak i tłumaczenia z języków
obcych. Celem moim było omówienie podstawowych praw fizjologicznych z pominięciem opisu metod, za pomocą których zostały one odkryte.
Pominąłem również wiadomości z nauk pokrewnych, to jest z anatomii,
histologii, biofizyki, biochemii i fizjologii porównawczej, sprowadzając
niektóre do niezbędnego minimum.
W tekście po polskich nazwach struktur anatomicznych podane są
w nawiasach nazwy łacińskie złożone kursywą. Pewną innowacją natomiast, nie praktykowaną w podręcznikach dla studentów, jest umieszczenie w nawiasach po niektórych polskich terminach z zakresu fizjologii ich odpowiedników angielskich. Podałem również pełne brzmienie
nazw angielskich po symbolach utworzonych z pierwszych ich liter.
Wprowadziłem to w celu ułatwienia polskiemu czytelnikowi korzystania
z obcego piśmiennictwa. Mam nadzieję, że podręcznik ten zostanie przez
studentów zaaprobowany i że spełni swoje zadanie. Będę bardzo zobowiązany za wszelkie uwagi.
Pragnę w tym miejscu wyrazić swoje najserdeczniejsze podziękowania
tym wszystkim, którzy służyli mi radą lub pomocą w przygotowaniu
podręcznika do druku, przede wszystkim Panu Profesorowi Julianowi
8
Słowo wstępne do pierwszego wydania
Walawskiemu, Panu Profesorowi Bronisławowi Filipowiczowi i Panu
Docentowi Janowi Guzkowi za zapoznanie się z całością książki i za
cenne uwagi. Z poszczególnymi rozdziałami zapoznali się i przekazali
mi swoje uwagi: Pan Profesor Andrzej Trzebski — „Prawa fizjologiczne
organizmu człowieka" i „Układ krążenia", Pan Docent Bohdan Lewartowski — „Czynność komórek nerwowych i mięśniowych" i „Serce",
Pan Docent Juliusz Narębski — „Czucie i ruch", Pan Docent Rudolf
KHmek — „Kontrola środowiska wewnętrznego", Pani Docent Zdzisława
Traczyk — „Krew i hemopoeza", Pan Docent Antoni Koziorowski —
„Oddychanie", Pan Profesor Stanisław Konturek — „Odżywianie" i Pan
Docent Marian Wojnarowski — „Płyny ustrojowe i czynność nerek".
Wszystkim składam wyrazy wdzięczności za poniesiony trud i wnikliwe
uwagi. Składam również podziękowanie Panu Profesorowi Hansowi
Schaeferowi, przewodniczącemu Komisji Mianownictwa Międzynarodowej Unii Nauk Fizjologicznych za udostępnienie mi wykazu mianownictwa fizjologicznego.
Łódź, dnia 18 lutego 1972 roku
Autor
Spis treści
1. Prawa fizjologiczne organizmu człowieka ......................................
17
Charakterystyka przedmiotu ................................................................
Środowisko biologiczne organizmu...............................................
Homeostaza....................................................................................
Czynność komórki................................................................................
Błona komórkowa .........................................................................
Jądro komórkowe .........................................................................
Siateczka śródplazmatyczna ziarnista ..........................................
Siateczka śródplazmatyczna gładka ..............................................
Lizosomy........................................................................................
Mitochondria .................................................................................
Metabolizm wewnątrzkomórkowy ........................................................
Samoregulacja metabolizmu komórkowego .................................
Cykl komórkowy i śmierć komórki ..............................................
Kontrola i regulacja funkcji..................................................................
Specjalizacja funkcji w organizmie ..............................................
Przetwarzanie i przenoszenie informacji ........................................
Komunikacja pomiędzy komórkami ..............................................
Sprzężenie zwrotne ........................................................................
17
18
22
22
22
34
37
42
42
44
45
48
48
49
49
51
53
56
2. Czynność komórek nerwowych i mięśniowych .............................
59
Pobudliwość i pobudzenie ...................................................................
Potencjał spoczynkowy błony komórkowej ..................................
Komórka nerwowa ...............................................................................
Potencjał czynnościowy .................................................................
Synapsy ..........................................................................................
59
60
62
66
73
1O
Spis treści
Kontrola ekspresji genów ..................................................................
Hamowanie presynaptyczne .................................................................
Przewodzenie impulsów we włóknach bezrdzennych .
. . .
Przewodzenie impulsów we włóknach rdzennych . . . .
Grupy włókien nerwowych ..................................................................
Nerwy czas zk owe i nerwy rdzeni owe ....................................................
Komórki glejowe ..........................................................................................
Mięśnie poprzecznie prążkowane .................................................................
Komórka mięśniowa ............................................................................
Skurcze mięśni .......................................................................................
Regulacja napięcia mięśniowego ..........................................................
Synapsa nerwowo-mięśniowa ...............................................
Mięśnie gładkie ..............................................................................................
Mięsień sercowy ..............................................................................................
Komórki układu przewod zą ce go serca .............................................
81
82
82
84
86
87
88
89
89
95
100
102
106
110
113
3. Czucie i ruch ......................................................................................
115
Czynność bioelektryczna mózgu spontaniczna ..................................
Odruchy.................................................................................................
Łuk odruchowy ..............................................................................
Ośrodki rdzenia kręgowego ............................................................
Czucie ....................................................................................................
Czucie teleceptywne...............................................................................
Zmysł powonienia .........................................................................
Zmysł wzroku ...............................................................................
Zmysł słuchu ................................................................................
Czucie eksteroceptywne ........................................................................
Czucie dotyku i ucisku ........................................................................
Czucie ciepła i zimna...................................................................
Czucie bólu powierzchownego .....................................................
Czucie smaku ................................................................................
Czucie proprioceptywne ........................................................................
Czynność błędnika ........................................................................
Czucie interoceptywne .........................................................................
Czucie bólu trzewnego ..................................................................
Drogi nieswoiste dla czucia i percepcji ..............................................
Ruchy i postawa ciała .........................................................................
Czynność układu pozapiramidowego ...........................................
Czynność móżdżku ........................................................................
Układ siatkowaty pnia mózgowia ...............................................
116
121
122
124
129
131
131
133
150
157
159
162
162
167
168
170
174
175
178
180
185
187
190
Spis treści
11
Czuwanie i sen.......................................................................................
Ośrodki kierujące zachowaniem ...........................................................
Ośrodki motywacyjne.....................................................................
Układ limbiczny ............................................................................
Uczenie się i zapamiętywanie..................................................................
Pamięć trwała .................................................................................
Zespoły czynnościowe ośrodkowego układu nerwowego .
.
.
197
201
201
203
204
205
210
4. Kontrola środowiska wewnętrznego .................................................
213
Zachowanie równowagi homeostatycznej ........................................
Układ nerwowy autonomiczny .............................................................
Część współczulna układu autonomicznego ..................................
Część przywspółczulna układu autonomicznego ..............................
Działanie na efektory impulsacji współczulnej i przywspółczulnej
Czynność gruczołów dokrewnych .......................................................
Hormony podwzgórzowe...............................................................
Przysadka — część gruczołowa .....................................................
Przysadka — część pośrednia ......................................................
Kora i rdzeń nadnerczy ..................................................................
Gruczoł tarczowy ...........................................................................
Jajniki .............................................................................................
Jądra . . . . ' . ................................................................
Gruczoły przytarczyczne ...............................................................
Wyspy trzustkowe ..........................................................................
Szyszynka .......................................................................................
Grasica ...........................................................................................
Hormony tkankowe ...............................................................................
Czynniki wzrostowe .......................................................................
Termoregulacja ......................................................................................
213
214
215
219
222
225
227
235
241
242
249
255
257
260
261
263
264
265
268
272
5. Krążenie ................................................................................................
276
Krew ......................................................................................................
Rola krwi w organizmie..................................................................
Czynność tkanek krwiotwórczych ........................................................
Szpik kostny..................................................................................
Grasica ...........................................................................................
Węzły chłonne i grudki chłonne ....................................................
Śledziona ........................................................................................
Elementy morfotyczne krwi ...................................................................
276
277
277
278
294
296
297
298
12
Spis treści
Erytrocyty ......................................................................................
Leukocyty.......................................................................................
Trombocyty ....................................................................................
Osocze....................................................................................................
Białka osocza ................................................................................
Składniki organiczne pozabiałkowe osocza ..................................
Lipidy osocza .................................................................................
Hemostaza ......................................................................................
Chłonka .................................................................................................
Układ sercowo-naczyniowy ..................................................................
Serce.......................................................................................................
Czynność bioelektryczna serca — EKG ........................................
Czynność mechaniczna serca ......................................................
Zjawiska akustyczne ....................................................................
Krążenie wieńcowe.........................................................................
Krążenie duże.......................................................................................
Zbiornik tętniczy duży ..................................................................
Zbiornik żylny duży ........................................................................
Krążenie krwi w naczyniach włosowatych ..................................
Krążenie chłonki ..........................................................................
Ośrodki kontrolujące krążenie krwi .....................................................
Ośrodek sercowy ...........................................................................
Ośrodek naczynioruchowy ............................................................
Regulacja ciśnienia tętniczego krwi ..............................................
Miejscowa regulacja przepływu krwi .....................................................
Krążenie płucne .....................................................................................
Krążenie krwi w mózgowiu ..................................................................
Krążenie wrotne .....................................................................................
298
306
318
319
321
325
325
326
333
334
336
336
348
356
357
360
360
368
371
373
375
375
378
382
386
389
392
394
6. Oddychanie ..........................................................................................
396
Oddychanie zewnętrzne .........................................................................
Wentylacja płuc ..............................................................................
Dyfuzja gazów w płucach..............................................................
Transport gazów ...........................................................................
Dyfuzja gazów w tkankach ........................................................
Regulacja oddychania ...........................................................................
Ośrodek oddechowy ......................................................................
Oddychanie wewnętrzne........................................................................
Przemiana pośrednia w komórkach ..............................................
396
397
403
405
409
410
410
415
415
Spis treści
13
Przemiana w organizmie .....................................................................
Podstawowa przemiana materii .....................................................
418
419
7. Układ trawienny .................................................................................
423
Odżywianie
....................................................................................
Równowaga energetyczna .............................................................
Trawienie pokarmów ............................................................................
Jama ustna i przełyk ...................................................................
Żołądek ..........................................................................................
Jelito cienkie ..................................................................................
Trawienie w jelicie cienkim ...........................................................
Wydzielanie soku trzustkowego .....................................................
Wydzielanie żółci ...........................................................................
Jelito grube ....................................................................................
Hormony żołądkowo-jelitowe ..............................................................
Wchłanianie w przewodzie pokarmowym ..............................................
Węglowodany .................................................................................
Tłuszcze ..........................................................................................
Białka i kwasy nukleinowe............................................................
Witaminy, sole mineralne i woda ...............................................
Czynność wątroby .................................................................................
Czynność zewnątrzwydzielnicza .....................................................
Czynność wewnątrzwydzielnicza.....................................................
Magazynowanie..............................................................................
Regulacja czynności ........................................................................
423
423
425
425
428
437
440
441
445
447
450
455
456
456
460
461
463
466
466
466
468
8. Płyny ustrojowe i czynność nerek ...................................................
469
Płyny i elektrolity ustrojowe ..............................................................
Woda ..............................................................................................
Elektrolity
..............................................................................
Utrata wody i elektrolitów............................................................
Czynność nerek .....................................................................................
Przepływ krwi ................................................................................
Resorpcja i sekrecja kanalikowa .................................................
Regulacja tworzenia się moczu .....................................................
Czynności wewnątrzwydzielnicze..................................................
Wydalanie moczu .................................................................................
469
469
471
473
474
476
482
490
492
494
14
Spis treści
9. Rozród ...................................................................................................
498
Determinacja pici ...........................................................................
Rozród u płci męskiej...........................................................................
Spermatogeneza ..............................................................................
Rozród u płci żeńskiej .........................................................................
Cykl płciowy żeński ........................................................................
Ciąża...............................................................................................
Poród ..............................................................................................
Połóg i laktacja .............................................................................
498
499
499
501
501
507
509
512
Skróty i symbole powszechnie używane w fizjologii
.
.
.
.
514
S k r ó t y a m i n o k w a s ó w w y s t ę p u j ą c y c h w b i a ł k a c h ................................
527
Mianownictwo steroidów
................................................................
528
Skorowidz ..................................................................................................
529
Podstawowe jednostki miar
15
Tabela I
Podstawowe jednostki miar międzynarodowego układu SI, jednostki pochodne* i jednostki dopuszczone do stosowania w medycynie** na podstawie uchwały Sesji Plenarnej
Światowej Organizacji Zdrowia z maja 1977 roku
*** Skrót jednostki objętości w postaci wielkiej litery L został wprowadzony do
piśmiennictwa medycznego w ostatnich latach z inicjatywy Redaktora Naczelnego czasopisma The Journal of Americal Medical Association. (G. D. Lundberg. Editorial. SI unit
irńplementation — the next step. JAMA, 1988; 260, nr 1, 73—76.)
16
Wielokrotności i podwielokrotności jednostek miar
Tabela II
Wielokrotności i podwielokrotności jednostek miar
Tabela III
Alfabet grecki*
* Transkrypcja nazw liter greckich jest zgodna z używaną w medycznej informacji
naukowej posługującej się językiem angielskim.
1
Prawa fizjologiczne
organizmu człowieka
I Charakterystyka przedmiotu
Fizjologia człowieka jest nauką o czynności żywego organizmu. Stanowi ona zbiór praw fizjologicznych (physiologic principles), jakim podlega
cały organizm oraz poszczególne jego układy, narządy, tkanki i komórki.
Prawa rządzące żywym organizmem są wykrywane doświadczalnie. Określają one zarówno warunki, w jakich przebiega prawidłowa czynność, jak
i mechanizmy fizjologiczne zapewniające prawidłową czynność całego
organizmu i jego poszczególnych komórek.
Warunki, w jakich żyje każdy organizm zwierzęcy, a więc i organizm
człowieka, wyznaczone są przez czynniki fizyczne i chemiczne stanowiące środowisko biologiczne (biologie environment). W środowisku tym
obowiązują prawa fizyczne i chemiczne, a w stosunku do organizmów
żywych również prawa biologiczne. Człowiek żyje w społeczeństwie i jest
wytworem życia społecznego, dlatego też odnoszą się do czynności jego
organizmu również prawa społeczne.
Mechanizmy fizjologiczne (physiologic mechanisms), dzięki którym
procesy życiowe w organizmie przebiegają prawidłowo, występują jednocześnie na wielu poziomach organizacji. Życie pojedynczej komórki
zależne jest od optymalnego działania wewnątrzkomórkowych mechanizmów fizjologicznych. Życie wielkiego zbiorowiska komórek, jakim
jest organizm, uwarunkowane jest sprawnym działaniem licznych mechanizmów fizjologicznych występujących pomiędzy tkankami, narządami
i całymi układami.
18
Prawa fizjologiczne organizmu człowieka
Istota procesów fizjologicznych
Wszystkie istoty żywe zarówno o prostej budowie — jednokomórkowce, jak i o złożonej budowie, np. organizm człowieka, charakteryzuje wspólna właściwość, którą jest przemiana materii — metabolizm.
Metabolizm (metabolism) obejmuje dwa przeciwne procesy, z których
jeden anabolizm (anabolism) — czyli asymilacja, przyswajanie — polega
na gromadzeniu energii w organizmie żywym, drugi zaś katabolizm
(catabolism) — czyli dysymilacja, rozpad — związany jest ze zmniejszaniem się zapasu tej energii. W warunkach fizjologicznych oba procesy
zachodzą jednocześnie.
W okresie wzrostu organizmu anabolizm przeważa nad katabolizmem,
a po osiągnięciu dojrzałości metabolizm wykazuje równowagę energetyczną
(energy balance).
W dojrzałym organizmie wahania w kierunku przewagi anabolizmu
lub katabolizmu stale zachodzą cyklicznie, ale nie przekraczają pewnych
granic, które można określić jako fizjologiczne granice dla procesów
życiowych. Przekroczenie tych granic w kierunku gromadzenia się albo
ubywania energii z organizmu lub poszczególnych jego układów i narządów stanowi przejście od prawidłowo przebiegających procesów życiowych — fizjologicznych, do nieprawidłowych — patologicznych.
W konsekwencji organizm traci optymalne warunki do życia w środowisku
biologicznym i szybciej lub wolniej obumiera.
I
Środowisko biologiczne
organizmu
Metabolizm jest podstawowym warunkiem życia biologicznego żywego
organizmu. Przebiega on wewnątrz organizmu, czyli w jego środowisku
wewnętrznym (internal environment). Każdy żywy organizm jest otoczony
przez środowisko zewnętrzne (external environment), które choć nie bierze
bezpośredniego udziału w metabolizmie komórkowym, to jednak wywiera
nań wielki wpływ.
W organizmie człowieka środowisko wewnętrzne od zewnętrznego stale
oddziela co najmniej jedna warstwa komórek. Do środowiska zewnętrznego zalicza się treść wypełniającą przewód pokarmowy oraz powietrze
w drogach oddechowych i w pęcherzykach płucnych.
Środowisko biologiczne organizmu
19
Przestrzenie płynów ustrojowych
Woda (water) jest podstawowym składnikiem organizmu wszystkich
ssaków (mammals), w tym również człowieka {homo sapiens). U noworodka ilość całkowitej wody organizmu — TBW (total body water) — wynosi
ponad 80%. W miarę wzrostu i dojrzewania organizmu procent wody
zmniejsza się i ustala na okres kilkudziesięciu lat życia w przedziale wieku
od lat około 18 do około 40. U mężczyzny w tym przedziale wieku
całkowita woda organizmu (TBW) stanowi przeciętnie około 62% masy
ciała. Pozostałe 38% masy ciała przypada na białka (około 18%), tłuszcze
(około 13%) i sole mineralne (około 7%). U kobiet ilość całkowitej wody
organizmu (TBW) jest od kilku do dziesięciu procent mniejsza w porównaniu do TBW u mężczyzn, w tym samym wieku, z uwagi na większy
u kobiet procentowy udział tkanki tłuszczowej w masie ciała.
Komórki (cellulae) i ich błony komórkowe dzielą wodę organizmu na
trzy przestrzenie płynów ustrojowych (body fluid compartments). Są to:
• płyn w przestrzeni wewnątrzkomórkowej (intracellular fluid compartment);
0 płyn w przestrzeni zewnątrzkomórkowej (extracellular fluid compartment);
• płyn w przestrzeni transkomórkowej (transcellular fluid compartment).
Przestrzeń wewnątrzkomórkową wypełnia płyn wewnątrzkomórkowy —
ICF (intracellular fluid) stanowiący około 40% masy ciała mężczyzn
1 około 30% masy ciała kobiet w wieku od 18 do 40 lat.
Płyn zewnątrzkomórkowy — ECF (extracellular fluid) nie jest jednorodny pod względem składu i rozmieszczenia. Obejmuje on osocze krwi
(blood plasma), płyn tkankowy (interstitiał fluid) i chłonkę (lymph).
Płyn w przestrzeni transkomórkowej zaliczany jest również do płynu
zewnątrzkomórkowego. Tworzą go płyn mózgowo-rdzeniowy, płyn w komorach oka, maź stawowa, płyn surowiczy w jamie opłucnej, płyn
osierdziowy i soki trawienne. Niezależnie od miejsca występowania
zawartość składników nieorganicznych w ECF jest dość stała. Poszczególne płyny ustrojowe należące do ECF różnią się natomiast dość znacznie
pod względem zawartości niektórych składników organicznych.
Stężenie jonów (concentration of ions) zarówno w płynie zewnątrzkomórkowym, jak i wewnątrzkomórkowym przyjęto wyrażać w molach
(mol) w litrze roztworu zgodnie z Międzynarodowym Układem Jednostek
Miar (SI) lub biorąc pod uwagę ładunek elektryczny w ekwiwalentach
(Eq). Jeden ekwiwalent, czyli równoważnik (1 Eq), jest to jeden mol
20
Prawa fizjologiczne organizmu człowieka
kationów lub anionów danej substancji wyrażony w gramach i podzielony
przez wartościowość jonów.
1 mol kationów tub anionów
q
7 ,
wartościowość jonów
np.:
23 g
1 EqNa+ = ------ = 23 g.
Drugą jednostką, za pomocą której określa się właściwości płynów
ustrojowych, jest osmól. Jeden osmól, jest to jeden mol związku chemicznego lub jonów występujący w objętości 1 litra roztworu podzielony przez
liczbę cząsteczek uwolnionych z każdej cząsteczki danego związku i swobodnie poruszających się w roztworze wodnym.
1 mol w litrze roztworu
1 Osm = -------------------------------l.s.w. x wsp.
gdzie:
l.s.w. — liczba swobodnych cząsteczek uwolnionych z jednej cząsteczki związku chemicznego. Dla związków nie dysocjujących l.s.w. = 1. a dla dysocjujących l.s.w.> I; wsp.
— współczynnik osmotyczny. Jest bliski jedności dla związków nie dysocjujących.
np.:
60.06
I Osm dla mocznika = --------= 60.06 g w I L roztworu.
I xl
Osmolarność (osmolarity) płynu określa ciśnienie osmotyczne panujące
w 1 litrze roztworu, a więc mówi o liczbie swobodnie poruszających się
cząsteczek i wywieranym przez nie ciśnieniu na błony półprzepuszczalne.
do których należą również błony komórkowe.
Związki dysocjujące, występujące w płynach ustrojowych, uwalniają
różną liczbę swobodnych cząsteczek (l.s.w.) i mają odmienne współczynniki osmotyczne. Ciśnienie osmotyczne płynów ustrojowych oznacza się
na podstawie pomiaru obniżenia punktu zamarzania roztworu. Obniżenie
punktu zamarzania o 1,86°C odpowiada ciśnieniu osmotycznemu równemu 1 osmolowi (Osm).
1 Osm obniża temperaturę zamarzania o - 1,86°C.
Z uwagi na małe stężenie związków chemicznych występujących
w płynach ustrojowych w fizjologii używane są jednostki tysiąc razy
mniejsze od ekwiwalentu i osmolą.
1 milirównoważnik = 1 mEq 1
miliosmol = 1 mOsm
__
Środowisko biologiczne organizmu
21
Punkt zamarzania osocza ludzkiego wynosi — 0,558°C (SI: 272,562 K),
co odpowiada osmolamości osocza równej 300 mOsm (0,558/0,00186),
mocz zaś zamarza w temperaturze np. — 2,232°C (SI: 270,948 K), co
odpowiada 1200 mOsm (2,232/0,00186).
W płynie zewnątrzkomórkowym występuje stosunkowo duże stężenie
jonów sodowych i chlorkowych oraz małe stężenie jonów potasowych
(tab. 1).
Tabela 1
Kationy i aniony w osoczu, w płynie tkankowym i w płynie wewnątrzkomórkowym
Uwaga: jony wymienione w tabeli nie są przedstawione tak, jak tego wymaga układ SI,
ponieważ z obu stron błony komórkowej równoważą się ładunki elektryczne wyrażone
w mEq/L H 2 O, a nie ilości substancji w mmol/L H 2 O.
• Zmienne stężenie zależne od stanu czystościowego komórek mięśniowych.
W płynie wewnątrzkomórkowym np. komórek mięśni poprzecznie
prążkowanych stężenie jonów sodowych i chlorkowych jest niewielkie,
natomiast jony potasowe i magnezowe występują w stosunkowo dużym
stężeniu. Mimo różnic w stężeniu poszczególnych składników pomiędzy
płynem zewnątrz- i wewnątrzkomórkowym ciśnienie osmotyczne obu tych
płynów jest jednakowe, czyli izoosmotyczne (izotoniczne).
22
Prawa fizjologiczne organizmu człowieka
I Homeostaza
Właściwością organizmu ssaków jest zdolność do zachowania stałego
środowiska wewnętrznego pomimo zmieniającego się środowiska zewnętrznego. Ta zdolność organizmu nazywana jest homeostazą (homeostasis).
Ze środowiska zewnętrznego działa na organizm wiele czynników
fizycznych, np. wzrost lub obniżenie temperatury otoczenia, promieniowanie cieplne itp., oraz czynników chemicznych, np. jony nieorganiczne
zawarte w pokarmach. Czynniki fizyczne i chemiczne mogą wywołać
znaczne przesunięcia w rozmieszczeniu wody w organizmie, jak również
istotnie zmienić stężenie poszczególnych jonów w komórkach i płynach
ustrojowych. W warunkach zachowanej homeostazy nie dochodzi jednak
do znacznych przesunięć w rozmieszczeniu wody i zmian w stężeniu
jonów. Dzięki wzajemnemu oddziaływaniu komórek, tkanek, narządów
i układów oraz działaniu odpowiednich mechanizmów fizjologicznych nie
występują istotne zmiany w środowisku wewnętrznym organizmu.
Zawartość wody w komórkach i stężenie poszczególnych jonów w płynach ustrojowych oscylują w pewnym zakresie wokół określonych wielkości fizjologicznych. Im mniejsze są te oscylacje, wywołane zmianami
w środowisku zewnętrznym, tym lepiej zachowane jest środowisko wewnętrzne organizmu, czyli tym lepsza jest homeostaza.
I Czynność komórki
Wszystkie tkanki, narządy i układy w organizmie człowieka utworzone
są z podstawowych jednostek czynnościowych jakimi są komórki (cellulae), które bez względu na stopień zróżnicowania cechuje metabolizm
i biosynteza. Są one zbudowane z cytoplazmy i struktur cytoplazmatycznych oraz z jądra komórkowego. Do struktur cytoplazmatycznych (organełłae cytoplasmaticae) zalicza się: błonę komórkową, siateczkę śródplazmatyczną, rybosomy, aparat Golgiego, mitochondria i lizosomy.
I Błona komórkowa
Od otoczenia odgranicza komórkę błona komórkowa. Podobne błony
otaczają większość struktur komórkowych, wydzielając je od cytoplazmy.
^
Czynność komórki
23
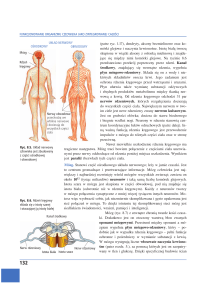
Ryc. 1. Budowa schematyczna komórki i struktur komórkowych. (Reprodukowano za
zezwoleniem Michała Karaska.)
Błona komórkowa {cytolemma) o grubości 7,4—10 nm, zgodnie z hipotezą
Singera i Nicolsona, ma płynną mozaikową strukturę (fluid mosaic
structure). Składa się z dwóch warstw cząsteczek fosfolipidów ustawionych
długą osią prostopadle do powierzchni błony. Hydrofobowe bieguny
Hpidowe obu warstw skierowane są do siebie, do wewnątrz błony, bieguny
fosfatydowe, hydrofilne zaś — od siebie, na zewnątrz błony.
W błonie komórkowej jak gdyby pływają białka globularne (globular
units), zajmujące warstwę zewnętrzną lub wewnętrzną fosfolipidową, jak
również całą grubość błony.
Białka błony komórkowej zazwyczaj zbudowane są z kilku podjednostek i można je podzielić na pięć grup w zależności od spełnianych przez
nie funkcji:
24
Prawa fizjologiczne organizmu człowieka
• białka integralne (structural proteins), których bieguny hydrofilne są
ustawione na zewnątrz błony;
• białka nośnikowe (transporting proteins) tworzące kanały aktywnego
transportu cząsteczek przez błonę komórkową;
• białka tworzące kanały jonowe (channel proteins);
• białka receptorowe (receptor proteins) wiążące swoiście cząsteczki
chemiczne, zwane Ugandami (ligants), takie jak przekaźniki chemiczne
i inne substancje występujące w płynie zewnątrzkomórkowym.
Ryc. 2. Schemat budowy błony komórkowej. ] — podwójna warstwa cząsteczek fosfolipidów, 2 - - białka integralne, 3 — białka w warstwie wewnętrznej fosfolipidów, 4 — białka
w warstwie zewnętrznej fosfolipidów, 5 — białka powierzchniowe od strony cytoplazmy.
6 — białka kanałowe- ECF — płyn zewnątrzkomórkowy, ICF — płyn wewnątrzkomórkowy.
Błony otaczające jądro komórkowe (nucleomembrand), siateczkę śródplazmatyczną {reticulum endoplasmaticum), lizosomy i mitochondria zbliżone są pod względem budowy do błony komórkowej. Różnią się od niej
liczbą warstw cząsteczek fosfolipidów oraz jakością cząsteczek białek
osadzonych na tych błonach.
Transport przez błonę komórkową zależy od masy cząsteczkowej,
właściwości, średnicy i ładunku elektrycznego cząsteczek związków chemicznych. Cząsteczki rozpuszczalne w tłuszczach przenikają przez warstwy
fosfolipidowe błony komórkowej w obu kierunkach na zasadzie dyfuzji,
bez względu na ich średnicę. Takie właściwości mają cząsteczki O2, CO2,
Czynność komórki
25
kwasy tłuszczowe, steroidy i rozpuszczalniki organiczne (alkohole, etery
itp.). Dyfundują one zgodnie z gradientem stężenia, to znaczy przenikają
przez błonę komórkową zawsze od strony większego stężenia do mniejszego stężenia.
Ułatwioną dyfuzją (facilitated diffusion) nazywany jest transport błonowy, w którym dodatkowy czynnik wspomaga dyfuzję zgodnie z gradientem stężeń. Tym dodatkowym czynnikiem może być np. ujemny ładunek
elektryczny wewnątrz komórki ułatwiający dokomórkową dyfuzję jonów
o dodatnim ładunku elektrycznym.
-■
U
KJ
Ryn
zewn
komórkowy
(ECF)
Bfona
komórkowa
MCnosacharydy
Aminokw asy
Płyn .
wewnątrzkomórkowy
(ICF)
Ryc. 3. Przenikanie przez błonę komórkową cząsteczek na zasadzie dyfuzji (O 2, CO2)
i aktywnego transportu (monosacharydy i aminokwasy) dzięki białku nośnikowemu.
Związki nierozpuszczalne w tłuszczach są aktywnie transportowane
przez białka nośnikowe, tworząc z nimi nietrwałe kompleksy. Po przeniesieniu cząsteczek połączonych z białkiem nośnikowym przez błonę komórkową kompleks rozpada się, uwalniając cząsteczki do cytoplazmy. Tego
typu transport przez błonę komórkową, przeciw gradientowi stężenia,
nazywa się aktywnym transportem (active transport). W ten sposób
przenoszone są przez błonę komórkową cząsteczki monosacharydów
i aminokwasów.
W błonie komórkowej znajdują się pory o średnicy 800 pm, przez
które przenikają małe cząsteczki o ładunku obojętnym, np. cząstecz ki
wody i mocznika. Są w niej również swoiste kanały (specific chan-nels)
dla dokomórkowych i odkomórkowych prądów poszczególnych jonów.
26
Prawa fizjologiczne organizmu człowieka
Białka tworzą kanały jonowe biernie lub aktywnie transportują kationy
i aniony zgodnie lub przeciw gradientowi stężenia i ładunku elektrycznego. Aktywny transport i ułatwiona dyfuzja wymagają zużycia energii,
która czerpana jest głównie z rozpadu adenozynotrifosforanów (ATP),
w czym bierze udział adenozyn otrifosfataza (adenosinetriphosphatase) —
białko enzymatyczne błony komórkowej.
Uniportem nazywa się przenoszenie cząsteczek jednej substancji chemicznej przez białko nośnikowe do komórki. Symportem zaś jednoczesne
przenoszenie dwóch cząsteczek substancji chemicznych do komórki, np.
jony Na + i glukozy do enterocytów w jelicie cienkim.
Ryc. 4. Przenikanie przez błonę komórkową jonów o ładunku dodatnim. Po związaniu się
cząsteczek transmittera z białkiem receptorowym następuje zmiana konformacji białek
kanałowych i jony Na" przenikają do wnętrza komórki.
Antyport jest to jednoczesne przenoszenie przez białko nośnikowe
cząsteczek dwóch różnych substancji chemicznych, jednych na zewnątrz
i drugich do wewnątrz komórki. Transport błonowy typu antyportu
występuje w błonach komórkowych neuronów i komórek mięśniowych.
Białkiem dostarczającym energii dla transportu czynnego jest adenozynotrifosfataza aktywowana przez sód i potas (Na + -K+-ATP-aza). Hydroliza ATP do ADP i ortofosforanu katalizowana przez Na +-K+-ATP-azę
umożliwia transport jonów Na + z komórki na zewnątrz komórki i jonów
K+ z płynu zewnątrzkomórkowego do komórki.
Czynność komórki
27
Receptory błonowe
Bardzo ważną rolę w odbiorze informacji przez komórki spełniają
białka receptorowe. Syntetyzowane są w siateczce śródplazmatycznej
ziarnistej, tak jak wszystkie pozostałe białka występujące w błonie
komórkowej. Przyczepiają się do wewnętrznej powierzchni błony komórkowej i następnie w procesie eksternalizacji białek receptorowych (receptor externalization) przenoszone są na zewnętrzną powierzchnię. Po
związaniu się przekaźników chemicznych (transmittery, hormony i inne
substancje) występujących w płynie zewnątrzkomórkowym z receptorami
na zewnętrznej powierzchni błony komórkowej białka receptorowe oddziałują na inne białka błony komórkowej. Wówczas białka nośnikowe
przyspieszają lub zwalniają aktywny transport przez błonę komórkową,
otwierają się lub zamykają kanały dla dokomórkowych i odkomórkowych prądów jonowych oraz następuje aktywacja lub inaktywacja białek
enzymatycznych.
ICF
ECF
Ryc. 5. Białka kanałowe w Wonie komórkowej: a — imiport — przenoszenie jednej
substancji chemicznej do komórki; b — symport — przenoszenie jednocześnie dwóch różnych cząsteczek; c — antyport — przenoszenie jednocześnie dwóch różnych cząsteczek
w przeciwnych kierunkach, do komórki i z komórki.
Białka receptorowe nie tylko oddziałują na inne białka błony komórkowej, ale również same transportują cząsteczki chemiczne z płynu
zewnątrzkomórkowego do cytoplazmy. Cząsteczki insuliny, białka wytwarzanego przez komórki B wysp trzustkowych (patrz str. 262), po związaniu się z receptorami insulinowymi na zewnętrznej powierzchni błony
28
Prawa fizjologiczne organizmu człowieka
Ryc. 6. Schemat budowy błony komórkowej i białek błony komórkowej z przyłączonymi
cząsteczkami oligosacharydów. (Reprodukowano za zezwoleniem z W. Sawicki Histologia,
II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
komórkowej dostają się do wnętrza komórki w procesie internalizacji
receptorów (receptor internalization).
Receptory błonowe mają zdolność do swoistego wiązania się (specific
binding site) z określonymi substancjami chemicznymi, czyli Ugandami,
endogennymi — występującymi w organizmie, jak i egzogennymi — wprowadzonymi z zewnątrz. W błonie komórkowej występują białka receptorowe o największym powinowactwie do licznych substancji. Aktywne
substancje chemiczne występujące w płynie zewnątrzkomórkowym konkurują ze sobą o miejsce wiązania się z receptorem (competitive binding
site). Z kilku substancji chemicznych o podobnej budowie wiążą się
z receptorem cząsteczki tej substancji, która ma największe powinowactwo do tego receptora. Może być niewiele receptorów wiążących jedną
substancję, w stosunku do jednostki powierzchni błony komórkowej
Czynność komórki
29
i nawet przy małym stężeniu cząsteczek w płynie zewnątrzkomórkowym
wszystkie receptory mogą być połączone z ligandem. Dalsze zwiększenie
stężenia cząsteczek substancji w płynie zewnątrzkomórkowym nie będzie
miało wpływu na jej działanie na komórkę z uwagi na małą pojemność
wiązania (capacity binding site).
Ryc. 7. Związanie się transmittera z receptorem powoduje otwieranie się kanałów błonowych i aktywację białek enzymatycznych. Zablokowanie receptorów zmienia właściwości
błony komórkowej, która staje się niewrażliwa na transmitter. ECF — płyn zewnątrzkomórkowy, ICF — płyn wewnątrzkomórkowy.
Po zadziałaniu na błonę komórkową substancji egzogennych blokujących określone receptory (blockers) przestają one wiązać się z substancjami endogennymi. W ten sposób receptory w błonie komórkowej zostają
na krótszy lub dłuższy czas zablokowane i komórka przestaje odbierać
informacje za pośrednictwem określonego receptora. Biologicznie aktywne
substancje egzogenne wykazujące powinowactwo do receptorów błonowych są antagonistami określonych substancji endogennych.
Odbiór informacji przez komórki
Informacja odbierana przez receptory w błonie komórkowej wywołuje
reakcje:
• krótkotrwałe, rzędu od ułamka milisekundy do kilkudziesięciu mili-
30
Prawa fizjologiczne organizmu cz/owieka
sekund, związane z ruchem jonów przez błonę komórkową, z czym
wiąże się z kolei występowanie czynnościowego potencjału elektrycznego;
• dłużej trwające, od milisekund do wielu godzin, w wyniku aktywacji
lub inaktywacji enzymów w błonie komórkowej i wzmożenia lub
osłabienia metabolizmu komórkowego;
• długotrwałe, od wielu godzin do wielu tygodni, kiedy dochodzi do
ekspresji genów w jądrze komórkowym, w których zakodowana jest
informacja genetyczna dla syntezy enzymów zmieniających metabo
lizm komórkowy.
Odbieranie i przetwarzanie informacji przez błonę komórkową przebiega w trzech etapach:
• rozróżnianie sygnałów (discrimination) — informację odbieraną stano
wią sygnały, czyli cząsteczki przekaźników chemicznych, działające
niezależnie od siebie na błonę komórkową. Receptory błonowe odróż
niają cząsteczki na nie działające, np. cząsteczki acetylocholiny od
cząsteczek noradrenaliny;
• przenoszenie sygnałów (transduction) — białka receptorowe związane ze
swoistymi przekaźnikami chemicznymi oddziałują na białka kanałów
jonowych i białka enzymatyczne;
• wzmacnianie sygnałów (amplification) — sygnały odebrane przez ko
mórkę, czyli na wejściu układu, są zazwyczaj zbyt słabe, aby mogły
wpływać istotnie na metabolizm wewnątrzkomórkowy. Muszą zostać
odpowiednio wzmocnione. Takimi uniwersalnymi wzmacniaczami licz
nych sygnałów odbieranych przez komórki są enzymy wytwarzające
drugi wewnątrzkomórkowy przekaźnik chemiczny.
Drugi przekaźnik wewnątrzkomórkowy
Krążące w płynach ustrojowych fizjologicznie aktywne, endogenne
substancje chemiczne, przede wszystkim hormony, stanowią pierwszy
przekaźnik informacji (first chemical messenger). Po odebraniu przez
receptory w błonie komórkowej informacji i jej przeniesieniu dochodzi do
aktywacji lub inaktywacji enzymów i do zwiększenia, lub zmniejszenia
w komórkach stężenia drugiego przekaźnika informacji (second chemical
messenger).
Do wewnątrzkomórkowych przekaźników informacji zalicza się: cykliczny adenozynomonofosforan, cykliczny guanozynomonofosforan, inozytolotrifosforan, diacyloglicerol i jony wapniowe.
Czynność komórki
31
Pod wpływem cyklazy adenylanowej, występującej po wewnętrznej
stronie błony komórkowej, odczepiają się pirofosforany (P2O*~) od
adenozynotrifosforanów i powstaje cykliczny adenozyno-3', 5'-monofostoran. czyli cykliczny AMP (cyclic adenosine-3', 5'-monophosphate —
cAMP). Stężenie powstających w komórce cząsteczek cyklicznego AMP
jest znacznie większe niż stężenie przekaźnika chemicznego działającego
na komórkę z zewnątrz. Cykliczny AMP wywołuje zwiększenie aktywności enzymu, fosforylazy glikogenowej i uwalniając glukozo-1-fosforan
przyspiesza metabolizm glukozy w komórkach. Za pośrednictwem cyklazy
adenylanowej i jej aktywacji lub inaktywacji działa większość hormonów
w organizmie, cykliczny AMP zaś jest najważniejszym „drugim przekaźnikiem" przenoszącym informację wewnątrz komórek. Tworzący się w
komórkach cykliczny guanozyno-3', 5'-monofosforan (cyclic guanosi-ne3', 5'-monophosphate — cGMP) z guanozynotrifosforanów pod wpłyTabela 2
Hormony działające na tkanki poprzez aktywację (+) lub inaktywację ( —) cyklazy
adenylanowej i tworzenie się pod jej wpływem cyklicznego AMP
Hormony
Tkanki i narządy
Adrenalina i noradrenalina
[+) wątroba ( + ) mięśnie szkieletowe i
mięsień sercowy ( + ) tkanka tłuszczowa ( +
) mózgowie ( + ) płuca (+) śledziona (+)
trzustka
Insulina
(—) wątroba ( —) mięśnie szkieletowe i
mięsień sercowy ( —) tkanka tłuszczowa
Glukagon
( + ) wątroba ( + )
mięsień sercowy
Hormon kortykotropowy (ACTH)
Hormon tyreotropowy (TSH)
Hormon gruczołów przytarczycznych
(PTH)
( + ) kora nadnerczy
( + ) tarczyca
( + ) tkanka kostna
Kalcytonina
( —) tkanka kostna
Wazopresyna (AVP)
Hormon luteinizujący (LH)
( + ) nerki
( + ) jajnik — ciałko żółte ( + ) jądra —
komórki śródmiąższowe
Prostaglandyny (PG)
( —) tkanka tłuszczowa
32
Prawa fizjologiczne organizmu człowieka
wem aktywacji cyklazy guanylanowej ma zazwyczaj działanie przeciwne
w stosunku do cyklicznego AMP.
W przenoszeniu sygnałów pomiędzy receptorem związanym z ligandem i enzymem pośredniczą białka błonowe G występujące w wewnętrznej
warstwie błony komórkowej. Ich właściwością jest zdolność do wiązania
się i hydrolizowania gua nożyno trifosforano w. Białka G składają się
z trzech podjednostek a, p i y. W zależności od typu białka G działają one
aktywująco lub inaktywująco na cyklazę adenylanową. Po odebraniu
sygnału od receptora w białku aktywującym Gs następuje zamiana
związanego z białkiem Gs guanozynodifosforanu (GDP) na guanozynotrifosforan (GTP), odłączenie się podjednostek |3 i y i połączenie podjednostki a z cyklazą adenylanową, pod której wpływem tworzy się cAMP.
Uwolnienie ligandu od receptora powoduje odwrotny proces — zamianę
związanego z białkiem Gs guanozynotrifosforanu (GTP) na guanozynodifosforan, odłączenie cyklazy adenylanowej od podjednostki a i ponowne
połączenie się ze sobą trzech podjednostek białka G s, czyli powrót do
stanu spoczynku.
Ryc. 8. Udział białka błonowego aktywującego Gs, składającego się z trzech podjednostek a,
P, y w aktywacji cyklazy adenylanowej: 1 — wiązanie się ligandu (L) z receptorem (R),
2 — wymiana guanozynodifosforanu (GDP) na guanozynotrifosforan (GTP), 3 — aktywacja cyklazy adenylanowej i tworzenie się cyklicznego adenozynomonofosforanu (cAMP)
z adenozynotri fosforanu (ATP), 4 — uwolnienie się ligandu (L) od receptora (R) powoduje wymianę GTP na GDP i inaktywację cyklazy adenylanowej.
Czynność komórki
33
Związanie się innego ligandu z innym receptorem wywołuje wytwarzanie się po wewnętrznej stronie błony komórkowej inozytołotrifosforanu
(inositol triphosphate) i diacyloglicerolu (diacylglycerol). Receptory związane z ligandem aktywują fosfolipazę C, która hydrolizuje 4, 5-difosforan fosfatydyloinozytolu do inozytołotrifosforanu (IP 3) i diacyloglicerolu
(DAG). W przenoszeniu sygnałów pomiędzy receptorem i fosfolipazą
C pośredniczy również białko błonowe G. Inozytolotrifosforan wywołuje
w komórce przepływ jonów wapniowych z siateczki śródplazmatycznej
do cytoplazmy, diacyloglicerol zaś aktywuje kinazę białkową C i zwiększa
metabolizm komórki.
Ryc. 9. Udział białka błonowego Gs składającego się z trzech podjednostek a, p, y w aktywacji fosfolipazy C: 1 — wiązanie się ligandu (L) z receptorem (R), 2 — wymiana
guanozynodifosforanu (GDP) na guanozynotrifosforan (GTP), aktywacja fosfolipazy
C (PLC), tworzenie się inozytołotrifosforanu (IP3) i diacyloglicerolu (DAG) z difosforanu
fosfatydyloinozytolu (PIP2).
Jony wapniowe uważane są również za „drugi przekaźnik" wewnątrzkomórkowej informacji. W płynie zewnątrzkomórkowym (ECF) jony
Ca2+ występują jako jony wolne lub związane z zewnętrzną powierzchnią
błony komórkowej. Do komórek napływają przez co najmniej trzy różne
kanały dla dokomórkowego ich prądu. Wolne jony Ca 2+ po wejściu
do komórki wiążą się z błoną komórkową po stronie wewnętrznej oraz
2 — Fizjologia człowieka
34
Prawa fizjologiczne organizmu człowieka
z błonami siateczki śródplazmatycznej i błonami mitochondrialnymi, jak
również z białkiem występującym w cytoplazmie — kalmoduliną, zbudowaną z 148 aminokwasów (16 700 m.cz.). Kompleks kalmodulina-jony
wapniowe przyłącza się do enzymów wewnątrzkomórkowych, przede
wszystkim do kinaz białkowych i aktywuje je. Po wejściu do komórek
jonów Ca2+ następuje przyspieszenie egzocytozy, w tym również uwalniania transmitterów z zakończeń komórek nerwowych.
I Jądro komórkowe
Jądro komórkowe (nucleus) oddziela od cytoplazmy podwójna błona
jądrowa (membrana nuclearis), w której występują pory o średnicy od
40 do 100 nm zazwyczaj zamknięte błonami. Przestrzeń znajdująca się
pomiędzy blaszkami błony jądrowej tworzy zbiorniki błony jądrowej
(cisternae caryothecae) i jednocześnie łączy się z przestrzenią wewnątrz
siateczki śródplazmatycznej.
W okresach pomiędzy podziałami komórki (interfaza) jądro komórkowe stanowią chromosomy zwinięte i ściśle do siebie przylegające.
W jądrach komÓFek człowieka występują 23 pary chromosomów: 22 pary
chromosomów somatycznych i 1 para chromosomów płciowych, łącznie
w każdej komórce 46 chromosomów.
Ryc. 10. Schemat cyklu komórkowego. Faza
Gj — przerwa, faza S — replikacja łańcuchów
DNA, faza G2 — interfaza, faza M — mitoza
i faza Go — spoczynek.
Przed podziałem komórki macierzystej na dwie komórki p otomne
w fazie S cyklu komórkowego następuje duplikacja chromosomów dzięki
wytworzeniu się podwójnych nici chromatydowych. Do każdej potomnej
komórki przesuwają się 23 pary chromosomów, czyli 46 chromosomów.
W stadium metafazy, w czasie podziału komórki, chromosomy są najlepiej widoczne, można je liczyć i identyfikować. Każdy z chromosomów
składa się z dwóch mci chromatydowych połączonych ze sobą za pomocą
Czynnoś
ć
komórki
35
Ryc. 11. Po lewej schemat podwójnej helisy łańcuchów DNA stanowiących fragment genu
oksytocyny od 319 do 345 pary zasad. Łańcuchy DNA są skręcone i jednocześnie przeplatają się wzajemnie tak, że każda kolejna jednostka pary zasad (np. 329) jest obrócona
0 180° w stosunku do pierwszej pary zasad <np. 319). Po prawej ten sam odcinek łańcuchów
DNA, zapisany zgodnie z sekwencją par zasad purynowych (adenina — A, guanina — G)
1 pirymidynowych (tymina — T, cytozyna — C), i odpowiadających im po transkrypcji
i translacji dziewięciu aminokwasom tworzącym cząsteczkę oksytocyny.
36
Prawa fizjologiczne organizmu człowieka
centromeru. W zależności od długości nici chromatydowych, miejsca ich
połączenia za pomocą centromeru i występowania prążków, chromosomy
komórek człowieka dają się identyfikować i dzielić na grupy, których
wyróżnia się siedem.
Nici chromatydowe w jądrze komórkowym zbudowane są z białek
jądrowych i łańcuchów kwasów deoksyrybonukleinowych (deoxyribonucleic acid — DNA) tworzących podwójną helisę DNA (double helical
DNA chains). Każda nić DNA jest łańcuchem polideoksyrybozylowym,
w którym cząsteczki rybozy połączone są wiązaniami diestrowymi.
Z cząsteczkami deoksyrybozy, obu łańcuchów tworzących podwójną
helisę DNA, łączy się jedna z dwóch zasad purynowych — adenina lub
guanina, lub jedna z dwóch zasad pirymidynowych — tymina lub
cytozyna. W podwójnej helisie DNA zasady purynowe połączone są
z zasadami pirymidynowymi wiązaniami wodorowymi zgodnie z regułą
komplementarności zasad. Wiązania wodorowe pomiędzy zasadami podwójnej helisy DNA nie są trwałe i ulegają rozerwaniu w czasie życia
komórki.
W fazie S cyklu komórkowego poprzedzającej podział komórki
w jądrze komórkowym następuje replikacja łańcuchów DNA (replication
of DNA chains). Każdy łańcuch DNA stanowi matrycę, na której zachodzi synteza komplementarnego łańcucha DNA. Po podziale komórki
jądra komórek potomnych zawierają nici chromatydowe o takiej samej
sekwencji zasad jak jądra komórki macierzystej. Dzięki temu komórki
potomne zawierają tę samą informację genetyczną zakodowaną w sekwencji zasad purynowych i pirymidynowych łańcuchów DNA. Trzy
kolejne zasady — triplcty (coding tripłets) — z każdego łańcucha DNA
stanowią jeden znak kodu genetycznego. Sekwencja tripletów w łańcuchach DNA warunkuje sekwencję aminokwasów w łańcuchach polipeptydowych, z których utworzone są białka. Przeniesienie informacji genetycznej z łańcucha DNA na łańcuchy polipeptydowe zachodzi dzięki
procesom transkrypcji i translacji.
Kontrola procesów zachodzących w komórce przez materiał genetyczny zawarty w jądrze komórkowym zapoczątkowana jest przez proces
transkrypcji. Na łańcuchach DNA zostają zsyntetyzowane komplementarne łańcuchy kwasów rybonukleinowych (ribonucleic acid — RNA), w
czym bierze udział enzym RNA polimeraza. W łańcuchach RNA
sekwencja zasad purynowych — adeniny i guaniny — oraz pirymidynowych — cytozyny i uracylu — jest komplementarna do nici matrycowej. Drugą nietranskrybowaną nicią DNA jest nić kodująca.
Wśród białek jądrowych, które stanowią trzy czwarte masy jądra
komórkowego, wyróżnia się białka histonowe, na których owinięte są
Czynność komórki
37
podwójne helisy DNA, i białka niehistonowe, których jest większość,
w tym białka enzymatyczne. Genom człowieka zawiera 3 miliardy
nukleotydów, w każdej komórce somatycznej zaś są dwie podwójne helisy
DNA, czyli 6 miliardów nukleotydów. W zróżnicowanych komórkach
somatycznych tylko nieznaczna liczba genów podlega ekspresji dzięki
transkrypcji matrycowego RNA na łańcuchach DNA, w czym bierze
udział RNA polimeraza. Na pozostałe odcinki łańcucha DNA oddziałuje
inny enzym — restryktaza, która przeciwdziała transkrypcji RNA i ekspresji genów.
W jądrze komórkowym na łańcuchach DNA syntetyzowane są trzy
rodzaje kwasów rybonukleinowych (RNA):
• matrycowy — mRNA (messenger RNA) o dużej cząsteczce zawiera
jącej od kilkuset do kilku tysięcy nukleotydów;
• transportujący — tRNA (transfer RNA) o mniejszej cząsteczce zbu
dowanej z 75—85 nukleotydów;
• rybosomalny — rRNA (ribosomal RNA).
W procesie transkrypcji powstają w jądrze komórkowym prekursory
mRNA, tRNA i rRNA. Cząsteczki pre-matrycowego RNA (pre-mRNA)
zawierają zarówno sekwencje nukleotydów kodujące, zwane egzonami,
jak i przedzielone niekodującymi sekwencjami nukleotydów, zwanymi
intronami. W procesie modyfikacji potranskrypcyjnej introny wycinane
są z pre-mRNA i egzony łączą się ze sobą w krótsze łańcuchy mRNA.
W jądrze komórkowym widoczne są struktury zwane jąderkami
(nucleoli). Są to skupienia kwasów rybonukleinowych — RNA, zsyntetyzowanych w jądrze i chwilowo tam zmagazynowanych, aż do czasu
ich przesunięcia do cytoplazmy.
I Siateczka śródplazmatyczna ziarnista
Siateczkę śródplazmatyczną ziarnistą {reticulum endoplasmaticum granulosum) stanowi błona tworząca kanaliki. Na zewnętrznej powierzchni
tej błony skierowanej do cytoplazmy znajdują się ziarnistości — rybosomy
{ribosoma) — o średnicy około 23 nm. W rybosomach zachodzi proces
translacji, czyli synteza łańcuchów polipeptydowych. Rybosomy przesuwają się po cząsteczkach matrycowego RNA i jak gdyby odczytują
trójki zasad — triplety, z których każda odpowiada jednemu określonemu aminokwasowi.
38
Prawa fizjologiczne organizmu człowieka
Cząsteczki transportującego RNA tworzą podwójny łańcuch z czterema
pętlami. Na jednym końcu tRNA znajduje się ortofosforan, na drugim
niezmienna sekwencja CCA (nukleotydy: cytydylowy, cytydylowy, adenilowy). Wytworzony w jądrze komórkowym transportujący RNA trafia do
cytoplazmy, gdzie koniec cząsteczki tRNA, zawierającej nukleotyd adenilowy, przyłącza jeden określony aminokwas, wraz z którym wędruje do
rybosomów. Aminokwas przyłączany jest do końca 3' cząsteczki tRNA.
Natomiast w drugiej pętli tRNA występują nukleotydy, z których trzy
są komplementarne w stosunku do odpowiedniego kodonu w matrycowym RNA i stanowią antykodnn.
Synteza białka — kod genetyczny
Inicjacja translacji, czyli początek syntezy polipeptydu, zaczyna się
w rybosomie od kontaktu matrycowego RNA z transportującym RNA
przenoszącym aminokwas — metioninę. Kolejne cząsteczki transportującego RNA dostarczają aminokwasów, które łączą się wiązaniami
peptydowymi w odpowiedniej kolejności, zgodnie z kodonem genetycznym zawartym w matrycowym RNA i w ten sposób wydłużają się łańcuchy
polipeptydowe. Proces ten nosi nazwę elongacji i kończy się tenninacją
translacji wtedy, kiedy rybosom przesunie się na kodon terminacyjny
w matrycowym RNA. W stosunku do kodonu terminacyjnego brakuje
transportującego RNA z komplementarnym antykodonem.
Zsyntetyzowane w rybosomach polipeptydy podlegają dalszej potranslacyjnej enzymatycznej modyfikacji (posttranslational enzymatic modification), w wyniku której powstaje:
• białko strukturalne wchodzące w skład organelli cytoplazmatycznych;
• białko enzymatyczne, z którym związany jest metabolizm wewnątrz
komórkowy;
• białko wydzielane przez komórki na zewnątrz.
Białko zsyntetyzowane w rybosomach przed wydzielaniem na zewnątrz
komórki gromadzi się w kanalikach siateczki śródplazmatycznej, następnie przesuwane jest do aparatu Golgiego (complexus Golgiensis), w którym ulega zagęszczeniu i zostaje otoczone błoną. W postaci pęcherzyków
kierowane jest ku powierzchni komórki. Po zetknięciu się z błoną
komórkową pęcherzyki otwierają się i opróżniają z zawartości na zewnątrz komórki.
Powstające w komórkach białka wydzielane na zewnątrz stanowią
podstawowy składnik płynów ustrojowych i soków trawiennych. Są one
Czynność komórki
39
również nośnikiem informacji w organizmie jako hormony białkowe
i polipeptydowe (patrz str. 225—241), jak również modulatory synaptyczne (patrz str. 79).
Polipeptydy w czasie potranslacyjnej enzymatycznej modyfikacji mogą
być dęte na krótsze polipeptydy o łańcuchach różnej długości. Na przykład gen oksytocyny (patrz str. 40) zbudowany jest z 1055 nukleotydów.
Gen ten składa się z części promotorowej, trzech egzonów, dwóch
intronów, i części końcowej.
Błona ziarnistej
siateczki środplazma tycznej
Syn teiyzowony
peptyd
I
Oligosacharyd
Ryc. 12. Schemat syntezy potipeptydów na rybosomach ziarnistej siateczki śródplazmatycznej. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo
Lekarskie PZWL, Warszawa 1997.)
Zsyntetyzowany w procesie translacji łańcuch polipeptydowy ma 125
aminokwasów. Jest to pre-pro-hormon, czyli pre-pro-oksytocyno-neurofizyna (pre-pro-OXY-NP I). Ostatecznym produktem potranslacyjnej
modyfikacji jest aktywny hormon — oksytocyna (OXY) — i nieaktywny
polipeptyd — neurofizyna oksytocynowa (NP I).
Enzymatyczna modyfikacja pre-pro-hormonu zaczyna się wewnątrz
siateczki śródplazmatycznej, jest kontynuowana w aparacie Golgiego
i kończy się w pęcherzykach odczepiających się od aparatu Golgiego,
zawierających oksytocynę zbudowaną z 9 aminokwasów i neurofizynę
oksytocynowa o znacznie dłuższym łańcuchu polipeptydowym składającym się z 94 aminokwasów.
40
Prawa fizjologiczne organizmu człowieka
94
Ryc. 13. Gen oksytocyny i powstawanie w wyniku: (1) transkrypcji — pre-matrycowego
RNA, (2) modyfikacji potranskrypcyjnej — matrycowego RNA, (3) translacji — pre-pro-oksytocyno-neurofizyny I i (4) modyfikacji potranslacyjnej — oksytocyny i neurofizyny I.
W łańcuchach DNA genu oksytocyny występują: promotor (nukleotydy 1—260), trzy egzony
e. A, e. B i e. C (nukleotydy 261—381, 595—822, 887—942), dwa introny i. 1 i i. 2
(nukleotydy 382—594, 823—886) i część końcowa (nukleotydy 943—1055).
Synteza białka, a więc procesy translacji i potranslacyjnej modyfikacji, zachodzą przy udziale odpowiednich enzymów wewnątrzkomórkowych. Aktywność tych enzymów zależy od metabolizmu wewnątrzkomórkowego uwarunkowanego napływem do komórek składników
energetycznych i budulcowych oraz cząsteczek tlenu. Również informacja
docierająca do komórek w postaci cząsteczek hormonów i innych przekaźników chemicznych oddziałuje na syntezę białka po:
• związaniu się z receptorami w błonie komórkowej i aktywacji lub
inaktywacji „drugiego przekaźnika" przenoszącego informację wewnątrz komórek;
Czynność komórki
41
• związaniu się z receptorami cytoplazmatycznymi, jak to ma miejsce
w przypadku steroidów;
• związaniu się z receptorem jądrowym, co odnosi się do hormonu
gruczołu tarczowego — trijodotyroniny (T 3).
Cząsteczki steroidów przenikają przez warstwę fosfolipidową błony
komórkowej i dostają się do cytoplazmy, gdzie łączą się z receptorem
cytoplazmatycznym. Taki kompleks receptor połączony z cząsteczką
steroidu wędruje do jądra komórkowego, modulując proces transkrypcji,
czyli tworzenia się pre-mRNA dla określonych białek. W przeciwieństwie do steroidów, trijodotyronina (T 3 ) wnika bezpośrednio
do jądra komórkowego, łączy się tam z odpowiednim receptorem jądrowym i na tej drodze oddziałuje na procesy związane z tran skrypcją.
Ryc. 14. Oddziaływanie steroidów na aparal genetyczny jądra komórkowego. Steroidy
po dostaniu się do cytoplazmy wiążą się z receptorem cytoplazmatycznym i w postaci
kompleksu steroid-receptor wnikają do jądra komórkowego, modyfikując proces transkrypcji.
42
Prawa fizjologiczne organizmu człowieka
I Siateczka śródplazmatyczna gładka
W obrębie siateczki śródplazmatycznej gładkiej (reticulum endo
plasmaticum nongranulosum) zachodzi biosynteza i magazynowanie nie
których związków, przede wszystkim tłuszczów, oraz polimeryzacja gluko
zy i tworzenie się ziarnistości glikogenu. W komórkach kory nadnerczy
jajników i jąder odbywa się w obrębie błon gładkich biosynteza hormonów
steroidowych, w komórkach wątroby odkładają się ziarna glikogenu
w komórkach mięśniowych poprzecznie prążkowanych zaś gromadzą si^
jony wapniowe.
I Lizosomy
Lizosomy {lysosoma) są to pęcherzyki o średnicy około 400 nm otoczone pojedynczą błoną i zawierające enzymy hydrolityczne trawiące
białka, kwasy nukleinowe i węglowodany. W obrębie lizosomów zachodzi
trawienie zarówno fragmentów własnej komórki, jak i fragmentów obcych
komórek i obcych cząsteczek, które zostały wchłonięte do wnętrza na
drodze fagocytozy lub pinocytozy. Niedostateczna prężność cząsteczek
tlenu (Po2) w komórce, czyli niedotlenienie, powoduje przechodzenie
enzymów przez błonę lizosomalną do cytoplazmy i samostrawienie
komórki.
Endocytoza
Endocytoza obejmuje dwa procesy: fagocytozę i pinocytozę. Fagocytoza zachodzi wtedy, kiedy większe fragmenty obcych komórek lub
mikroorganizmy zostają otoczone błoną komórkową i są wciągane
do wnętrza komórki, gdzie tworzą wakuole. Z kolei do tych wakuoli
otwierają się lizosomy zawierające enzymy. Dzięki nim sfagocytowane
fragmenty ulegają strawieniu w obrębie wakuoli i zostają uwolnione do
cytoplazmy, w której mogą pozostać w postaci ciał resztkowych.
Pinocytoza jest procesem podobnym do fagocytozy z tą różnicą, że
dotyczy cząsteczek związków chemicznych, które przyczepiają się do
zewnętrznej powierzchni błony komórkowej. W tym miejscu błona ulega
zagłębieniu aż do wytworzenia się wakuoli. Następnie cząsteczki te są
trawione przez enzymy zawarte w lizosomach, a produkty końcowe
Czynność komórki
43
Egzocytoza
Fagocyłoza
Pinocytoza
Ryc. 15. Endocytoza — fagocytoza i pinocytoza oraz egzocytoza. Enzymy lizosomalne
trawią wchłonięte fragmenty komórek lub płynne składniki. Egzocytoza — zsyntetyzowane
w siateczce śródplazmatycznej związki są przejściowo magazynowane w pęcherzykach
i wydalane na zewnątrz komórki.
44
Prawa fizjologiczne organizmu człowieka
hydrolizy przechodzą do cytoplazmy. Dzięki pinocytozie do wnętrza
komórki dostają się duże cząsteczki, np. białka, które nie przenikają
przez błonę komórkową.
Egzocytoza
Substancje wytworzone przez komórki gruczołów wydzielania zewnętrznego i wewnętrznego, przez komórki nerwowe i inne komórki
w organizmie są przejściowo magazynowane w pęcherzykach otoczonych
błoną. W komórkach gruczołowych pęcherzyki te nazywane są również
ziarnami wydzielniczymi, w komórkach nerwowych zaś — pęcherzykami
synaptycznymi. W komórkach nerwowych występują małe pęcherzyki
o średnicy od 20 do 120 nm, wypełnione związkami o bardzo dużej
aktywności fizjologicznej — transmitterami i modulatorami. W komórkach gruczołowych, w zależności od ich rodzaju, znajdują się pęcherzyki
o średnicy od kilkudziesięciu do kilkuset nanometrów. W pęcherzykach
tych zgromadzone są składniki wydzielane do krwi (hormony, białka
osocza) lub wydzielane na zewnątrz (enzymy w sokach trawiennych).
W czasie pobudzenia komórki dochodzi do zetknięcia się pęcherzyków
z powierzchnią wewnętrzną błony komórkowej. W tym miejscu następuje
jednoczesne pęknięcie błony komórkowej i błony pęcherzyków. Treść
wypełniająca pęcherzyki zostaje usunięta na zewnątrz. Puste pęcherzyki
zamykają się i są wciągane do wnętrza komórki, gdzie mogą ponownie
napełniać się.
I Mitochondria
Mitochondria wyposażone są w podwójną błonę mitochondrialną—zewnętrzną i wewnętrzną. Błona wewnętrzna jest silnie pofałdowana
i tworzy grzebienie mitochondrialne. Błony otaczają przestrzeń zamkniętą
wypełnioną macierzą (matrix). W obrębie błon mitochondrialnych znajdują się wszystkie enzymy cyklu kwasów trikarboksylowych, czyli cyklu
kwasu cytrynowego (tricarboxylic acid cycle, citric acid cycle), enzymy
łańcucha oddechowego (respiratory chain) oraz enzym monoaminooksydaza — MAO (monoamine oxidase). W macierzy występuje pewna ilość
kwasu deoksyrybonukleinowego (DNA) oraz jony Ca 2+ i Mg2 + .
Mitochondria są strukturami komórki, w których głównie wytwarzana
jest energia. W obrębie mitochondriów są syntetyzowane adenozynotrifosforany — ATP (adenosine triphosphate), które są uniwersalnymi
przenośnikami energii w komórce.
Metabolizm wewnątrzkomórkowy
45
Ryc. 16. Budowa mitochondrium: a — ogólna; b — grzebienia mitochondrialnego. 1 — błona zewnętrzna, 2 — błona
wewnętrzna, 3 — grzebień, 4 — macierz, 5 — ziarenko mitochondrialne, 6 — cząsteczka uszypułowana. (Reprodukowano za zezwoleniem Michała Karaska.)
I Metabolizm wewnątrzkomórkowy
Komórki pobierają z otoczenia, czyli z płynu międzykomórkowego,
składniki odżywcze. Składnikami tymi dla komórek są: glukoza, aminokwasy i kwasy tłuszczowe. Pobieranie składników odżywczych odbywa się
dzięki aktywnemu transportowi cząsteczek przez błonę komórkową
przeciw gradientowi stężeń, w wyniku pinocytozy i w niektórych rodzajach komórek również dzięki fagocytozie.
Wyzwolenie energii ze składników odżywczych odbywa się w procesie
odłączenia atomów wodoru od produktów rozpadu składników odżywczych i przenoszenia tych atomów na atomy tlenu. Na drodze dyfuzji
poprzez błonę komórkową stale są dostarczane atomy tlenu z zewnątrz,
czyli z płynu międzykomórkowego, do wnętrza komórki. Prężność cząsteczek tlenu (Po2) w każdej komórce jest wypadkową szybkości, z jaką
są one dostarczane, oraz szybkości, z jaką są one zużywane wewnątrz
komórki. Przy jednakowej szybkości dostawy mała prężność tlenu występuje w komórkach wytwarzających dużo energii, a więc intensywnie
pracujących.
Wytwarzanie energii w komórce jest istotą oddychania wewnętrznego
(internal respiration), czyli oddychania wewnątrzkomórkowego. Proces
oddychania wewnątrzkomórkowego przebiega w dwóch fazach. W pierwszej fazie — beztlenowej — i w drugiej — tlenowej. W fazie beztlenowej
energia pozyskiwana jest w procesie glikolizy z głównego składnika
odżywczego jakim jest glukoza. Przekształceniu glukozy w pirogronian
towarzyszy powstanie dwóch cząsteczek ATP. Nie wymaga to obecności
46
Prawa fizjologiczne organizmu człowieka
Ryc. 17. Wytwarzanie energii w komórce potrzebnej do syntezy ATP w następstwie rozpadu glikogenu komórkowego w warunkach: a — beztlenowych; b — tlenowych.
tlenu. W fazie tlenowej tlen jest niezbędny do dalszego pozyskiwania
energii z pirogronianu w wyniku jego rozkładu do dwutlenku węgla i wody.
W razie braku tlenu w komórce, zachodzi tylko glikoliza beztlenowa
kończąca się przekształceniem pirogronianu w mleczan.
Proces glikolizy i powstawanie pirogronianu przebiega w cytoplazmie
komórkowej. Natomiast w mitochondriach następuje włączanie do cyklu
kwasu cytrynowego zarówno pirogronianu, który jest wcześniejszym
ogniwem glikolizy, jak i innych metabolitów cyklu kwasów trikarboksylowych wytworzonych w procesie katabolizmu aminokwasów i kwasów
tłuszczowych.
Enzymy cyklu kwasu cytrynowego występują w macierzy mitochondrialnej, białka łańcucha oddechowego zaś znajdują się w wewnętrznej
błonie mitochondrialnej. W czasie przemian w cyklu kwasu cytrynowego
uwalniane są atomy wodoru oraz cząsteczki dwutlenku węgla. Atomy
wodoru są transportowane przez białka łańcucha oddechowego aż do
momentu połączenia ich z tlenem i powstania cząsteczki wody.
Energia wyzwolona w czasie rozpadu glukozy do pirogronianu zużyta
jest do syntezy ATP. Przeciętnie w fazie beztlenowej zostaje zsyntetyzowane około 5% ogólnej ilości ATP komórkowego. Pozostałe 95% ATP
komórkowego zostaje syntetyzowane w mitochondriach w fazie tlenowej.
Metabolizm wewnątrzkomórkowy
47
Adenozynotrifosforan (ATP) syntetyzowany jest z adenozynodifosforanu (ADP — adenosine diphosphate) i ortofosforanu. Adenozynotrifosforan jest bogaty w energię wykorzystywaną w procesach wewnątrzkomórkowych.
Energia wyzwolona w czasie rozpadu ATP zostaje zużyta:
• na transport aktywny jonów i substancji drobnocząsteczkowych przez
błonę komórkową wbrew gradientowi st ężenia oraz na transport
wewnątrzkomórkowy;
• do syntezy składników komórkowych, takich jak DNA, RNA, białka,
lipidy, oraz do resyntezy glukozy i polimeryzacji glikogenu komórko
wego;
• na pracę mechaniczną komórki polegającą na ruchu cytoplazmy komór
kowej lub skracaniu się białek kurczliwych w komórkach mięśniowych.
Uwolniony w czasie jednego z powyższych procesów ADP powraca
do mitochondriów, gdzie ponownie uczestniczy w resyntezie ATP.
W ten sposób ATP służy jako uniwersalny magazyn i jednocześnie
przenośnik energii w komórkach.
48
Prawa fizjologiczne organizmu człowieka
Samoregulacja metabolizmu
komórkowego
I
Procesy biosyntezy komórkowej podlegają kontroli humoralnej lub
kontroli nerwowej ze strony innych komórek znajdujących się często
w odległych narządach. Na równi z kontrolą zewnętrzną, humoralną
lub nerwową procesy biosyntezy podlegają samoregulacji w obrębie
pojedynczej komórki. Nadmiar jakiegoś produktu wytworzonego w komórce hamuje jego dalszą syntezę zarówno na etapach końcowych przez
inaktywowanie odpowiedniego enzymu, jak również na etapach pośrednich i początkowych dzięki hamowaniu syntezy tego enzymu. W ten
sposób komórka zabezpiecza się przed nagromadzeniem się w niej
pewnych związków w ilościach przekraczających granice fizjologiczne.
Przekroczenie tej granicy przy niedostatecznie sprawnej samoregulacji
może doprowadzić do obumierania komórki.
I
Cykl komórkowy i śmierć
komórki
Komórki nabłonka, krwiotwórcze i tkanki łącznej zachowują
zdolność do podziałów na komórki potomne w ciągu całego życia
organizmu człowieka. Nie dzielą się natomiast komórki nerwowe i
mięśni poprzecznie prążkowanych po zakończeniu wzrostu organizmu.
Komórka zdolna do podziałów pozostaje w spoczynku, czyli w fazie Go.
Komórki somatyczne z tej fazy spoczynkowej wstępują w fazę Gx i
rozpoczynają cykl komórkowy. Następną fazą jest faza S, w czasie
której zachodzi w jądrze komórkowym replikacja łańcuchów DNA, jak
również wzmożona synteza białka jądrowego, histonowego i
niehistonowego. Następna jest faza G2 , która tym się charakteryzuje, że
w jądrach komórek somatycznych występuje podwójna liczba
chromosomów i łańcuchów DNA. W fazie mitozy, następującej po fazie
G2, chromosomy rozdzielają się do dwóch komórek potomnych, po czym
komórki potomne mogą rozpocząć następny cykl komórkowy lub
pozostają w spoczynku w fazie Go (patrz ryc. 10, str. 34). Zarówno
komórki dzielące się, jak i nie dzielące się są wrażliwe na zmiany
zachodzące w środowisku wewnętrznym organizmu, szczególnie na
niedobór tlenu niezbędnego do metabolizmu wewnątrzkomórkowego.
Niedobór tlenu powoduje śmierć komórek, czyli ich nekrozę (necrosis),
przejawiającą się w samostrawieniu przez własne
Kontrola i regulacja funkcji
49
enzymy wewnątrzkomórkowe, rozpadzie błony komórkowej i wypłynięciu
składników komórkowych.
W przeciwieństwie do czynników fizycznych i chemicznych, działających na komórki z zewnątrz i powodujących ich śmierć, w warunkach
fizjologicznch w organizmie występuje również zaprogramowana śmierć
komórek, czyli apoptoza {apoptosis). Dochodzi do ekspresji określonej
grupy genów i zwiększenia aktywności innych enzymów komórkowych.
Komórki podlegające apoptozie są szybko usuwane przez komórki
żerne wędrujące w tkankach i pożerające, czyli fagocytujące, martwe
komórki.
I Kontrola i regulacja funkcji I
Specjalizacja funkcji w organizmie
W procesie ewolucji w organizmach zwierzęcych rozwinęły się wyspecjalizowane narządy i układy, dzięki którym mogą one coraz lepiej
przystosować się do zmieniającego się środowiska zewnętrznego. Mimo
znacznych i długotrwałych zmian otoczenia procesy życiowe organizmu
nie mogą przekraczać granic fizjologicznych.
Organizm człowieka, dzięki rozwojowi ośrodkowego układu nerwowego, jest najlepiej przystosowany do zmian zachodzących w środowisku
zewnętrznym. Z drugiej jednak strony społeczeństwo, w jakim żyje każdy
człowiek, czyli środowisko społeczne (social environment), i wytwory
cywilizacji stanowią najbardziej złożone środowisko zewnętrzne.
Powstałe w procesie ewolucji wyspecjalizowane układy przejęły na
siebie funkcję pośredniczenia pomiędzy środowiskiem zewnętrznym i wewnętrznym organizmu i optymalnego adaptowania się do otoczenia (adaptation to environment). Funkcje życiowe organizmu związane są z utrzymaniem jego życia osobniczego, a więc z metabolizmem oraz z utrzymaniem gatunku, czyli z rozrodem.
Metabolizm wymaga:
• odżywiania (nutrition), a więc pobierania ze środowiska zewnętrznego
materiałów energetycznych i budulcowych;
• oddychania (respiration), czyli doprowadzania ze środowiska zewnętrz
nego tlenu niezbędnego do procesów utleniania wewnątrzkomórkowe
go i usuwania nadmiaru dwutlenku węgla ze środowiska wewnętrznego;
50
Prawa fizjologiczne organizmu człowieka
Ryc. 19. Układy w organizmie człowieka: oddechowy, trawienny, sercowo-naczyniowy,
moczowo-płciowy, nerwowy (ośrodkowy i obwodowy), ruchowy, gruczoły dokrewne (G)
i receptory.
• krążenia (circulation) materiałów energetycznych, budulcowych, ga
zów (O2 i CO2), produktów przemiany materii i innych ciał pomiędzy
wyspecjalizowanymi komórkami stykającymi się bezpośrednio ze śro
dowiskiem zewnętrznym i nie stykającymi się z nim;
• wydalania (excretion), czyli usuwania ze środowiska wewnętrznego
wytworzonych nielotnych produktów przemiany materii.
Konieczność zapewnienia stałego metabolizmu i stałego środowiska
wewnętrznego organizmu doprowadziła w procesie ewolucji do wykształcenia się układu ruchowego (somatycznego), który zapewnił lepsze pobieranie materiałów energetycznych i budulcowych ze środowiska zewnętrznego oraz umożliwił jego zmianę. Układ ruchowy obejmuje przede
wszystkim układ mięśni szkieletowych, którego komórki kurcząc się
powodują ruchy ciała, układ szkieletowy stanowiący rusztowanie dla
mięśni szkieletowych, narządy odbierające bodźce, czyli receptory, oraz
Kontrola i regulacja funkcji
51
układ nerwowy somatyczny przekazujący pobudzenie od narządów odbierających (receptorów) do układu mięśniowego.
Utrzymanie stałego środowiska wewnętrznego wymaga stałej precyzyjnej kontroli czynności układów związanych z odżywianiem, oddychaniem, krążeniem i wydalaniem. Taka kontrola środowiska wewnętrznego przypada układowi nerwowemu. Część obwodowego układu nerwowego, jaką stanowi układ nerwowy autonomiczny, przekazuje impulsację bezpośrednio do tkanek i narządów wewnętrznych.
I Przetwarzanie i przenoszenie informacji
Zarówno w środowisku zewnętrznym (external environment), jak
i w środowisku wewnętrznym (internal environment) zachodzą stałe
zmiany. Nieznaczna nawet zmiana w środowisku jest źródłem sygnałów
odbieranych przez receptory. Każdy sygnał niesie informację ze środowiska zewnętrznego, np. fale świetlne, fale akustyczne itp., lub ze środowiska wewnętrznego, np. prężność tlenu we krwi, koncentracja jonów
wodorowych itp. W obrębie receptorów zachodzi przetwarzanie informacji (transduction of information). W organizmie informacja jest stale
przetwarzana w sposób analogowy i cyfrowy, jak również jest stale przenoszona. W procesie tym ilość informacji nie zmienia się.
Przetwarzanie, a następnie przenoszenie informacji (transmission of
information) w sposób ciągły, czyli analogowy, ma miejsce np. za pośrednictwem cząsteczek krążących we krwi hormonów. Proporcjonalnie do wzrostu koncentracji cząsteczek hormonu w określonym cza sie wzrasta ilość przenoszonej informacji w tym samym czasie. Przenoszenie analogowe informacji zachodzi zazwyczaj na drodze humoralnej. W jednym narządzie stanowiącym generator sygnałów, np.
gruczole dokrewnym, wytwarzane są substancje chemiczne, które za
pośrednictwem krwi i układu sercowo-naczyniowego przenoszone są
do innych narządów, na które wywierają swój wpływ. Na przykład
przez komórki dokrewne błony śluzowej dwunastnicy wydzielany jest
Ryc. 20. Odbiór informacji na wejściu
układu, przetwarzanie i generowanie informacji na wyjściu.
52
Prawa fizjologiczne organizmu człowieka
polipeptyd, zwany sekretyną, który krążąc we krwi pobudza trzustkę do
wydzielania soku trzustkowego. Ilość wydzielonego soku trzustkowego
w jednostce czasu jest proporcjonalna do koncentracji krążących we krwi
cząsteczek sekretyny.
Przetwarzanie, a następnie przenoszenie informacji w sposób przerywany, impulsowy (czyli cyfrowy, tak jak w komputerach), zachodzi np.
we włóknach nerwowych. Narządy receptorowe spełniają funkcję przetworników (transducers) zamieniających informację ze środowiska zewnętrznego lub wewnętrznego organizmu na salwy impulsów nerwowych
przewodzonych przez włókna nerwowe. Poszczególne impulsy nerwowe
przewodzone przez to samo włókno nerwowe, czyli przez jeden kanał
Kontrola i regulacja funkcji
53
informacyjny (information channel), nie różnią się między sobą. Informacja zakodowana jest w częstotliwości, z jaką impulsy nerwowe przewodzone są przez włókno nerwowe, czyli w długości interwałów pomiędzy
pojedynczymi impulsami.
Informacja przetwarzana jest w organizmie nie tylko przez receptory,
lecz również przez komórki nerwowe. Odebraną informację cyfrową
komórki nerwowe zamieniają na informację analogową, w postaci zmian
potencjału elektrycznego błony komórkowej. Ta sama komórka wysyła
następnie informację cyfrową do innych komórek nerwowych.
Narząd /
kontrolujący
Narząd U.
kontrolowany
Ryc. 22. Kontrola jednokierunkowa. Narząd I kontrolujący jest generatorem informacji
odbieranej przez narząd II kontrolowany.
Teoria informacji, która jest przydatna w fizjologii do analizowania
mechanizmów zapewniających prawidłowy przebieg procesów życiowych,
posługuje się jednostką informacji, którą jest bit (binary digit) oraz takimi
pojęciami, jak układy i systemy. System jest to zbiór elementów połączonych ze sobą zgodnie z określonym wzorem i spełniających określone czynności. Prosty system, którym może być nawet pojedyncza
komórka, jest nazywany układem. Połączenia pomiędzy elementami
systemów lub układów za pośrednictwem kanałów informacyjnych przedstawia się w postaci schematów blokowych. Poszczególne bloki czynnościowe mają swoje wyjścia i wejścia oraz łączą się kanałami, przez które
przenoszona jest informacja.
I Komunikacja pomiędzy komórkami
W organizmie człowieka prawidłowy przebieg różnorodnych funkcji
zależy od odpowiedniej komunikacji pomiędzy komórkami zarówno
sąsiadującymi ze sobą, jak również niesąsiadującymi. Komórki sąsiadu-
54
Prawa fizjologiczne organizmu człowieka
jące ze sobą przekazują informacje bezpośrednio z komórki do komórki.
z pominięciem płynu zewnątrzkomórkowego, dzięki plamkom przylega-i
nia i połączeniom szczelinowym (gap junction). W miejscu przylegania
do siebie dwóch komórek w ich błonach komórkowych występują
koneksony tworzące wspólny kanał, przez który przepływają z jednej
komórki do drugiej jony, cukry proste, aminokwasy i przekaźniki we-l
wnątrzkomórkowej informacji. Tak komunikują się ze sobą komórki
mięśniowe poprzecznie prążkowane mięśnia sercowego (patrz str. 110).
Ryc. 23. Połączenia pomiędzy cytoplazma dwóch komórek za pośrednictwem koneksonu
tworzącego wspólny kanał. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II
wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Drugi typ komunikacji związany jest z przenoszeniem informacji
w postaci substancji chemicznych przez płyn zewnątrzkomórkowy i płyny
ustrojowe, czyli na drodze humoralnej (humoral communication). Komórka wydziela substancje chemiczne, które wiążą się z receptorami błony
komórkowej tej samej komórki. Jest to komunikacja autokrynna (autocrine
communication). Działanie substancji chemicznych poprzez płyn zewnątrzkomórkowy na sąsiednie komórki stanowi komunikację parakrynną
(paracrine communication). Komunikacja endokrynna (endocrine communication) polega na wydzielaniu substancji chemicznych przez komórki
jednego narządu do krążących płynów ustrojowych, szczególnie do krwi,
i oddziaływaniu tą drogą na metabolizm komórkowy innych odległych
narządów (patrz str. 225).
Kontrola i regulacja funkcji
55
Ryc. 24. Przenoszenie informacji pomiędzy komórkami za pośrednictwem substancji
chemicznych uwolnionych do płynu zewnątrzkomórkowego: a — komunikacja autokrynna;
b — komunikacja parakrynna, c — komunikacja endokrynna. ECF — płyn zewnątrzkomórkowy, ICF — płyn wewnątrzkomórkowy.
Trzeci typ komunikacji odbywa się za pośrednictwem komórek nerwowych (neural communication). W błonie komórkowej ich wypustek
otwierają się kanały dla prądów jonowych, co powoduje przemieszczanie
się potencjału elektrycznego wzdłuż wypustek, czyli przewodzenia impulsów nerwowych. Przekazywanie informacji przez komórki nerwowe do
innych komórek nerwowych lub do komórek unerwianych narządów
zachodzi za pośrednictwem synaps. Znaczna ich większość są to synapsy
chemiczne, w których dochodzi do wydzielania przekaźnika chemicznego
Ryc. 25. Schemat synaps chemicznych (1, 2) przenoszących informację za pośrednictwem
przekaźników chemicznych i synapsy elektrycznej (3). (Reprodukowano za zezwoleniem
z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
56
Prawa fizjologiczne organizmu człowieka
do przestrzeni synaptycznej i następnie do jego związania się z recepto-1
rami w błonie komórkowej komórki odbierającej. W ośrodkowym ukła-l
dzie nerwowym występują również w znacznie mniejszej liczbie synapsy
elektryczne. Jeżeli przestrzeń synaptyczna jest wąska, to ruch jonów przez
błonę komórkową neuronu wysyłającego impulsy nerwowe powoduje
w komórce odbierającej otwieranie się kanałów bramkowanych napięciem
dla dokomórkowego prądu jonów o dodatnim ładunku elektrycznym.
W ten sposób odbierane są impulsy nerwowe i przenoszona jest informacja bez pośrednictwa synaptycznego przekaźnika chemicznego.
I Sprzężenie zwrotne
W organizmie czynność jednych układów (narządów) stale jest sterowana przez inne układy. Informacja przepływa od wyjścia układu sterującego do wejścia układu sterowanego, wymuszając na jego wyjściu określony proces. Przepływ informacji jest jednokierunkowy, tym samym
kontrola czynności jest jednokierunkowa o typie sprzężenia prostego.
Narzqd I
regulujący
Narząd U
regulowany
Ryc. 26. Kontrola wzajemnie zwrotna. Narząd I regulujący jest generatorem informacji
odbieranej przez narząd II regulowany. Informacja wysyłana przez narząd regulowany
trafia do układu komparatora (komp.) narządu regulującego dzięki sprzężeniu zwrotnemu.
Większość procesów fizjologicznych w organizmie podlega regulacji
dzięki sprzężeniom zwrotnym. Ujemne sprzężenie zwrotne (negative feedback) pomiędzy układem sterującym i sterowanym zapewnia stabilizację
funkcji fizjologicznych.
Kontrola i regulacja funkcji
57
Kontrola jednokierunkowa — zarówno nerwowa, jak i humoralna —
polega na przekazywaniu informacji tylko w jednym kierunku, z jednego
narządu do drugiego. Kontrola wzajemnie zwrotna, czyli regulacja, związana jest z dwukierunkowym przesyłaniem informacji zakodowanych zarówno w postaci impulsów nerwowych, jak i substancji chemicznych.
Wzmożona czynność jednego narządu jest źródłem informacji pobudzającej drugi narząd, który z kolei wysyła informacje hamujące czynność
pierwszego narządu.
Kontrola wzajemnie zwrotna jest wyższą formą kontroli i przede
wszystkim dzięki niej utrzymuje się stałe środowisko wewnętrzne organizmów stojących na wysokich szczeblach rozwoju filogenetycznego.
Informacja wysyłana przez układ sterowany przez kanał sprzężenia
zwrotnego, zwany również pętlą sprzężenia zwrotnego (feedback loop),
trafia do komparatora w układzie sterującym. W komparatorze zachodzi
porównanie informacji wysyłanej przez układ sterujący z informacją
odbieraną od układu sterowanego. W wyniku tego procesu dochodzi do
odpowiedniej zmiany informacji wysyłanej.
Przedstawienie zależności występujących pomiędzy poszczególnymi
narządami w postaci sprzężeń prostych i sprzężeń zwrotnych pomiędzy
układami blokowymi stanowi znaczne uproszczenie w porównaniu z rzeczywistymi zależnościami w żywym organizmie. Uwzględniane są tylko
wybrane zależności, zazwyczaj najbardziej istotne, a pomijane bardzo
liczne drugorzędne oddziaływania.
W komunikacji pomiędzy komórkami istotną rolę spełniają receptory
błony komórkowej. Zwiększenie stężenia przekaźnika chemicznego w środowisku otaczającym komórkę prowadzi do internalizacji receptorów,
ICF
Ryc. 27. Gęstość receptorów błonowych na zewnętrznej powierzchni błony komórkowej
zależna od stężenia przekaźnika chemicznego: a — duże stężenie przekaźnika chemicznego — „regulacja w dół"; b — pośrednie stężenie przekaźnika chemicznego; c — małe
stężenie przekaźnika chemicznego — „regulacja w górę".
58
Prawa fizjologiczne organizmu człowieka
czyli do zmniejszenia ich gęstości na zewnętrznej powierzchni błony
komórkowej. Proces ten nazywany jest „regulacją w dół" (down regułation). Przeciwnie — zmniejszenie stężenia przekaźnika chemicznego w środowisku otaczającym komórkę wywołuje eksternalizację receptorów błonowych i zwiększenie ich gęstości na powierzchni zewnętrznej błony
komórkowej. Zwiększenie gęstości receptorów błonowych jest to tak
zwana „regulacja w górę" (up regulation).
Regulacja gęstości specyficznych receptorów na błonie komórkowej
jest istotnym fizjologicznym mechanizmem występującym w ujemnych
sprzężeniach zwrotnych. Dzięki tym mechanizmom zapewniona jest
stabilizacja funkcji komórek, tkanek i narządów w granicach fizjologicznych.
Czynność komórek nerwowych i
mięśniowych
I Pobudliwość i pobudzenie
Pobudliwość i pobudzenie są to cechy żywych komórek. Pobudzenie
(excitation) jest to zmiana właściwości błony komórkowej lub metabolizmu komórkowego pod wpływem czynników działających z zewnątrz
komórki, czyli pod wpływem bodźców. W warunkach fizjologicznych
bodźcami działającymi na przeważającą liczbę komórek w organizmie
są substancje chemiczne. Komórki natomiast tworzące w organizmie
narządy odbiorcze, czyli receptory, odbierają w warunkach fizjologicznych
również bodźce fizyczne w różnej postaci, np. fal świetlnych, fal akustycznych, energii cieplnej, energii mechanicznej (jako ucisk lub rozciąganie itp.). Każdy bodziec fizyczny lub chemiczny, działając w dostatecznie
dużym natężeniu na komórki, może wywołać ich pobudzenie. Bodźce
fizjologiczne są to takie bodźce, które nie uszkadzają komórki i wywołują
całkowicie odwracalne procesy. Niektóre komórki w organizmie człowieka odbierają w czasie swojego życia kilka miliardów bodźców, zmieniając za każdym razem swój metabolizm.
Pobudliwość (excitability) jest to zdolność reagowania na bodziec.
Substancje chemiczne występujące w płynie zewnątrzkomórkowym, wiążąc się z receptorami w błonie komórkowej, otwierają kanały dla prądów
jonowych lub aktywują enzymy w niej zawarte. Komórki jednych tkanek
reagują bardzo szybko na bodziec, w ułamkach sekundy, otwieraniem się
kanałów jonowych, natomiast zmiana metabolizmu w innych tkankach
wymaga dłuższego czasu. Z tego względu do pobudliwych zalicza się te
tkanki, które szybko odpowiadają na bodźce. Są to tkanki zbudowane
z komórek nerwowych i ich wypustek oraz z komórek mięśniowych:
mięśni poprzecznie prążkowanych, mięśni gładkich i mięśnia sercowego.
60
I
Czynność komórek nerwowych i mięśniowych
Potencjał spoczynkowy błony
komórkowej
Pomiędzy wnętrzem komórek tkanek pobudliwych a płynem zewnątrz-1
komórkowym występuje stale w spoczynku ujemny potencjał elektryczny,!
czyli potencjał spoczynkowy błony komórkowej (resting membranę poten-1
tial). Ujemny potencjał spoczynkowy wewnątrz neuronu i jego wypustek I
wynosi od —60 do —80 mV, średnio —70 mV, w komórkach mięśnio-l
wych poprzecznie prążkowanych zaś od — 80 do — 90 mV.
Stężenie poszczególnych jonów w płynie wewnątrzkomórkowym znacz-1
nie się różni od ich stężenia w płynie zewnątrzkomórkowym (patrz tab. 1,1
str. 21). Wewnątrz komórek występują w znacznym stężeniu aniony I
organiczne nie przechodzące przez błonę komórkową. Nieznaczna przewa-1
ga ładunków ujemnych wszystkich anionów w stosunku do kationów I
w płynie wewnątrzkomórkowym jest przyczyną występowania ujemnego
potencjału spoczynkowego wewnątrz komórek. Błona komórkowa jest
spolaryzowana, po stronie wewnętrznej skupione są jony o ładunku
ujemnym, po stronie zewnętrznej zaś jony o ładunku dodatnim.
Stężenia poszczególnych jonów w płynie wewnątrzkomórkowym nie
zmieniają się, jeśli metabolizm nie ulega zmianie i jeśli na błonę komórkową nie działają bodźce z zewnątrz. W tych warunkach wytwarza się
równowaga pomiędzy stężeniem poszczególnych jonów na zewnątrz
i wewnątrz komórek. Równowaga ta jest wypadkową gradientów koncentracji i gradientów ładunków elektrycznych poszczególnych jonów płynu
zewnątrz- i wewnątrzkomórkowego.
Na podstawie równania Nernsta i znanej koncentracji poszczególnych
jonów w płynie zewnątrz- i wewnątrzkomórkowym został obliczony
potencjał elektryczny panujący wewnątrz komórek mięśni poprzecznie
prążkowanych. Jest to potencjał ujemny równy około —90 mV.
Równanie Nernsta w zastosowaniu do K+:
Pobudliwość i pobudzenie
61
Obliczony teoretycznie z równania Nernsta ujemny potencjał dla
komórek mięśni poprzecznie prążkowanych został potwierdzony za pomocą bezpośredniego pomiaru mikroelektrodami wewnątrzkomórkowymi. Pomiar ten jednocześnie wykazał, że dyfuzja jonów K+ do wnętrza
komórki przeciw istniejącemu gradientowi stężeń wymaga większej różnicy potencjałów niż —90 mV. Tym samym dyfuzja ta nie jest procesem
biernym, lecz czynnym, wymagającym zużycia energii.
Występowanie prądów jonowych w błonie komórkowej skierowanych
do wnętrza lub na zewnątrz komórki uzasadnia przyjęcie hipotezy
o istnieniu oddzielnych kanałów (ionic channels) w błonie komórkowej
dla poszczególnych jonów. W zależności od stanu czynnościowego kanały
dla prądów poszczególnych jonów otwierają się lub zamykają. W procesie tym biorą udział białka tworzące kanały jonowe, stanowiące integralną część błony komórkowej.
Pompa sod owo-potasowa
Utrzymanie wewnątrz komórek dużego stężenia jonów K + i małego
stężenia jonów Na + wymaga aktywnego transportu obu tych kationów
przez błonę komórkową przeciwko gradientowi stężeń. Kationy sodowe
napływające do wnętrza komórki przez kanały dla prądów jonów sodowych
zostają po stronie wewnętrznej błony komórkowej związane z enzymem.
Znajduje się on w samej błonie komórkowej i transportuje jony Na + na
zewnątrz błony. Jednocześnie ten sam enzym zabiera ze sobą jony K +
z zewnętrznej powierzchni błony komórkowej i transportuje je do wnętrza
komórki.
Enzym transportujący jony Na+ i K+ przez błonę komórkową przeciw
gradientowi stężeń czerpie energię z hydrolizy ATP do ADP. Enzym ten
stanowi cząsteczkę o masie około 294 kDa składającą się z dwóch
podjednostek alpha (2 x 92 kDa m.cz.) i beta (2 x 55 kDa m.cz.). Aktywowany jest przez jony Na+ i K+. Stąd pochodzi jego nazwa adenozynotrifosfataza aktywowana przez sód i potas: Na-K-ATP-aza (sodium-potassium-activated adenosine triphosphatase).
Energia wyzwolona z rozpadu 1 mol ATP do ADP wykorzystana jest
na antyport 3 mol Na + z komórki i 2 mol K+ do komórki. Rozpad ATP
do ADP pod wpływem Na-K-ATP-azy zachodzi w obecności jonów
magnezowych zawartych w płynie wewnątrzkomórkowym.
Napęd pompy sodowo-potasowej jest związany z metabolizmem wewnątrzkomórkowym. Około 30% całego metabolizmu komórkowego
tkanek pobudliwych pozostających w spoczynku jest na niego zużywane.
62
Czynność komórek nerwowych i mięśniowych
Optymalna praca pompy sodowo-potasowej i związana z tym optymalna pobudliwość wymaga:
• stałego dopływu do komórek tlenu i substancji energetycznych (gluko
zy);
• stałej resyntezy ATP z ADP i fosforanu w procesie oddychania
komórkowego;
• stałego odprowadzenia z komórek ostatecznego produktu rozpadu
substancji energetycznych — dwutlenku węgla;
• odpowiedniego stosunku kationów [Na +] do [K+] w płynie zewnątrzkomórkowym;
• odpowiedniej temperatury dla procesów enzymatycznych wewnątrz
komórkowych, która wynosi 37°C.
Wystarczy zmiana jednego z powyższych warunków, aby nastąpiło
zwolnienie lub zatrzymanie pompy sodowo-potasowej, np. na skutek
zatrzymania dopływu tlenu lub obniżenia temperatury tkanek. Po za
trzymaniu pompy następuje wyrównanie stężenia jonów Na+ i K+ po obu
stronach błony komórkowej i zanika różnica potencjałów elektrycznych
między wnętrzem komórki i otoczeniem. Proces ten przebiega wolniej,
w czasie kilkudziesięciu minut, w dużych komórkach mięśni szkieletowych i szybko, w czasie od kilku sekund do kilku minut, w komórkach
nerwowych i ich wypustkach.
Tkanki pobudliwe stanowią znaczny procent masy ciała człowieka.
W związku z tym zwolnienie lub zatrzymanie pompy sodowo-potasowej
prowadzi nie tylko do zmian składu płynu wewnątrzkomórkowego, lecz
również do zmian składu płynu zewnątrzkomorkowego, w którym stężenie
jonów Na + zmniejsza się i zwiększa stężenie jonów K +.
Po wyrównaniu różnicy potencjałów pomiędzy wnętrzem komórki i jej
otoczeniem, wskutek zatrzymania pompy, komórki tkanek pobudliwych
tracą swoje właściwości. Przestają reagować na bodźce i stają się niepo
budliwe.
I Komórka nerwowa
W organizmie człowieka jest około 1 biliona (10 12) komórek nerwowych i od kilkunastu do kilkudziesięciu razy więcej komórek podporowych dla neuronów, czyli komórek glejowych. Znaczna większość
neuronów jest skupiona w ośrodkowym układzie nerwowym. Stosun
kowo niewielka ich liczba występuje poza tym układem w zwojach
Komórka nerwowa
63
nerwowych. Są to neurony w zwojach nerwowych nerwów czaszkowych
i nerwów rdzeniowych, w zwojach nerwowych układu autonomicznego
oraz w splotach przewodu pokarmowego.
Zasadniczą funkcją neuronu jest przekazywanie informacji zakodowanych w postaci impulsów nerwowych. Przewodzenie impulsów nerwowych
przez neurony związane jest z procesami elektrochemicznymi przebiegającymi w ich błonie komórkowej.
Neurony różnią się dość znacznie pomiędzy sobą zarówno pod względem morfologicznym, jak i czynnościowym. Neurony składają się z ciała
komórkowego oraz dwóch rodzajów wypustek: jednego aksonu i licznych
dendrytów.
Ryc. 28. Komórka nerwowa: d — dendryty,
c—ciało komórki, w—wzgórek aksonu i odcinek
początkowy aksonu, a — akson z osłonką mielinową, o — gałąź oboczna aksonu, z — zakończenia aksonu.
Ciała neuronów mają różnorodny kształt i wielkość w granicach od 4
do 150 |im. Są utworzone z jądra komórkowego wraz z otaczającą go
cytoplazmą (neuroplazmą). Ciała neuronów są zasadniczym miejscem
metabolizmu i syntezy składników komórkowych. Zsyntetyzowana cytoplazmą wraz ze strukturami komórkowymi wytworzonymi w sąsiedztwie
jądra komórkowego przesuwa się do aksonów i stale przez nie przepływa
(axoplasma flow). Zsyntetyzowana cytoplazmą wraz z niektórymi organellami komórkowymi przepływa przez aksony w kierunku ortodromo-
64
Czynność komórek nerwowych i mięśniowych
wym, czyli postępującym (anterograde transport) — od ciała neuronu do
zakończeń aksonu. Wyróżniono dwa rodzaje przepływu ortodromowego
aksoplazmy:
• transport szybki, około 400 mm na dobę;
• transport wolny, od 0,5 do 10 mm na dobę.
W aksonach występuje również przepływ antydromowy aksoplazmy,
czyli wsteczny (retrograde transport), około 200 mm na dobę, mający
istotne znaczenie dla czynności neuronów. Tak jest transportowany
czynnik wzrostu nerwów (nerve growth factor — NGF), od unerwianych
narządów do ciała neuronów. Czynnik ten jest białkiem zbudowanym
z sześciu podjednostek: dwóch alpha, dwóch beta i dwóch gamma o masie
cząsteczkowej około 130 kDa. Aktywność wzrostowa związana jest
z podjednostkami beta, których budowa chemiczna zbliżona jest do
budowy insuliny.
Ryc. 29. Ciało neuronu (perykarionu). (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Komórka nerwowa
65
Ciało neuronu zazwyczaj otaczają liczne komórki należące do tkanki
podporowej, które dzielą się na komórki makrogleju i mikrogleju (patrz
str. 88).
W organizmie człowieka występują neurony o krótkich aksonach
rozgałęziających się w pobliżu ciała komórki, jak i o drugich aksonach,
z których najdłuższe mają około 1,2 m długości. Aksony rozpoczynają się
na powierzchni ciała neuronu od wzniesienia zwanego wzgórkiem aksonu,
przechodzącego następnie w odcinek początkowy aksonu. Na swoim
przebiegu niektóre długie aksony tworzą odgałęzienia w postaci gałęzi
obocznych — kolaterali. Z zewnątrz aksony otaczają komórki glejowe,
które pośredniczą w wymianie substancji odżywczych i metabolitów
pomiędzy aksonem i płynem międzykomórkowym oraz stanowią osłonę
mechaniczną.
Większość długich aksonów biegnących przez istotę białą w ośrodkowym układzie nerwowym oraz w nerwach rdzeniowych otoczona jest
przez dodatkową osłonkę mielinową. Długie aksony z osłonką mielinową,
zwaną również osłonką rdzenną, noszą nazwę włókien rdzennych w odróżnieniu od włókien bezrdzennych, czyli aksonów bez osłonki mielinowej.
Osłonka mielinową utworzona jest przez oligodendrocyty w ośrodkowym
układzie nerwowym i neurolemocyty w obwodowym układzie nerwowym,
które owijając się kilkakrotnie wokół aksonów tworzą tę osłonkę ze swojej
błony komórkowej ułożonej w kilka warstw. Spełnia ona jednocześnie
funkcję ochrony mechanicznej i izolatora elektrycznego aksonu. W odstępach około 1 mm w obrębie cieśni węzła (przewężeń Ranviera) włókna
rdzenne pozbawione są osłonek.
Ryc. 30. Akson (I) bez osłonki mielinowej i akson (II) z osłonką mielinową. Jądro lemocytu,
czyli komórki Schwanna — s.
3 — Fizjologia człowieka
66
Czynność komórek nerwowych i mięśniowych
Jedne z największych komórek nerwowych w organizmie człowieka,
0 drugich aksonach, występują w jądrach ruchowych pnia mózgowia]
1 rdzenia kręgowego. Aksony tych komórek biegną w nerwach cza
szkowych lub nerwach rdzeniowych do mięśni szkieletowych, gdzie
znajdują się ich zakończenia. Neurony małe, pośredniczące, zaopatrzone;
są w krótkie aksony, rozgałęziające się w pobliżu ciała komórkowego.
Przewodzą one impulsy nerwowe na małe odległości, pośrednicząc pomię
dzy neuronami o długich aksonach.
W warunkach prawidłowych aksony przewodzą impulsy nerwowe
tylko w jednym kierunku — od ciała neuronu do zakończeń aksonu.
W obrębie zakończeń aksonu następuje przekazywanie impulsu nerwowego na inne komórki.
Drugi rodzaj wypustek neuronu stanowią dendryty, zazwyczaj liczne,
rozgałęzione i krótsze od aksonów. Przewodzą one impulsy nerwowe
w kierunku do ciała komórkowego.
Odmienną budowę mają neurony czuciowe w zwojach rdzeniowych.
Jedna ich długa wypustka, podobna do aksonu, odbiera pobudzenie
z receptorów i przewodzi je w postaci impulsów nerwowych w kierunku
ciała komórkowego. Za pośrednictwem krótszej wypustki zaś, wstępującej do rdzenia kręgowego, przekazuje impulsy do innych komórek
nerwowych.
I Potencjał czynnościowy
Bodziec działając na błonę komórkową neuronu zmienia jej właściwości, co z kolei wywołuje potencjał czynnościowy (action potential). Do
wnętrza neuronu przez otwierające się kanały dla prądu jonów sodowych,
napływają jony Na +, co powoduje wyrównanie ładunków elektrycznych
pomiędzy wnętrzem i otoczeniem. Zjawisko to określa się jako depolaryzację błony komórkowej. Jony Na + początkowo wnikają do wnętrza
neuronu tylko w miejscu zadziałania bodźca. Z chwilą wyrównania
ładunków elektrycznych w tym jednym miejscu depolaryzacja zaczyna się
rozszerzać na sąsiednie odcinki błony komórkowej, przesuwając się
również wzdłuż aksonów. Impulsem nerwowym (neural impulse) jest
przesuwanie się fali depolaryzacji od miejsca zadziałania bodźca na błonę
komórkową, aż do zakończeń neuronu.
W organizmie impulsy nerwowe przekazywane są z jednej komórki
nerwowej na drugą za pośrednictwem zakończeń aksonów. Miejsce
stykania się ze sobą błony komórkowej zakończenia aksonu z błoną
Komórka nerwowa
67
komórkową drugiej komórki nosi nazwę synapsy. Błonę komórkową
neuronu przekazującego impuls przyjęto nazywać błoną presynaptyczną
(presynaptic membranę), błona komórkowa neuronu odbierającego impuls nosi zaś nazwę błony postsynaptycznej (postsynaptic membranę).
Postsynaptyczny potencjał pobudzający
Z zakończeń aksonów w obrębie synapsy wydzielają się przekaźniki
chemiczne, czyli transmittery, które zmieniają właściwości błony postsynaptycznej.
Pod wpływem cząsteczek transmittera, który pośredniczy w przewodzeniu przez synapsę impulsów pobudzających, jony Na + wnikają do
wnętrza neuronu odbierającego impuls nerwowy. Tej wędrówce jonów
Na+ przez błonę postsynaptyczną towarzyszy zmniejszenie spoczynkowego potencjału ujemnego wewnątrz komórki w stosunku do potencjału
zerowego panującego na zewnątrz komórki. Zmniejszenie się ujemnego
potencjału wewnątrzkomórkowego przyjęto nazywać postsynaptycznym
potencjałem pobudzającym — EPSP (excitatory postsynaptic potential),
synapsy wywołujące depolaryzację błony błony komórkowej zaś noszą
nazwę synaps pobudzających.
Sumowanie impulsów przestrzenne i w czasie
Stopień depolaryzacji błony postsynaptycznej zależy od liczby cząsteczek transmittera wydzielonych na synapsach. Im więcej cząsteczek
transmittera wiąże się z receptorami w błonie postsynaptycznej, tym
większy jest stopień depolaryzacji. Postsynaptyczny potencjał pobudzający (EPSP) zależy od liczby cząsteczek wydzielanych na poszczególnych synapsach oraz od liczby synaps, na których jednocześnie wydziela
się transmitter. Im więcej synaps jest pobudzonych jednocześnie, tym
wyższy jest potencjał pobudzający. Wzrastanie postsynaptycznego potencjału
pobudzającego (EPSP) w miarę zwiększania się liczby synaps przekazujących pobudzanie wskazuje na zjawisko sumowania się przestrzennego
impulsacji (spatial summation) w obrębie neuronu. Neuron i otaczające go
synapsy zajmują pewną trójwymiarową przestrzeń i dlatego ten typ
sumowania impulsów nosi nazwę sumowania przestrzennego.
Impulsy nerwowe są przewodzone przez komórki nerwowe nie pojedynczo, lecz zazwyczaj w postaci salw. W każdej salwie odstępy pomiędzy
impulsami mogą się zmniejszać, dochodzi wtedy do przyspieszenia często-
68
Czynność komórek nerwowych i mięśniowych
tliwości przewodzonych impulsów, lub zwiększać — następuje wówczas1
zwalnianie częstotliwości przewodzenia impulsów.
Impulsy nerwowe przewodzone przez synapsy w odstępach krótszych |
od 5 ms trafiają na resztki depolaryzacji wywołanej poprzednim impul-l
sem. Kolejne, występujące po sobie postsynaptyczne potencjały pobudza-1
jące (EPSP) częściowo nakładają się na siebie i coraz bardziej depola ryzują błonę komórkową. W ten sposób w obrębie neuronu dochodzi do
sumowania się impulsów występujących w krótkich odstępach czasu, czyli
do sumowania w czasie (temporal summation).
Ryc. 31. Postsynaptyczny potencjał pobudzający (EPSP). (I) Sumowanie impulsów przestrzenne: EPSP wywołany uwalnianiem transmittera przez jedną synapsę (1) i przez trzy
synapsy jednocześnie (x3). (II) Sumowanie impulsów w czasie: błona presynaptyczna
depolaryzuje się i uwalnia transmitter w odstępach 5 ms (u góry) i 3 ms (u dołu).
Potencjał iglicowy
Pod wpływem transmittera wydzielającego się na synapsach pobudzających depolaryzacja błony komórkowej neuronu osiąga pewien próg,
przy którym dochodzi do wyzwolenia potencjału iglicowego (spike poten-
Komórka nerwowa
69
tial). Potencjał progowy (firing level), czyli krytyczny potencjał błony
komórkowej ciała neuronu, wynosi — 55 mV. Po przekroczeniu tego progu
na skutek sumowania przestrzennego, czy też sumowania w czasie
impulsów, postsynaptyczny potencjał pobudzający przechodzi w potencjał iglicowy. Charakteryzuje się on szybko narastającą depolaryzacją
błony komórkowej. Potencjał wewnątrzkomórkowy z ujemnego zmienia
się na dodatni, osiągając na swoim szczycie +35 mV. Ten bardzo krótko
trwający dodatni potencjał wewnątrzkomórkowy określa się jako nadstrzał (overshoot).
Po szybko narastającej depolaryzacji zakończonej nadstrzałem następuje repolaryzacja błony komórkowej, w pierwszej fazie równie szybko
jak depolaryzacja. Potencjał iglicowy w zależności od typu komórek
nerwowych trwa od 0,5 do 2 ms.
Ryc. 32. Potencjał czynnościowy wnętrza neuronu: EPSP — postsynaptyczny potencjał
pobudzający, SP — potencjał iglicowy, ADP — podepolaryzacyjny potencjał następczy,
HAP — hiperpolaryzacyjny potencjał następczy.
70
Czynność komórek nerwowych i mięśniowych
Potencjał następczy
Repolaryzacja błony komórkowej neuronu ulega zwolnieniu po po-1
wrocie ujemnego potencjału wewnątrzkomórkowego i po przekroczeniu i
potencjału progowego. W tej fazie zjawisko to określa się jako potencjał I
następczy (after potential), w czasie którego błona komórkowa nie osiąga I
spoczynkowej polaryzacji ( — 70 mV). Nosi to nazwę: podepolaryzacyjnego I
potencjału następczego (after-depolarization potential). Potem potencjał I
następczy przekracza wartość spoczynkową, wnętrze komórki staje się I
bardziej ujemne, co określa się mianem hiperpolaryzacyjnego potencjału
następczego (after-hyperpolarization potential). Po czym ładunek elektryczny wnętrza komórki powraca całkowicie do wartości spoczynkowej
( — 70 mV). W czasie występowania potencjału iglicowego, a więc w zakresie powyżej potencjału progowego, błona komórkowa jest niewrażliwa
na bodźce. Jest to okres niepobudliwości bezwzględnej, czyli okres refrakcji
bezwzględnej (absolute refractory period). Po osiągnięciu potencjału |
progowego pobudliwość powraca i jest wzmożona w okresie podepolaryzacyjnego potencjału następczego, co trwa 4 ms. W okresie hiperpolaryzacyjnego potencjału następczego trwającego 35—40 ms pobudliwość
błony komórkowej jest zmniejszona. -
Powstawanie impulsów w aksonach
Potencjał iglicowy w obrębie ciała neuronu ruchowego w warunkach
prawidłowych występuje rzadko. Natomiast w obrębie wzgórka (axon
hillock) i odcinka początkowego aksonu (initial segment of axon) są
wytwarzane potencjały iglicowe przewodzone następnie w postaci pojedynczych impulsów lub całych salw aż do zakończeń aksonów. Potencjał
progowy wzgórka i odcinka początkowego aksonu jest bardziej zbliżony
do średniego potencjału spoczynkowego. Różnica wynosi około 15 mV
[( — 70 mV) — ( — 55 mV) = —15 mV]. W stosunku do ciała neuronu
różnica ta jest większa i wynosi około 20 mV [(— 70 mV) — (— 50 mV) =
= -20 mV].
Pobudliwość błony komórkowej neuronów zależy od gęstości kanałów
dokomórkowego prądu jonów Na + bramkowanych napięciem. W błonie
komórkowej wzgórka i odcinka początkowego aksonu gęstość tych
kanałów jonowych jest znacznie większa w stosunku do ich gęstości
w obrębie dendrytów i ciała komórkowego. Dzięki temu wzgórek i odcinek początkowy aksonu łatwiej generują potencjał iglicowy niż ciało
komórki. Są bardziej pobudliwe.
Komór k a ner w ow a
71
Ryc. 33. Potencjał czynnościowy neuronu ruchowego: EPSP — postsynaptyczny potencjał
pobudzający ciała neuronu, SP — potencjały czynnościowe części początkowej aksonu.
I — potencjał spoczynkowy, II — potencjał progowy części początkowej aksonu, III —
potencjał progowy ciała neuronu.
Postsynaptyczny potencjał hamujący
Poza synapsami pobudzającymi otaczającymi neurony istnieją również
synapsy, na których wydziela się transmitter hamujący przewodzenie
impulsów. Cząsteczki tego transmittera zmieniają właściwości błony
komórkowej w ten sposób, że jony K + uciekają z wnętrza neuronu na
zewnątrz i jednocześnie jony Cl wnikają do wnętrza przez odpowiednie
kanały jonowe. Ubywanie jonów o dodatnim ładunku el ektrycznym
i przybywanie jonów o ujemnym ładunku powoduje wzrost ujemnego
potencjału elektrycznego we wnętrzu komórki do około — 80 mV i wzrost
polaryzacji błony komórkowej. Ten stan hiperpolaryzacji błony komórkowej określa się jako postsynaptyczny potencjał hamujący — IPSP
(inhibitory postsynaptic potential).
72
Czynność komórek nerwowych i mięśniowych
Ryc. 34. Oddziaływanie transmitterów uwolnionych z zakończeń presynaptycznych na
błonę postsynaptyczną: I — synapsy pobudzające depolaryzują błonę komórkową i powstaje EPSP, II — synapsy hamujące hiperpolaryzują błonę komórkową i powstaje IPSP.
III — synapsy aksono-aksonalne (AA) zmniejszają zawartość transmittera pobudzającego
w synapsach aksonalno-somatycznych (AS) i nie występuje depolaryzacja błony komórkowej.
Synapsy wydzielające transmitter hiperpolaryzujący błonę komórkową
są synapsami hamującymi. Neurony, w których występuje postsynaptyczny potencjał hamujący, mają zmniejszoną pobudliwość. Ujemny potencjał elektryczny ich wnętrza jest bardziej oddalony od progu, przy którym wyzwalają się impulsy.
Neurony w organizmie stale odbierają jednocześnie impulsy przewodzone przez synapsy pobudzające i hamujące. Potencjał błony komórkowej neuronu jest wypadkową działania tych dwóch typów synaps.
W czasie przewagi przewodzenia impulsów przez synapsy pobudzające,
Komórka nerwowa
73
potencjał błony komórkowej zbliża się do progu (— 50 mV) i w tym samym
czasie w odcinku początkowym aksonu generowana jest salwa impulsów.
Przeciwnie, przewaga synaps hamujących hiperpolaryzuje błonę komórkową i uniemożliwia generowanie impulsów w odcinku początkowym
aksonu.
I Synapsy
Powierzchnię błony komórkowej ciała neuronu i dendrytów pokrywają
w około 40% struktury zwane kolbami końcowymi — synaptycznymi
(synaptic knobs). Błona komórkowa wzgórka aksonu i odcinka początkowego aksonu pokryta jest niewielką liczbą kolb końcowych. Kolby są
zakończeniami aksonów i dzięki nim odbywa się przekazywanie impulsów
z jednego neuronu na drugi.
Kolby końcowe są niewielkie, o średnicy około 1 um. Pokrywa je
błona presynaptyczna należąca do neuronu przekazującego impuls. Pomiędzy błoną presynaptyczna i błoną komórkową neuronu odbierającego
impulsy, zwaną błoną postsynaptyczną, istnieje szczelina synaptyczna
(synaptic cleft) o szerokości od 15 do 50 nm, średnio 20 nm.
Wewnątrz kolb synaptycznych znajdują się mitochondria oraz twory
0 średnicy od 20 do 60 nm zwane pęcherzykami synaptycznymi (synaptic
vesicles). Zagęszczenie pęcherzyków wewnątrz kolb zwiększa się w miej
scu stykania się błony presynaptycznej z błoną postsynaptyczną. Pęcherzy
ki synaptyczne zawierają transmittery i modulatory chemiczne, które
w czasie przewodzenia impulsu przez synapsę uwalniają się z pęcherzyków
do szczeliny synaptycznej i wiążą z receptorami postsynaptycznymi
1
presynaptycznymi. Transmittery i modulatory odczepiają się od
recep
torów nie zmienione. Metabolizowane są przez enzymy na nieaktywne
związki lub, na drodze internalizacji receptorów ze związanymi transmitterami i modulatorami, dostają się do wnętrza komórek postsynaptycznych i presynaptycznych.
Synapsy częściej przewodzące impulsy mają większe zagęszczenie
pęcherzyków synaptycznych i eksternalizowanych receptorów w sąsiedztwie szczeliny synaptycznej niż synapsy rzadko przewodzące impulsy.
Przez kilkadziesiąt lat powszechnie uznawane było tzw. prawo Dale'a,
zgodnie z którym komórka nerwowa uwalnia na swych synapsach tylko
jeden przekaźnik chemiczny. Później wykazano, że komórka nerwowa
może uwalniać na swych zakończeniach synaptycznych jednocześnie
74
Czynność komórek nerwowych i mięśniowych
Ryc. 35. Transmittery i modulatory z pęcherzyków synaptycznych dostają się do prze strzeni synaptycznej i po związaniu się z receptorami oraz po ich internalizacji przechodzą
do cytoplazmy zarówno komórek postsynaptycznych, jak i komórek presynaptycznych.
kilka przekaźników chemicznych o silnym biologicznym działaniu. Można
je podzielić umownie na transmittery synaptyczne — związki o małej
cząsteczce, i na modulatory synaptyczne — związki o większej cząsteczce,
do których zalicza się kilkadziesiąt peptydów. Przekaźniki chemiczne
nazwane umownie transmitterami i modulatorami są nie tylko wytwarzane
i uwalniane przez komórki nerwowe, ale również przez inne komórki,
szczególnie komórki wydzielania wewnętrznego.
Komórka nerwowa
75
Transmittery pobudzające
Do tej grupy związków należą transmittery chemiczne depolaryzujące
błonę postsynaptyczną. Po ich związaniu się z receptorem w błonie
postsynaptycznej dochodzi do tzw. aktywacji sodowej — otwierają się
kanały dla dokomórkowego prądu jonów sodowych. Po 0,5 ms następuje
inaktywacja sodowa z jednoczesną aktywacją potasową — wychodzenie
jonów potasowych z komórek w czasie repolaryzacji.
Do transmitterów chemicznych pobudzających zalicza się acetylocholinę, aminy (dopamina, noradrenalina, serotonina), adenozynę oraz aminokwasy pobudzające (sole kwasu asparaginowego i glutaminowego).
Ryc. 36. Synteza acetylocholiny (Ach) w zakończeniach cholinergicznych z choliny
(Ch) i rodnika acetylowego (A), jej rozpad pod wpływem enzymów i wychwyt zwrotny
choliny (Ch) z krwi krążącej.
76
Czynność komórek nerwowych i mięśniowych
Acetylocholina (Ach — acetylcholine) jest syntetyzowana w zakońl
czeniach cholinergicznych z choliny i kwasu octowego, w czym bierze
udział enzym acetylotransferaza cholinowa. Cząsteczki acetylocholiny
uwolnione z pęcherzyków synaptycznych do przestrzeni synaptycznej!
w wyniku egzocytozy wiążą się z receptorem cholinergicznym w błonie!
postsynaptycznej i otwierają się kanały dla dokomórkowego prądu!
jonów Na +. Receptor cholinergiczny jest białkiem błony komórkowej!
zbudowanym z czterech podjednostek (alpha, beta, gamma i delta)]
o masie cząsteczkowej 250 kDa. Wyróżnia się dwa receptory cholinergiczne — nikotynowy i muskarynowy (tab. 3).
Ryc. 37. Gromadzenie się noradrenaliny (NA) w zakończeniach adrenergicznych, wsteczny
wychwyt z przestrzeni synaptycznej i z krwi krążącej oraz powstawanie jej metabolitów
pod wpływem enzymów: metylotransferazy katecholowej (COMT) i oksydazy monoaminowej (MAO).
Tabela 3
Transmittery synaptyczne, ich receptory, działania na kanary jonowe w błonie postsynaptycznej, tworzenie się drugiego wewnątrzkomórkowego przekaźnika i metabolizm komórkowy. Otwieranie się kanałów dla prądu jonowego dokomorkowego — T, prądu jonowego
odkomórkowego — TT, zwiększenie stężenia w komórkach drugiego przekaźnika — T i aktywacja cyklazy adenylanowej — ( + ). Zamykanie się kanałów dla prądu jonowego
dokomorkowego — 1, prądu jonowego odkomórkowego — li i inaktywacja cyklazy adenylanowej — (—).
78
Czynność komórek nerwowych i mięśniowych
W błonie postsynaptycznej, na którą działa Ach, znajduje się enzym-I
swoista esteraza cholinowa (AChE — specific acetylcholine esterase) rozkładający acetylocholinę na cholinę i kwas octowy. W osoczu i w in-1
nych tkankach występują enzymy rozkładające Ach. Są to nieswoista
esterazy cholinowe (non specific cholinesterase, pseudocholinesterase). I
Dopamina (dopamine) i noradrenalina (noradrenaline, norepinephrinem
stanowią ogniwa w łańcuchu przemian aminokwasu fenyloalaniny (patrzl
ryc. 114, str. 217). Dopamina działa poprzez pięć receptorów od D: do D5.|
Działanie poprzez receptor D x powoduje aktywację enzymu — cyklazy
adenylanowej zależnej od dopaminy (dopamine-sensitive adenylate cyc-1
łase). Noradrenalina wywiera wpływ na komórki poprzez receptory!
adrenergiczne alpha i beta. Serotonina — 5-HT (5-hydroxytryptaminę) —jest metabolitem aminokwasu tryptofanu i wywiera swój wpływ!
za pośrednictwem odpowiednich receptorów serotoninoergicznych.
Aminokwasy pobudzające — sole kwasu glutaminowego (glutamicl
acid) i kwasu asparaginowego (aspartic acid) — działają na neurony!
postsynaptyczne za pośrednictwem receptorów metabolitotropowych i re-l
ceptorów jonotropowych.
Działanie metabolitotropowe aminokwasów pobudzających związane I
jest z aktywacją enzymów wewnątrzkomórkowych zależnych od inozoty-1
lotrifosforanu (IP 3) i diacyloglicerolu (DAG). Działanie jonotropowe I
polega na otwieraniu się jednego typu kanałów dla dokomorkowego prądu I
jonów Na + po związaniu się tych aminokwasów z receptorem AMPA I
(receptorem a-amino-3-hydroksy-5-metylo-4-izoksazolopropionianu) lub I
z receptorem kainianowym. Po związaniu się aminokwasów pobudzających
z receptorem NMDA (receptorem N-metylo-D-asparaginianu) otwierają
się jednocześnie dwa inne typy kanałów dla dokomorkowego prądu jonów
Na+ i Ca2 + .
Trzy czwarte transmisji pobudzającej w mózgowiu stanowi transmisja
za pośrednictwem aminokwasów pobudzających. Mają one działanie nie
tylko jako przekaźniki synaptyczne, ale wykazują również wpływ cytotoksyczny na neurony. W czasie ostrego niedotlenienia całego mózgowia
lub jego poszczególnych struktur są one uwalniane z neuronów i wykazują
działanie niszczące, cytotoksyczne.
Do transmitterów synaptycznych zalicza się tlenek azotu — NO (nitric
oxide), który spełnia również inne funkcje w organizmie. W komórkach
nerwowych powstają z aminokwasu argininy pod wpływem mózgowej
izoformy enzymu syntazy tlenku azotu (brain nitric oxide synthase
isoform). Tlenek azotu uwolniony z błony postsynaptycznej działa na
błonę presynaptyczną jako wsteczny przekaźnik (retrograde messenger),
ułatwiając uwalnianie innego trasmittera do przestrzeni synaptycznej.
Komórka nerwowa
79
Transmitter hamujący
Postsynaptyczny potencjał hamujący (IPSP), czyli hiperpolaryzacja
błony postsynaptycznej, powstaje pod wpływem transmittera hamującego, jakim jest kwas gamma-aminomasłowy — GABA (gamma-aminobutyric acid). Tworzy się w neuronach w wyniku dekarboksylacji aminokwasu — kwasu glutaminowego (glutamic acid). Wiąże się z dwoma
receptorami GABAA i GABAB. Związanie się GABA z receptorem
GABAA powoduje otwarcie się kanałów dokomórkowego prądu jonów
Cl" i zwiększenie ujemnego potencjału w neuronach. Działanie GABA za
pośrednictwem drugiego receptora GABAB jest odmienne. Powoduje
otwieranie się kanałów odkomórkowego prądu jonów K+, zamykanie się
kanałów dokomórkowego prądu jonów Ca 2+ i zwiększenie stężenia
w neuronach drugiego przekaźnika — inozytolotrifosforanu (IP3) i diacyloglicerolu (DAG).
W tabeli 3 wymienione są transmittery synaptyczne, receptory błonowe, z którymi się wiążą, oraz wywołane przez nie reakcje jonotropowe
i metabolitotropowe.
Modulatory synaptyczne
Z pęcherzyków synaptycznych w czasie egzocytozy uwalniają się poza
transmitterami również związki o większych cząsteczkach, którym przypisuje się rolę modulatorów synaptycznych lub kotransmitterów (cotransmitters). Do tych związków zalicza się biologicznie aktywne peptydy.
Modulatory synaptyczne uwolnione z zakończeń nerwowych oddziałują
na błonę postsynaptyczną i presynaptyczną. Aktywują lub inaktywują
enzymy występujące w tych błonach, wpływają na internalizację i eksternalizację receptorów błonowych, tym samym zmieniają właściwości błon
komórkowych, wzmacniając lub tłumiąc działanie transmitterów. Ostateczny ich wpływ na przewodnictwo synaptyczne jest wypadkową licznych procesów zachodzących na synapsach. Ten sam modulator może
działać torujące lub tłumiąco w zależności od miejsca, w którym jest
uwalniany.
Nazwy modulatorów synaptycznych często odnoszą się do miejsca,
w którym po raz pierwszy je odkryto i wykazano ich biologiczne działanie. Wcześniej niż w pęcherzykach synaptycznych wykryto ich występowanie w podwzgórzu i w przysadce, przypisując im wyłącznie
działanie hormonalne, oraz w przewodzie pokarmowym jako hormony
Tabela 4
Modulatory i miejsce ich występowania w ośrodkowym i obwodowym układzie nerwowym
Komórka nerwowa
81
żołądkowo-jelitowe wydzielane przez rozsiane w organizmie komórki
wydzielania wewnętrznego.
Osobną grupę modulatorów stanowią peptydy opioidowe działające
na komórki za pośrednictwem receptorów opioidowych (mu, delta, kappa).
Peptydy te działają jednokierunkowo na przewodnictwo synaptyczne
tłumiące je. Prekursorami peptydów opioidowych są: pre-pro-enkefalina,
pre-pro-dynorfina i pre-pro-opiomelanokortyna. Z pre-pro-enkefaliny
o cząsteczce zbudowanej z 263 aminokwasów powstaje po fragmentacji
6 cząsteczek enkefaliny metioninowej (Met-enkephalin) i 1 cząsteczka
enkefaliny leucynowej (Leu-enkephalin). Z pre-pro-dynorfiny B (256 aminokwasów) po enzymatycznej fragmentacji powstaje alpha-neo-endorfina
(alpha-neo-endorphin) i dynorfina 1—17 (dynorphin) lub dynorfina 1—8.
Z pre-pro-opiomelanokortyny (265 aminokwasów) tworzą się w komórkach części pośredniej przysadki cząsteczki hormonów melanotropowych: alpha, beta i gamma (melanocyte stimulating hormones), enkefalina metioninowa oraz cząsteczka peptydu kortykotropowopodobnego
(corticotropin-like intermediate lobe peptide — CLIP) stanowiąca fragment hormonu kortykotropowego — ACTH 18—39.
Poza peptydami również adenozynotrifosforany i prostaglandyny modulują przewodnictwo synaptyczne, ale mechanizm ich uwalniania i działania jest odmienny w porównaniu do peptydów.
I Kontrola ekspresji genów
Przepływ informacji między komórkami nerwowymi nie ogranicza się
do procesów zachodzących na błonie komórkowej, lecz dotyczy mechanizmów syntezy transmitterów i modulatorów. W komórkach nerwowych
stale zachodzi postępujący (anterograde) przepływ substancji chemicznych od perykarionu do końców wypustek i wsteczny (retrograde) — od
wypustek do perykarionu. Substancje chemiczne, przede wszystkim peptydy, są wstecznie transportowane do perykarionu z przestrzeni synaptycznej lub płynu międzykomórkowego, po związaniu się z receptorem
w błonie presynaptycznej i po internalizacji.
Po wniknięciu do jądra komórkowego oddziałują na aparat genetyczny. Dochodzi do przyspieszenia lub opóźnienia procesu transkrypcji
mRNA dla polipeptydów spełniających rolę modulatorów synaptycznych
i dla białek receptorowych i kanałowych lub enzymów uczestniczących
w syntezie transmitterów. Dzięki kontroli ekspresji genów w aparacie
genetycznym jądra komórkowego neurony wzajemnie oddziałują na siebie.
82
Czynność komórek nerwowych i mięśniowych
Nadrzędne neurony przewodzą impulsy nerwowe, przekazując je za
pośrednictwem synaps na neurony podrzędne. Te neurony zaś, uwalniając
związki biologicznie czynne, przede wszystkim peptydy, wstecznie modulują procesy syntezy transmitterów i modulatorów w komórkach nadrzędnych.
I Hamowanie presynaptyczne
W zależności od umiejscowienia na błonie postsynaptycznej neuronu
odbierającego synapsy dzielą się na trzy zasadnicze rodzaje. Są to I
zakończenia synaptyczne aksonów na błonie postsynaptycznej:
• dendrytów, zwane synapsą aksono-dendrytyczną;
• ciała neuronu, zwane synapsą aksono-somatyczną;
• aksonów, zwane synapsą aksono-aksonalną.
Neurony kończące się synapsami pobudzającymi mogą również hamować przekazywanie impulsów przez inne neurony pobudzające na drodze
hamowania presynaptycznego (presynaptic inhibition). Ten typ hamowania wywołują synapsy aksono-aksonalne. Impulsy nerwowe przewodzone
przez synapsy pierwszego aksonu, znajdujące się w pobliżu zakończeń
drugiego aksonu, depolaryzują jego błonę presynaptyczną i zmniejszają
liczbę uwalnianych cząsteczek transmittera pobudzającego. Tym samym
impulsy przewodzone przez ten drugi akson nie depolaryzują błony
postsynaptycznej.
I
Przewodzenie impulsów we włóknach
bezrdzennych
W warunkach fizjologicznych błona komórkowa odcinka początkowego aksonu depolaryzuje się pod wpływem potencjału pobudzającego
występującego w ciele neuronu. W odcinku początkowym aksonu dochodzi do generowania impulsu nerwowego przesuwającego się następnie ortodromowo — w kierunku od ciała komórki do zakończeń aksonu.
Włókno nerwowe w organizmie, jak i wyizolowane z organizmu, można
również pobudzić, działając bodźcem na dowolny jego odcinek. W tym
wypadku impuls nerwowy rozchodzi się od miejsca pobudzenia w obu
kierunkach, do ciała komórki — antydromowo, i do zakończeń aksonu — ortodromowo.
Komórka nerwowa
83
Ryc. 38. Błona komórkowa ciała neuronu i odcinka początkowego aksonu: a — w spoczynku; b — w czasie depolaryzacji ciała neuronu; c — w czasie przewodzenia depolaryzacji wzdłuż aksonu bez osłonki mielinowej.
Pod wpływem postsynaptycznego potencjału pobudzającego, występującego w ciele neuronu, otwierają się kanały dla dokomórkowego prądu
jonów Na + w błonie komórkowej aksonu. Po przekroczeniu potencjału progowego liczba jonów Na + napływających do wnętrza aksonu
jest tak duża, że nie tylko błona komórkowa depolaryzuje się, lecz także
pojawia się potencjał iglicowy o dodatnim ładunku elektrycznym, czyli
nadstrzale.
84
Czynność komórek nerwowych i mięśniowych
Ruch jonów Na+ do wnętrza aksonu odciąga je z sąsiadującej zewnętrznej powierzchni błony komórkowej aksonu, powodując jej depolaryzację. Dzięki, temu we włóknach bez osłonki mielinowej fala depolaryzacji przesuwa się w sposób ciągły.
Na szczycie potencjału iglicowego dalszy napływ jonów Na + do wnętrza aksonu zostaje zahamowany na skutek inaktywacji kanału dokomór
kowego prądu jonów sodowych. Jednocześnie rozpoczyna się proces otwierania się kanałów dla odkomórkowego prądu jonów K + i ucieczki na
zewnątrz aksonu dodatnio naładowanych jonów potasowych. To zaś prowadzi do repolaryzacji błony komórkowej aksonu. Po przejściu impulsu
nerwowego pompa sodowo-potasowa przywraca koncentrację jonów Na +
i K+ wewnątrz aksonu do stanu poprzedzającego przewodzenie impulsu.
Przewodzenie impulsu nerwowego we włóknach bezrdzennych bez
osłonki mielinowej polega na przesuwającej się w sposób ciągły fali
depolaryzacji, za którą postępuje repolaryzacja. Prędkość przewodzenia
impulsów we włóknach bezrdzennych jest nieduża i wynosi od 0,5 do 2 m/s,
potencjał iglicowy zaś trwa średnio 2 ms.
I
Przewodzenie impulsów we włóknach
rdzennych
Depolaryzacja odcinka początkowego aksonu jest spowodowana gwałtownym napływem jonów sodowych do wnętrza aksonu i ruchem tych
jonów w całym otoczeniu. Powoduje to otwieranie się kanałów dla
dokomórkowego prądu jonów Na + bramkowanych napięciem i depolaryzację błony aksonu w obrębie cieśni węzłów. Następnie depolaryzacja
skokowa (saltatory conduction) obejmuje odcinki błony komórkowej
w coraz to dalszych cieśniach węzłów, w których jest największa gęstość
kanałów jonowych.
Osłonka mielinowa spełnia funkcję izolatora i w miejscach, w których
otacza ona akson, jest mała gęstość kanałów jonowych i nie występuje
ruch jonów przez błonę komórkową. Dzięki temu depolaryzacja błony
komórkowej aksonu może przeskakiwać od cieśni do cieśni węzła,
pomijając odcinki aksonu otoczone osłonką mielinowa.
Przeskakiwanie depolaryzacji pomiędzy cieśniami węzłów powoduje
skokowe przewodzenie impulsów nerwowych we włóknach rdzennych,
które jest znacznie szybsze niż we włóknach bezrdzennych. W zależności od średnicy aksonu prędkość przewodzenia we włóknach rdzennych wynosi od kilku do 120 m/s. Im większa średnica aksonu, tym
Komórka nerwowa
85
szybciej włókna te przewodzą impulsy nerwowe. Potencjał iglicowy
w tych włóknach trwa około 0,5 ms. Dzięki temu włókna drażnione
prądem elektrycznym mogą przewodzić do 2000 impulsów na sekundę.
W organizmie człowieka impulsy przewodzone są we włóknach nerwowych na zmianę z narastającą i zmniejszającą się częstotliwością.
Dzięki temu tworzą one salwy impulsów. W pojedynczym włóknie
nerwowym unerwiającym komórki mięśni szkieletowych impulsy wystę-
Na*
c
Ryc. 39. Błona komórkowa ciała neuronu i odcinka początkowego aksonu: a — w spoczynku; b — w czasie depolaryzacji ciała neuronu; c — w czasie przewodzenia skokowego
depolaryzacji wzdłuż aksonu z osłonką mielinową.
86
Czynność komórek nerwowych i mięśniowych
pują z częstotliwością od 60 do 80 Hz. W czasie maksymalnego skur
mięśnia częstotliwość impulsów w pojedynczym włóknie nerwowym osią
ga 150 Hz.
I Grupy włókien nerwowych
Włókna nerwowe dzieli się zarówno pod względem morfologicznym, jak
i czynnościowym. Morfologiczne kryteria podziału odnoszą się do wy
stepowania lub niewystępowania osłonki mielinowej, do średnicy aksonów oraz do umiejscowienia zarówno w ośrodkowym układzie nerwowym, gdzie tworzą drogi nerwowe, jak i obwodowym układzie nerwowym, gdzie są skupione w postaci nerwów.
Pod względem czynnościowym włókna nerwowe dzielą się na przewodzące impulsy z obwodu do ośrodków — są to włókna dośrodkowe,
czyli aferentne, i z ośrodków na obwód — włókna odśrodkowe, czyli
Tabela 5
Podział włókien nerwowych w zależności od ich budowy i czynności
Nerwy czaszkowe i nerwy rdzeniowe
87
eferentne. Różnią się także szybkością przewodzenia impulsów, czasem
trwania potencjału iglicowego, długością okresu bezwzględnej niewrażliwości, czyli refrakcji (refractory period), oraz charakterem
transmitterów i modulatorów uwalnianych z zakończeń aksonów.
Uwzględniając zarówno kryteria morfologiczne, jak i czynnościowe
dzieli się włókna nerwowe na cztery grupy:
• włókna nerwowe grupy A — mają osłonkę mielinową i są to zarówno
włókna aferentne — przewodzące czucie, jak i eferentne — soma
tyczne. W zależności od średnicy aksonów dzielą się na podgrupy: alpha
(średnicy 12—20 um), beta (średnicy 5—12 um), gamma (średnicy 3—6
Hm) i delta (średnicy 2—5 um);
• włókna nerwowe grupy B — mają osłonkę mielinową, należą do układu
autonomicznego, a na ich zakończeniach uwalnia się acetylocholina
(ACh). Do tej grupy zalicza się włókna przywspółczulne oraz współczulne przedzwojowe, do których należą np. gałęzie łączące białe
(rami communicantes albi);
• włókna nerwowe Cs (sympathetic C fibres) — nie mają osłonki
mielinowej, na ich zakończeniach uwalnia się noradrenalina (NA).
Są to włókna współczulne zazwojowe, np. gałęzie łączące szare (rami
communicantes grisei);
• włókna nerwowe C d.r. (dorsal root C fibres) — nie mają osłonki
mielinowej. Są to włókna aferentne wstępujące do rdzenia kręgowego
przez korzenie grzbietowe (radices dorsales medullae spinalis). Prze
wodzą one niektóre rodzaje czucia takie, jak czucie bólu, oraz sta
nowią część aferentną trzewnych łuków odruchowych.
Włókna aferentne należące zarówno do grupy A, jak i C d.r. przewodzą impulsy od receptorów do ośrodkowego układu nerwowego. Tym
samym stanowią część łuków odruchowych zarówno somatycznych, jak
i autonomicznych.
Włókna eferentne należące do grupy A i B opuszczają ośrodkowy
układ nerwowy w nerwach czaszkowych (oprócz nerwu: I, II i VIII)
oraz nerwach rdzeniowych.
I
Nerwy czaszkowe i nerwy
rdzeniowe
W organizmie człowieka znajduje się 12 par nerwów czaszkowych oraz
31 par nerwów rdzeniowych, poprzez które włókna nerwowe wchodzą lub
opuszczają ośrodkowy układ nerwowy.
88
Czynność komórek nerwowych i mięśniowych
Nerwy rdzeniowe (nervi spinałeś) o włóknach aferentnych i eferentnych dzielą się na:
• 8 nerwów szyjnych (nervi cervicales) Cl—C8;
• 12 nerwów piersiowych (nervi thoracici) Thl—Thl2;
• 5 nerwów lędźwiowych {nervi lumbales) LI—L5;
• 5 nerwów krzyżowych (nervi sacrales) Sl—S5;
• 1 nerw guziczny (nervus coccygeus).
We wszystkich bez wyjątku nerwach czaszkowych i rdzeniowych!
biegną włókna aferentne, pokryte osłonką mielinową lub bez osłonki!
mielinowej. Włókna eferentne opuszczające ośrodkowy układ nerwowy!
biegną do:
• mięśni szkieletowych poprzecznie prążkowanych i są zaliczane do
układu somatycznego
lub
• mięśni gładkich i komórek gruczołowych, stanowiąc część układu]
autonomicznego.
Włókna nerwowe układu somatycznego znajdują się w nerwach czaszkowych: III, IV, V, VI, VII, IX, X, XI i XII oraz we wszystkich nerwach!
rdzeniowych. Włókna nerwowe należące do układu autonomicznego
(systema nervosum autonomicum) dzielą się na przywspółczulne (pars
parasympathica) oraz współczulne (pars sympathica). Włókna przywspółczulne biegną w nerwach czaszkowych: III, VII, IX i X oraz w nerwach
krzyżowych S2—S4. Włókna współczulne opuszczają rdzeń kręgowy
w nerwach piersiowych Thl—Thl2 i w nerwach lędźwiowych LI—L3.
I Komórki glejowe
Drugi rodzaj komórek występujących w układzie nerwowym ośrodkowym i obwodowym są to komórki glejowe. Są one kilkakrotnie
liczniejsze w stosunku do komórek nerwowych. Komórki glejowe nie
przewodzą impulsów nerwowych, lecz stanowią tkankę podporową dla
komórek nerwowych, wychwytują i metabolizują transmittery uwolnione
do przestrzeni synaptycznej. W ten sposób wpływają na przewodnictwo
synaptyczne. Komórki glejowe dzielą się na neuroglej i mezoglej (określany
również jako mikroglej). Do neurogleju należą: astrocyty, oligodendrocyty, ependymocyty w ośrodkowym układzie nerwowym i gliocyty oraz
neurolemocyty (komórki Schwanna) w obwodowym układzie nerwowym.
Mięśnie poprzecznie prążkowane
89
Oligodendrocyty wytwarzają mielinę spełniającą funkcję izolatora oddzialającego od siebie komórki nerwowe. Wypustki komórek nerwowych wychodzące z ośrodkowego układu nerwowego w postaci włókien
nerwowych są również otoczone izolacją w postaci osłonki miel inowej wytwarzanej przez komórki zwane neurolemocytami (komórki
Sch wanna).
Astrocyty są to duże komórki glejowe mające liczne wypustki. Część
wypustek otacza naczynia włosowate krwionośne, inne wypustki zaś
stykają się z komórkami nerwowymi. W ten sposób astrocyty, oddzielając komórki nerwowe od bezpośredniego kontaktu z naczyniami włosowatymi krwionośnymi, jednocześnie pośredniczą w wymianie substancji
budulcowych (aminokwasy), energetycznych i produktów przemiany
materii (metabolity transmitterów synaptycznych) pomiędzy krwią i komórkami nerwowymi. Odprowadzają także jony potasowe z płynu międzykomórkowego z otoczenia repolaryzujących się neuronów.
Drugi rodzaj komórek glejowych stanowią małe komórki, czyli
komórki mikrogleju. Wykazują właściwości żerne, otaczając miejsca
uszkodzone w ośrodkowym układzie nerwowym i pożerając fragmenty
obumarłych komórek nerwowych. Wytwarzają również interleukinę 1
(IL-1) — czynnik o budowie peptydowej wywołujący odczyn gorączkowy.
I Mięśnie poprzecznie prążkowane I
Komórka mięśniowa
Mięsień poprzecznie prążkowany, czyli szkieletowy, jest zbudowany
z wielu tysięcy komórek mięśniowych tworzących pęczki. W zależności
od długości samego mięśnia jego komórki mają od kilku milimetrów do
około 50 cm długości. Na obu końcach komórki mięśniowe są przyczepione do ścięgien.
Komórka mięśniowa poprzecznie prążkowana (myocytus transversostriatus) jest wielojądrzastą komórką, cylindryczną na przekroju poprzecznym, o średnicy około 50 um. Komórka mięśniowa otoczona jest sarkolemą — pobudliwą błoną komórkową. Wnętrze komórki wypełnia
sarkoplazma i pęczki włókienek mięśniowych.
Wlókienko mięśniowe, czyli miofibryla, ma odcinki o większym i mniejszym współczynniku załamania światła występujące naprzemiennie. Odcinki silniej załamujące światło tworzą ciemniejsze prążki zwane prążkami anizotropowymi (A), odcinki słabiej załamujące światło tworzą zaś
90
Czynność komórek nerwowych i mięśniowych
jasne prążki izotropowe (I). W miofibrylach położonych obok siebie
prążki anizotropowe i izotropowe sąsiadują ze sobą i w ten sposób
tworzą poprzeczne prążkowanie całej komórki mięśniowej.
Włókienko mięśniowe składa się z grubych i cienkich nitek białek
kurczliwych. Nitkę grubą tworzą cząsteczki miozyny (m.cz. 480 kDa),
z których każda zbudowana jest z dwóch ciężkich i czterech lekkich
łańcuchów polipeptydowych. Łańcuchy ciężkie splecione ze sobą ślimakowato z jednego końca tworzą ogon, niesplecione zaś z drugiego końca
mają kształt dwóch głów. W każdej głowie występują również dwa
łańcuchy lekkie.
Nitka cienka utworzona jest z cząsteczek aktyny (m.cz. 43 kDa)
i tropomiozyny (m.cz. 70 kDa), tworząc jak gdyby cztery sznury paciorków skręconych ślimakowato (patrz ryc. 43, str. 94). Na tropomiozynie
osadzone są cząsteczki troponiny, z których każda ma trzy podjednostki (T, I, C) o odmiennych właściwościach. Za pośrednictwem podjednostki T troponina łączy się z tropomiozyną. Podjednostka I ma
duże powinowactwo do aktyny, podjednostka C zaś do jonów wapniowych.
Sarkomer
Sarkomer obejmuje jeden cały prążek anizotropowy i sąsiadujące
z nim dwie połówki prążka izotropowego. Prążek anizotropowy tworzą
nitki grube miozyny, prążek izotropowy zaś nitki cienkie aktyny, które
są doczepione do błony granicznej Z (telophragma). Błona Z dzieli na
dwie połówki każdy prążek izotropowy należący do dwóch sąsiednich
sarkomerów. Nitki aktyny doczepione do błony granicznej Z w postaci
grzebieni wchodzą pomiędzy nitki miozyny. Każda z nitek miozyny jest
otoczona sześcioma nitkami aktyny.
W czasie skracania się komórki mięśniowej nitki aktyny tworzące
prążki izotropowe wsuwają się pomiędzy nitki miozyny i dzięki temu
wszystkie prążki I nikną. Podczas rozkurczu nitki aktyny wysuwają się
spomiędzy nitek miozyny i prążki I ponownie stają się widoczne (ryc. 40).
Mięśnie poprzecznie prążkowane
91
Ryc. 40. Sarkomer w rozkurczu i w skurczu. Prążki A — anizotropowe, prążki I — izotropowe.
Układ sarkotubularny
Układ sarkotubularny jest strukturą komórkową pośredniczącą w przenoszeniu pobudzenia wewnątrz całej komórki mięśniowej. Układ ten
składa się z cewek poprzecznych (tubuli transversi) i siateczki sarkoplazmatycznej {reticulum sarcoplasmaticum). Końce cewek poprzecznych
dochodzą do błony komórkowej, wewnątrz komórki mięśniowej zaś
znajdują się pomiędzy miofibrylami na granicy prążków I i A.
Miofibryle otacza siateczka sarkoplazmatyczna. Siateczka ta rozciąga
się wzdłuż miofibryli prostopadle do cewek poprzecznych. W miejscach
92
Czynność komórek nerwowych i mięśniowych
stykania się siateczki sarkoplazmatycznej z cewką poprzeczną siateczka
tworzy zbiornik końcowy (cisterna terminalis), w którym znajdują się
jony wapniowe w dużym stężeniu. W czasie depolaryzacji błony cewek
poprzecznych następuje ruch ładunków elektrycznych, pod wpływem
których otwierają się w błonie zbiorników końcowych kanały jonów
wapniowych i jony te napływają pomiędzy nitki białek kurczliwych, wiążąc
Ryc. 41. Miofibryla zbudowana z nitek miozyny i nitek aktyny przymocowanych do
błony granicznej Z otoczona jest cewkami poprzecznymi dochodzącymi do sarkolemy
i zbiornikami końcowymi siateczki sarkoplazmatycznej.
Mięśnie poprzecznie prążkowane
93
się z podjednostką C troponiny. W czasie rozkurczu komórki mięśniowej
pompa wapniowa w błonie zbiorników końcowych ponownie gromadzi
w nich jony wapniowe.
Mechanizm molekularny skurczu
Pod wpływem bodźca fizjologicznego, którym w przypadku komórek
mięśniowych poprzecznie prążkowanych jest acetylocholina uwolniona na
synapsach nerwowo-mięśniowych, dochodzi do pobudzenia błony komórkowej, czyli do jej depolaryzacji. Pod wpływem acetylocholiny błona
komórkowa zmienia swoje właściwości. Dochodzi do aktywacji w niej
kanałów dla dokomórkowego szybkiego prądu jonów sodowych.
Depolaryzacja przesuwa się po powierzchni błony komórkowej komórek mięśniowych poprzecznie prążkowanych i jednocześnie za pośred-
Acetylocholina
Nitki aktyny
wślizgują się
noTnędzy nitki miozyny
Ryc. 42. Mechanizm molekularny skurczu komórki mięśniowej poprzecznie prążkowanej.
94
Czynność komórek nerwowych i mięśniowych
nictwem cewek poprzecznych obejmuje wnętrze komórki. Ze zbiorników
końcowych uwalniają się wolne jony wapniowe. Jony wapniowe wiążą się
z podjednostką C troponiny i zmniejszają jej powinowactwo do aktyny.
Cząsteczki aktyny uwolnione od hamującego wpływu troponiny stykają się z głowami cząsteczek miozyny, wyzwalając jej aktywność enzymatyczną. Pod wpływem aktywnej miozyny adenozynotrifosforany
(ATP) rozkładane są do adenozynotrifosforanów (ADP) i fosforanu.
Głowy cząsteczek miozyny, stykając się z cząsteczkami aktyny, hydrolizują ATP i zmieniają swoją konformację względem nitki miozyny
(ryc. 43). Następnie natychmiast powracają do poprzedniego położenia.
Stykają się z innymi, dalej położonymi cząsteczkami aktyny i ponownie
zmieniają swoją konformację względem nitki miozyny. Dzięki temu
wsuwają się nitki cienkie aktyny pomiędzy nitki grube miozyny.
Ślizgowe nasuwanie się nitek cienkich aktyny na nitki grube miozyny,
powoduje skracanie się komórki mięśnia poprzecznie prążkowanego
i skurcz całego mięśnia. Czerpanie z hydrolizy ATP energii niezbędnej
Ryc. 43. Mechanizm ślizgowy skurczu komórki mięśniowej — wsuwania się nitek aktyny
(n.a.) pomiędzy nitki miozyny (n.m.): a — rozkurcz — głowy cząsteczek miozyny (1) nie
wykazują aktywności enzymatycznej i nie stykają się z cząsteczkami aktyny (2) hamowanymi przez podjednostkę I troponiny (3) przymocowanej do tropomiozyny (4); b —
skurcz — głowy cząsteczek miozyny (1) hydrolizują ATP, zmieniają swoją konformację
i przesuwają nitkę aktyny.
Mięśnie poprzecznie prążkowane
95
dla czynności mechanicznej komórki mięśniowej nosi nazwę sprzężenia
mechaniczno-chemicznego (mechano-chemical coupling).
Nitki cienkie aktyny wsunięte są pomiędzy nitki grube miozyny tak
długo, jak długo wolne jony wapniowe oddziałują na podjednostkę C
troponiny. W czasie rozkurczu pompa wapniowa w błonie zbiorników
końcowych wciąga wolne jony Ca 2+ do zbiorników końcowych i nitki
cienkie wysuwają się spomiędzy nitek grubych. Następna depolaryzacja
błony komórkowej komórki mięśniowej powoduje ponowne otwarcie się
kanałów wolnego prądu jonów wapniowych i ponowne wsuwanie się
nitek cienkich pomiędzy nitki grube. Proces ten nosi nazwę sprzężenia
elektromechanicznego (excitation-contraction coupling).
I Skurcze mięśni
Pod wpływem działającego na komórkę mięśniową pojedynczego
bodźca o sile progowej lub większej od progowej, jej błona komórkowa
ulega depolaryzacji, po której następuje skurcz całej komórki. Bodziec
podprogowy nie wywołuje depolaryzacji błony komórkowej i komórka
mięśniowa nie kurczy się. Komórka mięśniowa odpowiada na bodziec
zgodnie z prawem „wszystko albo nic", to znaczy pod wpływem każdego bodźca o intensywności progowej lub większej od progowej reaguje
maksymalnie, nie odpowiada zaś na bodźce podprogowe.
Depolaryzacja błony komórkowej komórki mięśniowej trwa 1—3 ms.
W tym czasie błona komórkowa jest niewrażliwa na bodźce i ich nie
odbiera. Jest to okres bezwzględnej niewrażliwości (absolute refractory
period). Po fazie depolaryzacji następuje faza repolaryzacji, w czasie
której potencjał elektryczny wewnątrz komórki mięśniowej powraca do
stanu wyjściowego, tj. do stanu w okresie spoczynku przed działaniem
bodźca.
Depolaryzacja błony komórkowej wyprzedza o kilka milisekund początek skracania się mięśnia, czyli jego skurcz. Skurcz pojedynczy w mięśniach szybko kurczących się trwa krótko, około 7,5 ms, w mięśniach
wolno kurczących się trwa zaś znacznie dłużej — do 100 ms. Po skurczu
mięśnia następuje jego rozkurcz.
Wyróżnia się dwa rodzaje pojedynczych skurczów mięśni szkieletowych: izotoniczne i izometryczne. W czasie skurczu izotonicznego (isotonic contraction) komórki mięśniowe skracają się i cały mięsień ulega
skróceniu, jego napięcie zaś nie zmienia się. Przyczepy mięśnia w układzie
szkieletowym zbliżają się do siebie. Skurcz izometryczny (isometric con-
96
Czynność komórek nerwowych i mięśniowych
Ryc. 44. Skurcz pojedynczy mięśnia poprzecznie prążkowanego: a — skurcz izotoniczny;
b — skurcz izometryczny. Bodziec (B) wyzwala czynnościowy potencjał elektryczny
w komórce oraz zmianę długości mięśnia {a) lub napięcia mięśnia (b).
traction) charakteryzuje się zwiększeniem napięcia mięśnia bez zmian
jego długości. Przyczepy mięśnia w układzie szkieletowym nie zmieniają
swojej odległości.
Powtarzające się bodźce nadprogowe z pewną częstotliwością wywołują kolejne skurcze mięśnia. Przy pewnej częstotliwości bodźca zaczyna
występować zjawisko sumowania się skurczów pojedynczych (summation
of contractions). Dochodzi do skurczu tężcowego zupełnego (complete
tetanic contraction) lub do skurczu tężcowego niezupełnego (incomplete
tetanic contraction) (ryc. 45). Skurcz tężcowy zupełny występuje wtedy,
kiedy bodźce pobudzają mięsień w odstępach czasu krótszych niż trwa
skurcz pojedynczy. Pobudzenie mięśnia w odstępach czasu dłuższych
niż czas trwania skurczu pojedynczego pozwala na częściowy rozkurcz
mięśnia pomiędzy bodźcami.
Mięśnie szybko kurczące się wymagają ponad stu bodźców na sekundę,
aby wystąpił skurcz tężcowy zupełny. W mięśniach wolno kurczących się
wystarcza kilkanaście bodźców na sekundę, aby uzyskać skurcz tężcowy
zupełny.
W czasie skurczu tężcowego izotonicznego mięsień skraca się, jego
napięcie zaś nie ulega zmianie. Skurcz tężcowy izometryczny charakteryzuje się zwiększeniem napięcia mięśnia, któremu nie towarzyszy
zmniejszenie jego długości. Ruchy kończyn i całego ciała są spowo -
Mięśnie poprzecznie prążkowane
97
Ryc. 45. Skurcz tężcowy mięśnia poprzecznie prążkowanego: a — skurcz tężcowy niezupełny, bodziec o częstotliwości 25 Hz; b — skurcz tężcowy zupełny, bodziec o częstotliwości 50 Hz. Zmiany potencjału elektrycznego wnętrza komórki i długości mięśnia.
dowane przede wszystkim skurczami tężcowymi mięśni szkieletowych
o typie skurczów auksotonicznych, czyli z jednoczesnym zbliżeniem przyczepów i zwiększeniem napięcia.
Mięsień nie rozciągnięty kurczy się z małą siłą. W miarę rozciągania
mięśnia skurcze stają się coraz silniejsze aż do optymalnego rozciągnięcia,
przy którym mięsień kurczy się z maksymalną siłą. Dalsze rozciąganie
mięśnia powoduje stopniowe zmniejszenie się siły skurczów.
Słabe bodźce działające na mięsień pobudzają do skurczu tylko część
komórek mięśniowych. Zwiększając stopniowo siłę bodźca, uzyskuje się
coraz silniejsze skurcze. Pobudzenie wszystkich komórek mięśniowych
wywołuje skurcz maksymalny.
4 — Fizjologia człowieka
98
Czynność komórek nerwowych i mięśniowych
Energetyka skurczu mięśnia
Bezpośrednim źródłem energii potrzebnej do skurczów mięśnia szkieletowego jest adenozynotrifosforan (ATP). Rozkłada się on w czasie
skurczu do adenozynodifosforanu (ADP) i fosforanu. Energia do resyntezy ATP czerpana jest w procesie metabolizowania składników
odżywczych aż do końcowych produktów, tj. do dwutlenku węgla i wody.
Całkowity rozpad podstawowego składnika odżywczego, jakim jest glukoza, do końcowych produktów metabolizmu wewnątrzkomórkowego
dostarcza najwięcej energii do resyntezy ATP. Dzieje się to w czasie
glikolizy tlenowej, kiedy prężność tlenu w komórce (Po 2) jest dostateczna.
Glikoliza tlenowa
+
1 mol glukozy + 02 ------------- —— ----- =^-—- C02 H2 0
W czasie szybko narastającego wysiłku fizycznego dowóz tlenu do
komórek mięśniowych nie nadąża za zapotrzebowaniem na energię i P o,
w komórkach znacznie się zmniejsza. Dochodzi wtedy do dysocjacji
mioglobiny (barwnika zbliżonego budową i właściwościami do hemoglobiny), która uwalnia związany tlen. Jest to jedyny magazyn tlenu
wewnątrzkomórkowego, z którego komórka może go czerpać w czasie
zwiększonego zapotrzebowania na energię lub zatrzymania krążenia krwi.
Jednakże ilość tlenu związanego z mioglobiną jest niewielka w porównaniu z zapotrzebowaniem na niego. W tym stanie energia potrzebna do
resyntezy ATP jest czerpana w procesie glikolizy beztlenowej oraz z hydrolizy fosfokreatyny.
AD P+ P
ATP
Hydroliza fosfokreatyny:
Fosfokreatyna+ADP^ATP+Kreatyna
Ryc. 47. Glikoliza beztlenowa.
Mięśnie poprzecznie prążkowane
99
Czerpanie energii potrzebnej do resyntezy ATP w fazie beztlenowej jest
znacznie mniej wydajne w porównaniu z fazą tlenową i jest ograniczone
w czasie ze względu na zmniejszenie wartości pH w komórce na skutek
gromadzenia się mleczanów.
Większa część energii (od 75 do 80%) wytworzonej w komórce
mięśniowej z metabolizowania glukozy jest zamieniana na energię cieplną.
Na energię mechaniczną związaną ze skracaniem się mięśnia jest wykorzystane tylko od 20 do 25% energii wytworzonej w komórce mięśniowej.
Energia cieplna powstaje w komórce mięśniowej w:
• procesie spoczynkowego metabolizmu wewnątrzkomórkowego (resting
heat);
• czasie reakcji chemicznych związanych z aktywowaniem miozyny (activation heat);
• czasie skracania się komórki mięśniowej (shortening heat);
• procesie rozkurczu i wydłużania się komórki mięśniowej (relaxation
heat);
• procesie odnowy związanej z resyntezą ATP i działaniem pompy
sodowo-potasowej (recovery heat).
Jednostka motoryczna
Komórki mięśni poprzecznie prążkowanych szkieletowych są unerwiane przez komórki nerwowe skupione w jądrach ruchowych pnia
mózgowia i rdzenia kręgowego. Komórki nerwowe jąder ruchowych
wysyłają swoje wypustki — aksony — do komórek mięśniowych. Pojedyncza komórka nerwowa unerwia średnio około 150 komórek mięśniowych. Mięśnie stale pracujące i wykonujące precyzyjne ruchy, np. mięśnie
gałek ocznych, są kontrolowane przez znaczną liczbę komórek nerwowych. Jeden neuron unerwia średnio tylko 10 komórek mięśniowych.
W mięśniach siłowych zaś, utrzymujących wyprostną postawę ciała,
na jedną komórkę nerwową przypada znacznie więcej komórek mięśniowych. Jeden neuron unerwia średnio 200 komórek mięśniowych.
Jedna komórka nerwowa, jej wypustka, biegnąca do mięśnia i wszystkie komórki mięśniowe przez nią unerwiane stanowią jednostkę motoryczna (motor unit).
W warunkach prawidłowych w organizmie człowieka pojedyncze
skurcze mięśni szkieletowych prawie nie występują. Ruchy kończyn i ruchy
całego ciała są spowodowane skurczami tężcowymi izotonicznymi i izometrycznymi oraz auksotonicznymi.
1 00
Czynność komórek nerwowych i mięśniowych
Siła skurczu mięśnia w organizmie zależy od:
• liczby jednostek motorycznych biorących udział w skurczu;
• częstotliwości, z jaką poszczególne jednostki motoryczne są pobu
dzane;
• stopnia rozciągnięcia mięśnia przed jego skurczem.
Jednostki motoryczne każdego mięśnia szkieletowego nie pracują
synchronicznie. W związku z tym stopniowanie siły skurczu lub jej
zmniejszenie odbywa się w sposób ciągły, prawie niedostrzegalny. W czasie słabego skurczu niewielki procent jednostek motorycznych wykazuje pobudzenie występujące z małą częstotliwością, niesynchronicznie.
W miarę narastania pobudzenia, które obejmuje coraz większą liczbę
jednostek motorycznych, pracujących z coraz większą częstotliwością —
również niesynchronicznie — zwiększa się siła skurczu.
I Regulacja napięcia mięśniowego
Neurony ruchowe skupione w jądrach ruchowych pnia mózgowia
i rdzenia kręgowego stale wysyłają impulsy do mięśni szkieletowych.
Nawet mięśnie nie pracujące wykazują napięcie mięśniowe spoczynkowe.
Jest' to związane z pobudzeniem niewielkiej liczby jednostek motorycznych. Tym samym napięcie mięśniowe jest to słaby skurcz tężcowy
izometryczny bardzo długo utrzymujący się.
Napięcie mięśniowe jest regulowane:
• przez nadrzędne ośrodki ruchowe w ośrodkowym układzie nerwowym;
• dzięki samoregulacji (muscle tonus regulation).
Samoregulacja napięcia mięśniowego
Rozciągnięcie mięśnia wyzwala odruchowe zwiększenie jego napięcia.
Na przykład skurcz izotoniczny mięśni zginaczy powoduje jednoczesne
rozciągnięcie mięśni antagonistycznych, czyli prostowników, i zwiększenie ich napięcia mięśniowego na drodze odruchowej.
W mięśniach szkieletowych występują dwa rodzaje komórek mięśniowych: komórki ekstrafuzalne i komórki intrafuzalne.
Komórki mięśniowe ekstrafuzalne (extrafusal fibres) mają jednolitą
budowę na całej długości, są skupione w pęczki i oba ich końce są
przyczepione do ścięgien.
Mięśnie poprzecznie prążkowane
1 01
Komórki mięśniowe intrafuzalne (intrafusal fibers) w swej części środkowej nie mają poprzecznego prążkowania i część ta nie kurczy się. Są
skupione w pęczki, czyli wrzecionka nerwowo-mięśniowe (fusus neuromuusculańs), które otacza torebka łącznotkankowa. Wrzecionka nerwowomięśniowe przyczepiają się swoimi końcami do komórek ekstrafuzal-nych.
We wrzecionkach nerwowo-mięśniowych znajdują się receptory wrażliwe
na rozciąganie mięśnia.
Komórki mięśniowe ekstrafuzalne, które stanowią podstawową masę
każdego mięśnia szkieletowego, unerwione są przez duże neurony ruchowe, zwane neuronami alpha. Komórki intrafuzalne unerwiane są przez
mniejsze neurony ruchowe, zwane neuronami gamma. Pobudliwość receptorów wrażliwych na rozciąganie we wrzecionkach nerwowo-mięśniowych jest regulowana przez te neurony. Im silniej są pobudzone neurony
gamma i komórki intrafuzalne skurczone, tym bardziej zwiększa się
wrażliwość receptorów na rozciąganie.
Pobudzone receptory we wrzecionkach nerwowo-mięśniowych na skutek rozciągania mięśnia wysyłają impulsy do neuronów alpha w jądrach
ruchowych pnia mózgowia lub rdzenia kręgowego. Z kolei pobudzone
neurony alpha wysyłają impulsy do komórek ekstrafuzalnych i kurczą je.
Skurcz izotoniczny komórek ekstrafuzalnych zmniejsza pobudliwość
receptorów we wrzecionkach nerwowo-mięśniowych, co prowadzi do
zmniejszenia napięcia tego mięśnia. Natomiast skurcz izometryczny ko-
Ryc. 48. Wrzecionko nerwowo-mięśniowe. W nerwie mięśniowym biegną
włókna dosrodkowe (aferentne): pierwotne — od zakończeń pierścieniowato-spiralnych, i wtórne — od zakończeń w postaci „wiązanki kwiatów",
oraz włókna odśrodkowe (eferentne):
kończące się zakończeniami synaptycznymi nerwowo-mięśniowymi w odcinkach poprzecznie prążkowanych komórek śródwrzecionkowych (intrafuzalnych). (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd.
Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
1 02
Czynność komórek nerwowych i mięśniowych
Ryc. 49. „Pętla mięśniowa", dzięki której utrzymana jest stała kontrola napięcia mięśniowego. Impulsy krążą pomiędzy y-motoneuronami, wrzecionkami nerwowo-mięśniowymi
(A) i a-motoneuronami. B — komórki mięśniowe ekstrafuzalne, S — receptory skórne.
mórek ekstrafuzalnych nie zmniejsza pobudliwości receptorów i napięcie
mięśniowe utrzymuje się. Impulsy nerwowe stale krążące pomiędzy
receptorami we wrzecionkach nerwowo-mięśniowych, motoneuronami
w jądrach ruchowych i komórkami mięśniowymi zapewniają odpowiednie
napięcie wszystkich mięśni szkieletowych dostosowane do pozycji całego
ciała oraz ustawienie kończyn i głowy w stosunku do tułowia. Impulsy
krążące po „zamkniętej pętli" sprzężenia zwrotnego (muscles-spinal cord
feedback loop) pomiędzy rdzeniem kręgowym a mięśniami utrzymują
samoregulację napięcia mięśniowego (patrz Odruch monosynaptyczny,
str. 123).
I Synapsa nerwowo-mięśniowa
W każdej jednostce motorycznej występuje jedna komórka nerwowa,
która łączy się z komórkami mięśniowymi za pomocą zakończeń aksonu.
Miejsce stykania się wypustki komórki nerwowej — aksonu — z komórką
mięśniową nosi nazwę zakończenia synaptycznego nerwowo-mięśniowego
(terminatio synapsis neuromusculariś).
Mięśnie poprzecznie prążkowane
1 03
Włókno nerwowe typu A w pobliżu unerwianej komórki mięśniowej
traci osłonkę mielinową i rozdziela się na wiele stopek końcowych (pes
terminalis). Wszystkie stopki końcowe tworzą strukturę zwaną zakończeniem synaptycznym nerwowo-mięśniowym. Błona komórkowa komórki mięśniowej jest w miejscu stykania się z płytką końcową zagłębiona i pofałdowana, tworzy rynienki otaczające poszczególne stopki
końcowe. Pomiędzy błoną presynaptyczną otaczającą stopki końcowe
i pofałdowaną błoną postsynaptyczną komórki mięśniowej występuje
niewielka przestrzeń nosząca nazwę szczeliny synaptycznej (fissura synapticd).
Impuls nerwowy, przesuwając się wzdłuż włókna nerwowego, obejmuje
stopki końcowe, depolaryzując ich błonę presynaptyczną. Pod wpływem
depolaryzacji otwiera się do szczeliny synaptycznej pewna liczba pęcherzyków synaptycznych zgromadzonych w stopkach końcowych. Z pęcherzyków synaptycznych uwalnia się acetylocholina (Ach), co jest nazywane
sprzężeniem elektrowydzielniczym.
Acetylocholina wiążąc się z receptorem w błonie postsynaptycznej,
otwiera kanały dla dokomórkowego szybkiego prądu jonów
sodowych i jony te wnikają do wnętrza komórki mięśniowej. Błona
komórkowa komórki mięśniowej w miejscu zetknięcia się z
zakończeniem
synaptycznym
nerwowo-mięśniowym
zostaje
zdepolaryzowana, co powoduje wahania potencjału elektrycznego w tej
okolicy. Zjawisko to nosi nazwę potencjału zakończenia synaptycznego
nerwowo-mięśniowego (endplate potential).
Mięsień poprzecznie prążkowany Ryc. 50. Budowa schematyczna
zakończenia synaptycznego nerwowo-mięśniowego.
1 04
Czynność komórek nerwowych i mięśniowych
Komórka
t
Komórka
mięśniowa --------------- -mięśniowa
rozkurczona
skurczona
Ryc. 51. Procesy zachodzące w obrębie synapsy nerwowo-mięśniowej w czasie przewodzenia impulsu.
Depolaryzacja błony postsynaptycznej i pojawienia się potencjału
zakończenia synaptycznego nerwowo-mięśniowego wymaga uwolnienia
do szczeliny synaptycznej 10 amol acetylocholiny. Tym samym przewodzenie przez synapsę nerwowo-mięśniową jednego impulsu wymaga
wydzielania do szczeliny synaptycznej od 1000 do 10 000 cząsteczek Ach
w około 200 punktach błony presynaptycznej.
Potencjał zakończenia synaptycznego nerwowo-mięśniowego rozchodzi się wzdłuż komórki mięśniowej, depolaryzując jej błonę komórkową,
co wyzwala skurcz.
Receptory w błonie postsynaptycznej wrażliwe na Ach mogą być
zablokowane przez związek wprowadzony do organizmu z zewnątrz —
kurarę.
W czasie działania kurary Ach wydzielona do szczeliny synaptycznej
nie łączy się z receptorem i nie wywołuje depolaryzacji błony postsynaptycznej. Przewodzenie nerwowo-mięśniowe jest zablokowane.
W błonie postsynaptycznej znajduje się enzym rozkładający Ach
uwolnioną do szczeliny synaptycznej. Enzymem tym jest esteraza choli-
Mięśnie poprzecznie prążkowane
1 05
Ryc. 52. Synapsa nerwowo-mięśniowa. Rk — receptor wiążący acetylocholinę (Ach)
blokowany przez kurarę, RACIIE — receptor wiążący acetylocholinę blokowany przez
atropinę.
nowa (AChE), rozkładająca acetylocholinę i kwas octowy. Związki
hamujące działanie esterazy cholinowej — inhibitory esterazy cholinowej — zwiększają wrażliwość błony postsynaptycznej na działanie Ach.
Mniejsza liczba cząsteczek Ach jest w stanie zdepolaryzować błonę
postsynaptyczną i tym samym wywołać skurcz komórki mięśniowej.
Jony wapniowe i magnezowe oddziałują również na przewodnictwo
nerwowo-mięśniowe. Jon wapniowy jest niezbędny w procesie przewodzenia przez synapsę nerwowo-mięśniową, gdyż warunkuje uwalnianie
cząsteczek Ach z zakończenia synaptycznego nerwowo-mięśniowego. Jon
magnezowy ma działanie przeciwne — hamuje uwalnianie Ach.
1 06
Czynność komórek nerwowych i mięśniowych
I Mięśnie gładkie
Komórka mięśniowa gładka {myocytus glaber) w odróżnieniu od komórki mięśniowej poprzecznie prążkowanej nie ma jednostek kurczliwych w postaci sarkomerów. Wnętrze komórki mięśniowej gładkiej
wypełnione jest nitkami kurczliwymi ułożonymi równolegle i biegnącymi
wzdłuż długiej osi komórki. W czasie skurczu nitki te skracają się na
skutek nasuwania się cząsteczek aktyny na cząsteczki miozyny. Występująca w cytoplazmie kalmodulina łączy się z napływającymi przez
dokomórkowe kanały błonowe jonami wapniowymi i aktywuje właściwości enzymatyczne jednego z łańcuchów lekkich w głowie cząsteczki
miozyny. Następuje hydroliza ATP i zmiana konformacji głowy cząsteczki
miozyny w stosunku do nitki grubej miozyny. W wyniku tego nitki cienkie
aktyny przesuwają się wzdłuż nitek grubych miozyny.
Komórki mięśniowe gładkie występują w organizmie w dwóch różnych
pod względem czynnościowym skupieniach. Tworzą:
• wielojednostkowe mięśnie gładkie (multi-unit smooth muscle) — w któ
rych poszczególne komórki kurczą się niezależnie i pobudzenie nie
przenosi się z jednej komórki na drugą. Występują w ścianach naczyń
krwionośnych i w tęczówce;
• trzewne mięśnie gładkie (single-unit smooth muscle) — stanowią
warstwy lub pierścienie równolegle ułożonych komórek, w których
pobudzenie przenosi się z jednej komórki na drugą dzięki połączeniom
szczelinowym (gap junctions). Tworzą tym samym czynnościowe syncytia. Występują w ścianach przewodu pokarmowego, w moczowodach, pęcherzu moczowym i macicy.
Komórki trzewnych mięśni gładkich spełniają dwa typy czynności
mechanicznej: skurcze i zmiany napięcia. Oba typy czynności mechanicznej mogą występować jednocześnie lub niezależnie od siebie.
Wnętrze komórek mięśniowych gładkich ma ujemny potencjał elektryczny wynoszący w spoczynku średnio — 50 mV. Zwiększenie pobudliwości wyraża się długotrwałym zmniejszeniem ujemnego potencjału
wewnątrzkomórkowego. To zmniejszenie potencjału spowodowane jest
otwieraniem się w błonie komórkowej kanałów jonów wapniowych, co
prowadzi do częściowej długotrwałej depolaryzacji na skutek zwiększonego dokomórkowego prądu jonów Ca 2 +.
Komórka o zmniejszonej pobudliwości ma wnętrze bardziej ujemne,
osiągając —65 mV (hiperpolaryzacja), i jest całkowicie rozkurczona.
Mięśnie gładkie
1 07
Ryc. 53. Potencjał elektryczny wnętrza komórki i długość mięśnia gładkiego. Mięsień gładki:
I — w czasie przeciętnej pobudliwości, II — pobudzany w czasie zmniejszonej pobudliwości,
III — w czasie wzmożonej pobudliwości.
Hiperpolaryzacja błony komórkowej spowodowana jest ucieczką jonów
K+ z wnętrza komórki lub utrudnionym wchodzeniem jonów Ca 2+ do
komórki.
Skurcz komórek mięśniowych gładkich poprzedza:
• potencjał czynnościowy iglicowy trwający około 50 ms;
• potencjał iglicowy przechodzący w plateau depolar yzacji, co trwa
łącznie od 100 ms do 1 s.
W komórkach częściowo zdepolaryzowanych skurcz nie jest poprzedzany przez potencjał czynnościowy.
Skurcz poprzedzany przez potencjał czynnościowy rozpoczyna się po
upływie około 200 ms od początku depolaryzacji i osiąga swoje maksimum po upływie 500 ms.
Mięśnie gładkie kurczą się po wpływem:
• samoistnego pobudzenia, które występuje rytmicznie w niektórych
komórkach trzewnych mięśni gładkich. Szerzy się na komórki sąsiednie,
doprowadzając cały mięsień do skurczu. Komórki te spełniają funkcję
rozrusznika dla pozostałych komórek. Wyzwalają samoistne skurcze
z częstotliwością od 0,2 Hz, tj. jednego skurczu na 5 s, do jednego
skurczu na kilka minut;
1 08
Czynność komórek nerwowych i mięśniowych
• czynnika miejscowego mechanicznego lub chemicznego działającego
bezpośrednio na komórki, np. rozciąganie mięśnia, zmiany pH, zwięk
szenie prężności dwutlenku węgla;
• przekaźników chemicznych wytworzonych w odległych tkankach
i przenoszonych drogą humoralną — kontrola humoralna, np. hor
mony rdzenia nadnerczy;
• przekaźników chemicznych wydzielonych z aksonów neuronów nale
żących do układu autonomicznego, czyli na zasadzie kontroli nerwo
wej za pośrednictwem uwalnianych transmitterów. Aksony neuronów
układu autonomicznego biegną pomiędzy komórkami mięśni gładkich,
tak że uwalniając transmitter, działają jednocześnie na kilka komórek
(fr. synapsę en passant).
Na komórki mięśniowe gładkie najczęściej działają dwa transmittery:
• noradrenalina (NA) — wydzielana na zakończeniach neuronów zazwojowych należących do układu współczulnego (pars sympathetica
systematis autonomia);
• acetylocholina (Ach) — wydzielana na zakończeniach neuronów układu
przywspółczulnego (pars parasytnpathetica systematis autonomici).
Różna jest odpowiedź narządów efektorowych na działanie transmitterów układu autonomicznego (patrz tab. 9, str. 223). Na przykład
trzewne mięśnie gładkie przewodu pokarmowego rozkurczają się pod
wpływem noradrenaliny, kurczą zaś pod wpływem acetylocholiny.
POBUDLIWOŚĆ
Ryc. 54. Potencjał elektryczny wnętrza komórki i długość mięśnia gładkiego. Mięsień gładki:
I — pobudzany w czasie przeciętnej pobudliwości, II — pobudzany w czasie wzmożonej
pobudliwości.
Mięśnie gładkie
1 09
Noradrenalina działa na komórki mięśni gładkich za pośrednictwem
dwóch receptorów błonowych: alpha i beta receptorów adrenergicznych.
Po związaniu się noradrenaliny z receptorami alpha, jony Ca2+ wychodzą
z komórek mięśniowych, co powoduje ich rozkurcz. Pod wpływem
noradrenaliny, działającej za pośrednictwem receptora beta, zostaje zaktywowana cyklaza adenylanowa, zwiększa się zawartość cyklicznego
AMP w komórkach, jony Ca2+ są wiązane wewnątrz komórek i stężenie
wolnych jonów Ca2+ w komórkach zmniejsza się. Następuje rozkurcz
komórek mięśni gładkich.
Acetylocholina po związaniu się z receptorami muskarynowymi M2
i M4 wywołuje aktywację fosfolipazy C w błonie komórkowej i uwalnianie się inozytolotrifosforanu (IP 3). Zwiększenie zawartości IP 3 w cytoplazmie wywołuje zwiększenie stężenia wolnych jonów Ca2 + w komórkach
mięśni gładkich i ich skurcz.
W organizmie zazwyczaj jednocześnie działają na komórki mięśni
gładkich oba transmittery układu autonomicznego (NA i Ach). Stan
mięśni gładkich jest wypadkową ich antagonistycznego działania. Na
trzewne mięśnie gładkie przewodu pokarmowego noradrenalina działa
rozkurczająco, acetylocholina zaś kurcząco. Przeciwnie — na wielojedno-
Ryc. 55. Wpływ noradrenaliny (NA) na komórkę mięśnia gładkiego w zależności od wzajemnego stosunku ilościowego alpha- i beta-receptorów w błonie komórkowej.
110
Czynność komórek nerwowych i mięśniowych
stkowe mięśnie gładkie w ścianach tętnic zaopatrujących mięśnie szkieletowe acetylocholina działa rozkurczająco, noradrenalina zaś kurcząco
za pośrednictwem receptora alpha adrenergicznego.
I Mięsień sercowy
Mięsień sercowy składa się w istocie z dwóch mięśni. Są to: mięsień
przedsionków i mięsień komór, odgrodzone od siebie pierścieniami
włóknistymi {anuli fibrosi) otaczającymi ujście przedsionkowo-komorowe
(ostia atrioventricularis). Mięsień sercowy zbudowany jest z komórek
mięśniowych (myocyti cardiaci), które mają włókienka mięśniowe i podstawowe elementy kurczliwe tworzące sarkomery. Błony komórkowe
sąsiednich komórek mięśniowych ściśle do siebie przylegają w miejscach
występowania prążków Z, tworząc pozazębianą błonkę zwaną wstawką
{discus intercalatus). Dzięki tym wstawkom i połączeniom komunikującym (gap junctions) pobudzenie przenosi się z jednej komórki na drugą
i mięsień sercowy stanowi syncytium fizjologiczne (functional syncytium).
Odpowiada on na bodziec skurczem maksymalnym zgodnie z prawem
„wszystko albo nic".
Potencjał spoczynkowy komórki mięśnia sercowego wynosi 90 mV.
W spoczynku jony potasowe stale wychodzą z komórek mięśnia sercowego — przez tzw. kanały potasowe w błonie komórkowej — na zewnątrz, tworząc odkomórkowy wolny prąd jonów potasowych. Jednocześnie jony sodowe stale wchodzą do komórek, przez tzw. kanały
sodowe, tworząc dokomórkowy wolny prąd jonów sodowych. Przesunięcia w rozmieszczeniu jonów spowodowane wolnym prądem jonów K +
i Na+ są wyrównywane przez pompę sodowo-potasową usuwającą jony
Na+ z komórek i wciągającą jony K+ do wnętrza komórek. Utrzymywanie
się potencjału spoczynkowego jest uwarunkowane działaniem pompy
jonowej w błonie komórkowej komórek mięśnia sercowego.
W czasie pobudzenia komórek mięśnia komór serca występuje czynnościowy potencjał składający się z czterech faz. Rozpoczyna się fazą
0 — bardzo szybko zachodzącą depolaryzacją, po której występuje
nieznaczna repolaryzacja, czyli faza 1, przechodząca w fazę 2, charakteryzującą się utrzymywaniem stałej depolaryzacji w czasie około 300 ms.
W fazie 3 następuje powrót do potencjału spoczynkowego.
Faza 0 potencjału czynnościowego jest wywołana dokomórkowym
szybkim prądem jonów sodowych (fast inward sodium current) i w niewielkim stopniu przez dokomórkowy wolny prąd jonów wapniowych
Mięsień sercowy
Skurcz
komór
Rozkurcz komór
Skurcz
komór
111
Rozkurcz komór
Ryc. 56. Czynności bioelektryczne i mechaniczne mięśnia sercowego. Potencjał wewnątrzkomórkowy komórki węzła zatokowo-przedsionkowego (w.z.p.), komórki mięśnia komór
sercowych (k.s.) — fazy repolaryzacji: 0, 1, 2, i 3, elektrokardiogram (EKG) — załamki P, Q,
R, S, T i czynność mechaniczna komór (c.m.k.).
(slow inward calcium current). Faza 1 w komórkach mięśnia przedsionków wiąże się z odkomorkowym prądem potasowym (outward potassium
current), natomiast w komórkach mięśnia komór jest wywołana przez
dokomórkowy prąd jonów chlorowych.
112
Czynność komórek nerwowych i mięśniowych
W czasie fazy 2 występuje równowaga pomiędzy dokomórkowym
prądem jonów Ca 2+ i odkomórkowym prądem jonów K + . W fazie 0, 1
i 2 jony wapniowe napływają do sarkoplazmy z zewnętrznej powierzchni
sarkolemy, gdzie są magazynowane w glikokaliksie, i tylko częściowo ze
zbiorników sarkoplazmatycznych. W fazie 3 następuje przewaga odkomórkowych prądów jonów dodatnich, głównie wywołana przez jony K +,
i powrót ujemnego potencjału w komórce.
Okres bezwzględnej niewrażliwości, czyli bezwzględnej refrakcji (absolute refractory period), obejmuje depolaryzację oraz fazy 1 i 2 repolaryzacji. Okres względnej niewrażliwości, czyli względnej refrakcji (relative
refractory period), przypada na fazę 3 repolaryzacji. Łącznie oba te
okresy niewrażliwości są dłuższe od czasu skurczu mięśnia sercowego.
Dzięki temu w warunkach prawidłowych w mięśniu tym nie występują
skurcze tężcowe.
Siła skurczu mięśnia sercowego zależy od początkowej długości jego
komórek, podobnie jak to występuje w mięśniach poprzecznie prążkowanych szkieletowych. Przy optymalnym wypełnieniu jam serca i optymalnym rozciągnięciu komórek mięśnia sercowego występują maksymalne skurcze zgodne z tzw. prawem Starlinga.
Energię potrzebną do skurczów mięsień sercowy czerpie ze składników
odżywczych, którymi są: w 60% kwasy tłuszczowe, których połowę
stanowią wolne kwasy tłuszczowe — FFA (free fatty acid), w 35%
węglowodany oraz aminokwasy, ciała ketonowe, kwas mlekowy i kwas
pirogronowy.
Ryc. 57. Wewnątrzkomórkowy potencjał
komórki mięśnia komór serca. 0, 1, 2
i 3 — fazy potencjału czynnościowego,
4 — potencjał spoczynkowy. Prądy jonowe
w czasie występowania potencjału czynnościowego: Na+ —szybki dokomórkowy
prąd jonów sodowych, Ca2+ — wolny
dokomórkowy prąd jonów wapniowych,
K+ — odkomórkowy prąd jonów potasowych.
Mięsień sercowy
113
I Komórki układu przewodzącego serca
Poza komórkami mięśnia przedsionków i mięśnia komór występują
w sercu komórki układu przewodzącego (cardiac conduction system).
Różnią się od pozostałych komórek zarówno pod względem morfologicznym, jak i czynnościowym. W komórkach tych nie ma wyraźnego
poprzecznego prążkowania. Są bogate w glikogen i zawierają więcej
sarkoplazmy. Leżą bezpośrednio pod wsierdziem, tworząc skupienia:
węzeł zatokowo-przedsionkowy, czyli węzeł Keitha-Flacka, węzeł przedsionkowo-komorowy, czyli węzeł Aschoffa-Tawary, i pęczek przedsionkowo-komorowy, czyli pęczek Paladino-Hisa, który dzieli się na dwie
odnogi kończące się komórkami mięśniowymi sercowymi przewodzącymi (włóknami Purkinjego).
Komórki układu przewodzącego samoistnie i rytmicznie pobudzają
się. Tworzą tym samym rozrusznik (pacemaker) dla całego mięśnia
sercowego. Komórki węzła zatokowo-przedsionkowego pobudzają się
samoistnie najszybciej w rytmie około 1,2 Hz, tj. 72 razy na minutę.
Pozostałe komórki układu przewodzącego pobudzają się samoistnie
w rytmie wolniejszym. W związku z tym komórki węzła zatokowo-przedsionkowego narzucają swój rytm wszystkim pozostałym komórkom
układu przewodzącego i komórkom całego mięśnia sercowego.
Samoistne pobudzenie komórek węzła zatokowo-przedsionkowego
jest wywołane powolną depolaryzacją błony komórkowej występującą
pomiędzy potencjałami czynnościowymi. Jest to prepotencjał rozrusznika
(pacemaker potential). Występuje na skutek nasilania się dokomórkowego prądu jonów wapniowych aż do osiągnięcia potencjału progowego,
przy którym wyzwala się potencjał czynnościowy. Potencjał czynnościowy, czyli depolaryzacja, powstający w węźle zatokowo-przedsionkowym rozchodzi się na mięsień przedsionków, a następ nie na węzeł
i pęczek przedsionkowo-komorowy, komórki mięśniowe serca przewodzące i mięsień komór (patrz Czynność bioelektryczna serca, str. 336).
Skurcze mięśnia przedsionków i mięśnia komór są wywołane wyłącznie
stanem czynnym powstającym w komórkach układu przewodzącego.
Komórki nerwowe znajdujące się w samym sercu lub wysyłające swe
aksony do serca nie uczestniczą w wyzwalaniu pobudzenia w sercu.
Z aksonów neuronów należących do układu współczulnego i przywspółczulnego są uwalniane transmittery, które mają tylko działanie modelujące czynność serca — przyspieszają lub zwalniają jego skurcze.
Acetylocholina uwolniona z zakończeń neuronów przywspółczulnych, działając poprzez receptor muskarynowy (M2), zwiększa wychodzę-
114
Czynność komórek nerwowych i mięśniowych
Ryc. 58. Częstotliwość występowania potencjału wewnątrzkomórkowego komórki węzła
zatokowo-przedsionkowego: a — pod wpływem acetylocholiny uwalnianej z zakończeń
neuronów przywspółczulnych; b — bez wpływu transmitterów układu autonomicznego;
c — pod wpływem noradrenaliny uwalnianej z zakończeń neuronów wspołczulnych.
nie jonów K+ z komórek rozrusznika i wydłuża czas trwania prepotencjału rozrusznika. Stan czynny w komórkach układu przewodzącego
występuje rzadziej i serce kurczy się w zwolnionym rytmie. Noradrenalina wydzielona z zakończeń neuronów wspołczulnych, wiążąc się z receptorami beta-adrenergicznymi, których jest znacznie więcej w sercu niż
receptorów alpha-adrenergicznych, zwiększa napływ jonów Ca 2+ do
komórek rozrusznika. Czas trwania prepotencjału rozrusznika skraca się,
stan czynny w jego komórkach występuje częściej i przyspiesza się częstość skurczów serca.
U człowieka pozostającego w spoczynku zablokowanie farmakologiczne receptorów cholinergicznych wywołuje przyspieszenie częstości
skurczów serca. Przeciwny stan występuje po zablokowaniu receptorów
adrenergicznych — częstość skurczów serca ulega zwolnieniu. Po farmakologicznym zablokowaniu obu receptorów, cholinergicznych i adrenergicznych, częstość skurczów serca przyspiesza się w porównaniu do
częstości występującej w spoczynku, ponieważ przeważa oddziaływanie
na serce układu przywspółczulnego w stosunku do układu wspołczulnego.
Czucie i ruch
Czynność ośrodkowego układu nerwowego człowieka ściśle wiąże się
z trzema podstawowymi procesami, którymi są:
• odbieranie bodźców (reception) ze środowiska zewnętrznego i wewnę
trznego organizmu;
• adekwatne reagowanie (adeąuate response) na odbierane bodźce;
• przechowywanie śladów po bodźcach, czyli ich zapamiętywanie (memorization).
Za pośrednictwem aferentnych włókien nerwowych impulsy nerwowe
są przewodzone od receptorów (receptors) do niższych pięter ośrodkowego
układu nerwowego. Następnie poprzez coraz to wyższe piętra impulsy
nerwowe są przekazywane do najwyższego piętra — do kory mózgu.
W kierunku przeciwnym — od kory mózgu do efektorów (effectors) —
impulsy nerwowe przekazywane są przez coraz to niższe piętra i eferentne
włókna nerwowe, biegnące w nerwach czaszkowych i rdzeniowych.
Pomiędzy ośrodkami znajdującymi się na różnych piętrach układu
nerwowego impulsy przewodzone są jednocześnie za pośrednictwem
połączeń o małej i dużej liczbie neuronów. Impulsy przekazywane są za
pośrednictwem małej liczby neuronów w obrębie układów swoistych
(specific systems) — szybko przewodzących aferentnych: układu wstęg
i dróg rdzeniowo-wzgórzowych, oraz szybko przewodzących eferentnych:
dróg korowo-rdzeniowych. Duża liczba neuronów uczestniczy w przekazywaniu impulsów w układach nieswoistych (nonspecific systems) — wolno
przewodzących.
Układy swoiste są filogenetycznie młodsze. Potrzeba szybkiego reagowania na bodźce z otoczenia doprowadziła do wykształcenia się w ośrodkowym układzie nerwowym dróg, ośrodków i całych układów szybko
przekazujących impulsy od receptorów do efektorów za pośrednictwem
116
Czucie i ruch
ściśle określonych ośrodków swoistych dla miejsca i charakteru działającego bodźca.
W obrębie układów nieswoistych, wolno przewodzących impulsy,
znajdują się skupienia neuronów o wyspecjalizowanej funkcji. Skupienia
tych neuronów tworzą ośrodki kontrolujące czynność poszczególnych
narządów i układów, jak np. układu sercowo-naczyniowego, układu
oddechowego, układu trawiennego i innych. Ośrodki kontrolujące zachowanie człowieka, przystosowujące jego zachowanie do bodźców z otoczenia, jak i do bodźców ze środowiska wewnętrznego, znajdują się
również w układach nieswoistych. W tych układach występują komórki
nerwowe stanowiące rozruszniki dla spontanicznej czynności bioelektrycznej mózgu.
I
Czynność bioelektryczna mózgu
spontaniczna
Czynność bioelektryczną mózgu bada się za pomocą elektrod odbierających (exploring electrodes) przystawianych do skóry głowy. Zarejestrowane zmiany potencjału elektrycznego mózgu przyjęto nazywać
elektroencefalogramem (EEG). Elektrody można również przystawić bezpośrednio do odsłoniętej w czasie operacji kory mózgu i zarejestrować
potencjały elektryczne nazywane elektrokortykogramem (ECoG).
W zależności od miejsca, z którego odbiera się potencjały bioelektryczne, mają one różną amplitudę i występują z różną częstotliwością.
Każdy człowiek ma indywidualny zapis potencjałów elektrycznych
mózgu, choć jednocześnie zapisy z poszczególnych okolic różnią się.
Potencjały elektryczne zarejestrowane na taśmie papieru mają postać fal
o różnej amplitudzie i częstotliwości. Wyróżnia się fale EEG: alpha, beta,
theta i delta (alpha, beta, theta i delta waves). Fale alpha występują
przeważnie w okolicy ciemieniowo-potylicznej z częstotliwością od 8 do
13 Hz o amplitudzie do 50 uV, fale beta — w okolicy czołow ej
z częstotliwością od 14 do 60 Hz o amplitudzie mniejszej — do 30 \JM.
Amplituda fal beta jest zawsze niższa w porównaniu z amplitudą fal
alpha. Fale alpha i beta są to zasadnicze rodzaje fal rejestrowane
u człowieka czuwającego, spoczywającego, siedzącego lub leżącego nieruchomo z zamkniętymi oczami.
Elektroencefalogram jest sumą potencjałów wytwarzanych przez pobudzone ciała neuronów i ich wypustki. Im więcej neuronów jest pobudza-
Czynność bioelektryczna mózgu spontaniczna
17
nych rytmicznie w krótkich odstępach czasu, tym wypadkowa tych
potencjałów jest większa. Określa się ten stan jako synchronizację czynności elektrycznej mózgu (EEG synchronization). Niejednoczesne pobudzenie neuronów powoduje, że poszczególne potencjały komórkowe,
przede wszystkim potencjały dendrytyczne, nie sumują się wzajemnie.
Rejestruje się wtedy potencjały o niskiej amplitudzie i dużej częstotliwości,
czyli niesynchroniczną aktywność neuronów — desynchronizację.
W celu uzyskania zapisu EEG w badaniach rutynowych, zgodnie
z przyjętym przez Międzynarodową Federację EEG układem elektrod
10-20 (ten-twenty system), na powierzchni skóry na głowie w miejscach
ściśle określonych umieszcza się 19 elektrod odbierających. Nad każdą
półkulą mózgu znajduje się po 8 elektrod, 3 elektrody zaś przystawione
są do skóry w linii środkowej. Nad korą mózgu płatów czołowych zostaje
umieszczonych 7 elektrod (Fp15 Fp2, F3, F4, F7, F8 i Fz), na pograniczu
płatów czołowych i ciemieniowych — 3 elektrody (C 3, C 4 i C 2), nad
płatami ciemieniowymi — 3 elektrody (P 3, P 4 i P z), nad płatami skro-
Ryc. 59. Elektroencefalogram człowieka odebrany jednocześnie przy zastosowaniu szesnastu
dwubiegunowych odprowadzeń (I—XVI). Odbiór czynności bioelektrycznych przeprowadzono, stosując tzw. podłużny montaż dwubiegunowy (longitudinal bipolar montage).
Poszczególne odprowadzenia uzyskano z par aktywnych elektrod położonych względem
siebie w linii strzałkowej.
118
Czucie i ruch
niowymi — 4 elektrody (T 3, T4, T5, i T6) i nad płatami potylicznymi — 2
elektrody (Oj i O2). Odbierając wahania potencjałów elektrycznych
występujące pod sąsiadującymi ze sobą elektrodami, uzyskuje się tzw.
odprowadzenia dwubiegunowe (bipolar derivation) i można jednocześnie
zarejestrować EEG z 16 odprowadzeń (I—XVI). Stosując umowne
układy odbioru EEG z określonych par elektrod, według norm przyjętych przez Międzynarodową Federację EEG, można zarejestrować czynność bioelektryczną znacznych obszarów kory mózgu obu półkul.
Po otwarciu oczu, z czym związany jest odbiór wrażeń wzrokowych,
czynność bioelektryczna w okolicy potylicznej kory mózgu ulega desynchronizacji. W EEG rejestrowanym z odprowadzeń okolicy potylicznej
fale alpha zanikają, co nazywane jest również tłumieniem fal alpha (alpha
wave blocking) (patrz ryc. 106, str. 198).
Elektrogeneza potencjałów korowych
Potencjały elektryczne odbierane z powierzchni kory mózgu powstają
na skutek przesuwania się wzdłuż dendrytów depolaryzacji ich błony
komórkowej. Depolaryzacja dendrytów, przesuwając się od powierzchni
kory mózgu w kierunku ciał neuronów, kolejno zmienia potencjał elektryczny odbierany z powierzchni kory. Wolno przesuwający się stan
czynny wzdłuż dendrytów powoduje przesuwanie się ujemnego potencjału
z powierzchni w głąb kory mózgu i powierzchnia kory na ułamek sekundy
ma dodatni ładunek.
Samoistna czynność bioelektyczna mózgu związana jest ze stałym
krążeniem impulsów pomiędzy neuronami należącymi do różnych struktur mózgowia tworzących mechanizmy desynchronizujące i mechanizmy
synchronizujące. Neurony, pomiędzy którymi krążą impulsy, znajdują się
w korze mózgu, we wzgórzu, w jądrze ogoniastym, w hipokampie
i w innych strukturach podkorowych. Impulsacja z układu siatkowatego
wstępującego pobudzającego — RAS (reticular activating system) — zakłóca synchroniczne krążenie impulsów po zamkniętych łańcuchach
neuronów i wywołuje desynchronizację (patrz Układ siatkowaty wstępujący, str. 191).
Czynność bioelektryczna wywołana
Pobudzając receptory uzyskuje się salwę impulsów aferentnych przewodzonych przez szereg neuronów czuciowych do kory mózgu. Salwa
Czynność bioelektryczna mózgu spontaniczna
119
impulsów wyzwala w niej wahania potencjału elektrycznego. Wywołane
potencjały elektryczne nakładają się na spontaniczną czynność bioelektryczną kory mózgu i w czasie rutynowego badania EEG trudno jest
te dwie odmienne czynności bioelektryczne odróżnić od siebie. Wielokrotnie powtarzając ten sam bodziec działający na te same receptory,
można za pomocą komputera uśrednić kilkanaście czy kilkaset potencjałów wywołanych. Spontaniczne wahania potencjałów kory mózgu
w czasie uśredniania wzajemnie się znoszą. Przeciwnie — potencjały
wywołane, rejestrowane i uśredniane synchronicznie z działającym bodźcem, sumują się ze sobą.
Działając na siatkówkę błyskami lampy stroboskopowej, kiedy powieki są zamknięte, uzyskuje się w okolicy potylicznej kory mózgu
wzrokowy potencja] wywołany — VEP (visual evoked potential). Podobnie
Ryc. 60. Badanie wzrokowych potencjałów wywołanych. Fotostymulator jednocześnie
pobudza siatkówkę i wyzwala komputer. Komputer uśrednia wzrokowe potencjały wywołane
(VEP), które z okolicy potylicznej odebrał i wzmocnił elektroencefalograf (EEG).
1 20
Czucie i ruch
powtarzane bodźce słuchowe pozwalają zarejestrować z okolicy skroniowej kory mózgu słuchowy potencjał wywołany — AEP (auditory
evoked potential), bodźce czuciowe zaś — czuciowy potencjał wywołany —
SSEP (somatosensory evoked potential).
Potencjał wywołany charakteryzuje się szeregiem załamków ujemnych i dodatnich. Załamki te występują w okresie od kilku do kilkuset
milisekund od początku zadziałania bodźca. Kształt potencjału wywołanego jest indywidualny dla każdego człowieka, choć szereg załamków
zarówno dodatnich, jak i ujemnych uzyskuje się u wszystkich ludzi.
Amplituda załamków i ich okres utajonego pobudzenia zależą od wielu
czynników, a przede wszystkim od częstotliwości, siły i liczby bodźców.
0
100
200
300
400
SOOms
Ryc. 61. Wzrokowy potencjał wywołany (VEP) zawierający składowe wczesne (20—80 ms),
pośrednie (80—140 ms) i późne (140—450 ms). Składowe potencjały oznaczono literą P —
dodatnie potencjały elektryczne, lub literą N — ujemne potencjały elektryczne, i okresem
utajonego pobudzenia w milisekundach. Literami P lub N i okresem utajonego pobudzenia
oznaczono tylko składowe potencjały elektryczne pochodzenia mózgowego. Składowe
potencjały nie oznaczone są to artefakty towarzyszące wzrokowym potencjałom wywołanym.
Pochodzą one z siatkówki (ERG) i z mięśni głowy (EMG). (Do wykreślenia krzywej VEP
wykorzystano wyniki badań opublikowane w: T. Allison, Y. Matsumiya, G. D. Goff,
W. R. Goff The scalp topography of human visual evoked potentials. Electroenceph. Clin.
Neurophysiol., 1977, 45, 185—197).
Odruchy
121
Na podstawie różnicy okresów utajonego pobudzenia załamków czuciowych potencjałów (SSEP) wywołanych drażnieniem przez skórę w okolicy kostek nerwu piszczelowego i w okolicy nadgarstka nerwu pośrodkowego oznacza się prędkość przewodzenia impulsów w drogach czuciowych w rdzeniu kręgowym. Z różnicy w okresie utajonego pobudzenia
załamków SSEP, odbieranego z rdzenia kręgowego w odcinku szyjnym
(C3—C4) i z pola czuciowego w korze mózgu, dokonuje się pomiaru
czasu ośrodkowego przewodzenia (central conduction time), który u dorosłych ludzi wynosi 5,6 ±0,5 ms.
I Odruchy
Odbieranie bodźców i adekwatne reagowanie na nie, co nosi nazwę
czynności odruchowej, jest najbardziej istotnym i podstawowym przejawem funkcji ośrodkowego układu nerwowego. Odruch (reflex) jest to
odpowiedź efektora wywołana przez bodziec działający na receptor
i wyzwolona za pośrednictwem układu nerwowego. Droga jaką przebywa
impuls nerwowy od receptora do efektora, nazywa się łukiem odruchowym
(reflex arc).
W rozwoju filogenetycznym wykształciły się drogi dla impulsów
nerwowych łączące określone receptory z określonymi efektorami. Są
to połączenia wrodzone, które w miarę wzrostu i dojrzewania organizmu, a wraz z nim ośrodkowego układu nerwowego, zaczynają przewodzić impulsy nerwowe. Dzięki tym wrodzonym połączeniom bodziec
działający na określony receptor wyzwala odpowiedź określonego efektora. Odpowiedź na bodziec jest odruchem wrodzonym zwanym również
odruchem bezwarunkowym — UR (unconditioned reflex).
W rozwoju ontogenetycznym wytwarzają się również nowe połączenia
pomiędzy różnymi ośrodkami. Dzięki temu pojawiają się nowe odruchy,
które są odruchami nabytymi, czyli odruchami warunkowymi — CR
(conditioned reflexes). Wytworzenie się nowego połączenia pomiędzy
ośrodkami wymaga pewnych określonych warunków. Występowanie
odruchu nabytego jest uwarunkowane działaniem zespołu bodźców.
Odruchy nabyte charakteryzują się dużą zmiennością odpowiedzi na
bodźce, w przeciwieństwie do odruchów wrodzonych, które dają zawsze
tę samą odpowiedź na ten sam bodziec.
1 22
Czucie i ruch
I Łuk odruchowy
Droga jaką przebywa impuls nerwowy od receptora do efektora,
stanowi łuk odruchowy.
Łuk odruchowy składa się z pięciu zasadniczych części:
•
•
•
•
•
receptora;
aferentnego włókna nerwowego (dośrodkowego);
ośrodka nerwowego;
eferentnego włókna nerwowego (odśrodkowego);
efektora.
W zależności od liczby neuronów w ośrodkach nerwowych, przewodzących impuls nerwowy od receptora do efektora, odruchy dzielą się
na proste i złożone. Odruchy proste są to przede wszystkim odruchy
rdzeniowe. Łuki odruchowe tych odruchów składają się z dwóch lub
trzech komórek nerwowych. Impulsy nerwowe do i od rdzenia kręgowego
biegną przez nerwy rdzeniowe, które składają się zarówno z włókien
aferentnych, jak i eferentnych. W miejscu rozdzielenia się nerwu rdzeniowego na korzeń grzbietowy (radix dorsalis) i na korzeń brzuszny
{radix ventralis) rozdzielają się również włókna aferentne od eferentnych.
Włókna aferentne wstępują do rdzenia kręgowego w korzeniach grzbietowych, a eferentne opuszczają rdzeń przez korzenie brzuszne.
Korzeń grzbietowy na swoim przebiegu ma zgrubienie stanowiące zwój
rdzeniowy (ganglion spinale). W zwoju rdzeniowym znajdują się ciała
neuronów czuciowych odbierających pobudzenie z receptorów. Neurony
czuciowe wysyłają wypustkę kończącą się synapsami w istocie szarej
rdzenia kręgowego. Synapsy neuronu czuciowego otaczają bezpośrednio
Odruchy
123
neuron ruchowy w rogach przednich rdzenia i w takim łuku odruchowym
występuje tylko jedna synapsa pomiędzy neuronem czuciowym i ruchowym. Jest to łuk odruchowy odruchu monosynaptycznego (monosynaptic
reflex arc).
Część neuronów czuciowych ma wypustki, które otaczają synapsami
neurony pośredniczące znajdujące się w istocie szarej rdzenia kręgowego.
Z kolei aksony neuronów pośredniczących kończą się synapsami na
neuronach ruchowych w rogach przednich. W takim łuku odruchowym
występują trzy neurony i dwie synapsy pomiędzy nimi. Jest to łuk
odruchowy odruchu polisynaptycznego (polysynaptic reflex arc). W łukach
odruchowych może występować więcej neuronów niż dwa lub trzy.
Istnieją odruchy, w których uczestniczą dziesiątki i setki neuronów.
Odruch monosynaptyczny
W organizmie człowieka występuje tylko jeden rodzaj odruchów
monosynaptycznych. Jest to odruch na rozciąganie (stretch reflex). Odruch
ten zostaje wywołany przez rozciągnięcie mięśnia szkieletowego. Na
skutek wydłużenia komórek intrafuzalnych we wrzecionkach nerwo-womięśniowych dochodzi do pobudzenia zakończenia pierścieniowa-tospiralnego. Jest to zakończenie neuronu czuciowego, którego ciało
znajduje się w zwoju rdzeniowym. Na skutek rozciągania zakończenie
pierścienowato-spiralne depolaryzuje się i salwy impulsów biegną do
rdzenia kręgowego. Tam po przejściu przez jedną synapsę pobudzają
neurony ruchowe, które z kolei wysyłają impulsy wywołujące skurcz
izometryczny lub izotoniczny mięśnia szkieletowego (patrz str. 102).
Odruch polisynaptyczny
Bodziec działając na receptory, np. znajdujące się w skórze, pobudza je,
czyli depolaryzuje. Pobudzenie receptorów jest odbierane przez otaczające je włókna nerwowe — wypustki neuronów czuciowych ze zwojów
rdzeniowych. Impulsy nerwowe przewodzone są przez wypustki neuronów czuciowych od receptorów do istoty szarej rdzenia kręgowego.
Tam są odbierane przez neurony pośredniczące, które z kolei pobudzają
neurony ruchowe. W wyniku działania na kończynę silnego bodźca
uszkadzającego tkanki dochodzi do skurczu mięśni zginaczy i do odruchowego zgięcia kończyny. Odruch zginania (withdrawal reflex) kończyn
jest stosunkowo prostym odruchem polisynaptycznym.
1 24
Czucie i ruch
Ryc. 63. Łuk odruchowy somatyczny: monosynaptyczny (a) i polisynaptyczny (b) oraz
łuk odruchowy autonomiczny (c).
I Ośrodki rdzenia kręgowego
Ośrodki nerwowe są to skupienia neuronów w ośrodkowym układzie
nerwowym zawiadujące określoną funkcją. Od czynności ośrodka zależy,
czy odruch wystąpi, jaki będzie jego okres utajonego pobudzenia, czyli
Odruchy
125
latencja (latent period), i z jaką siłą zostanie pobudzony efektor. Przewodzenie impulsów przez ośrodki nerwowe różni się znacznie od przewodzenia impulsów w nerwach obwodowych. Przewodzenie przez ośrodki
polega na przenoszeniu się impulsów z jednych neuronów na drugie,
w czym pośredniczą synapsy (patrz str. 71).
Decydującym elementem dla odruchów rdzeniowych jest czynność
neuronów w jądrach ruchowych (motor nuclei) w rogach przednich
rdzenia kręgowego. Jądra te skupiają neurony ruchowe unerwiające
poszczególne mięśnie szkieletowe. Neurony ruchowe odbierają i przewodzą różnego rodzaju impulsację. Z tego względu zostały nazwane
„końcową wspólną drogą" („finał common path").
Neurony ruchowe odbierają impulsy nerwowe biegnące od:
• neuronów czuciowych zwojów rdzeniowych;
• neuronów pośredniczących w istocie szarej rdzenia kręgowego;
• neuronów wyższych pięter ośrodkowego układu nerwowego.
Impulsy od neuronów czuciowych w zwojach rdzeniowych do neuronów ruchowych przekazywane są bezpośrednio przez synapsy pobudzające. Natomiast impulsacja neuronów pośredniczących i neuronów wyższych pięter może być przekazywana zarówno przez synapsy pobudzające,
jak i hamujące. W warunkach fizjologicznych impulsacja wysyłana przez
neurony ruchowe do mięśni szkieletowych jest wypadkową działania
synaps pobudzających i hamujących.
W obrębie błony komórkowej neuronów ruchowych, odbierających
liczne impulsy od synaps wokół nich rozmieszczonych, zachodzi zjawisko
sumowania przestrzennego i sumowania w czasie impulsów nerwowych
(patrz str. 67).
Pobudliwość ośrodków rdzenia kręgowego
Jednoczesne słabe podrażnienie włókien aferentnych w dwóch nerwach
może wywołać znacznie silniejszą reakcję efektora, np. skurcz mięśnia
szkieletowego, niż suma reakcji wywołana oddzielnym podrażnieniem,
bodźcem o tej samej sile tych samych nerwów. Silniejsza reakcja efektora
wywołana jednoczesnym podrażnieniem dwóch nerwów jest to zjawisko
torowania (facilitation) zachodzące w ośrodkach nerwowych rdzenia
kręgowego. Wokół neuronów pobudzonych podrażnieniem poszczególnych nerwów znajdują się neurony w strefie zwiększonej podprogowo
pobudliwości (subliminal fringe zone). Neurony zachodzących na siebie
1 26
Czucie i ruch
stref zwiększonej podprogowo pobudliwości konwergują impulsację przewodzoną przez dwa nerwy i wysyłają impulsację do efektora.
Zjawiskiem przeciwnym do torowania jest zjawisko okluzji (occlusion).
Jednoczesne maksymalne podrażnienie dwóch nerwów daje słabszą reakcję efektora niż suma reakcji wywołana oddzielnym podrażnieniem tych
samych nerwów. Strefy pobudzenia (discharge zone) obu ośrodków
zachodzą na siebie. Tym samym jednoczesne maksymalne podrażnienie
dwóch nerwów pobudza mniejszą liczbę neuronów w porównaniu z sumą
neuronów pobudzanych oddzielnie.
Pomiędzy neuronami ośrodków przewodzone są impulsy wzmacniane
przez zwielokrotnione łańcuchy neuronów i krążące po zamkniętych
łańcuchach neuronów. Krążenie i sumowanie impulsów w ośrodkach
rdzenia kręgowego wywołuje stałe wahania postsynaptycznego potencjału pobudzającego (EPSP) w neuronach ruchowych. Wysyłane przez te
neurony eferentne impulsy nerwowe mają w związku z tym większą lub
mniejszą częstotliwość i unerwione przez nie mięśnie szkieletowe silniej
lub słabiej kurczą się.
Ryc. 64. Dwa typy dróg przekazywania impulsacji pomiędzy neuronami: a — zwielokrotniony łańcuch neuronów; b — zamknięty łańcuch neuronów.
Opóźnienie synaptyczne
Impulsy nerwowe są przewodzone przez włókna nerwowe ze stałą
prędkością. Prędkość przewodzenia impulsów znacznie zwalnia się w ośrodkach rdzenia kręgowego. Zwolnienie ośrodkowe zależy od liczby
synaps, przez które są przenoszone impulsy. Przejście impulsu w rdzeniu
kręgowym przez jedną synapsę trwa około 0,8 ms. Czas trwania opóźnienia synaptycznego (synaptic delay) podlega wahaniom — może ulec
skróceniu (torowanie) do 0,5 ms lub wydłużeniu (tłumienie) do 0,9 ms
w zależności od stanu czynnościowego ośrodka ruchowego rdzenia
kręgowego.
Odruchy
127
Odruchy własne rdzenia kręgowego
Odruch monosynaptyczny na rozciąganie mięśnia i odruch polisynaptyczny zginania kończyny występują również po przerwaniu połączeń
rdzenia kręgowego z wyższymi piętrami ośrodkowego układu nerwowego.
Dlatego nazywa się je odruchami własnymi rdzenia kręgowego. Wystąpienie każdego z tych odruchów wiąże się z pobudzeniem ośrodka
ruchowego dla mięśni kurczących się, znajdującego się w rogach przednich
rdzenia kręgowego.
Wzajemnie zwrotne stosunki pomiędzy ośrodkami
Wzajemnie zwrotne unerwienie mięśni szkieletowych (reciprocal innervation) i wzajemnie zwrotne stosunki pomiędzy ośrodkami dla tych mięśni
polegają na tym, że pobudzenie jednych ośrodków wywołuje jednoczesne
hamowanie innych ośrodków.
Neurony ruchowe w rogach przednich rdzenia kręgowego unerwiają
wszystkie mięśnie poprzecznie prążkowane tułowia i kończyn. Neurony
dla poszczególnych mięśni i grup mięśni tworzą ośrodki ruchowe. W czasie
pobudzenia ośrodka ruchowego, np. dla mięśni zginaczy kończyny
górnej, ośrodki ruchowe dla mięśni antagonistycznych — prostowników — są zahamowane, a mięśnie rozkurczone.
W ośrodku ruchowym zostają pobudzone jednocześnie neurony ruchowe alpha, unerwiające komórki mięśniowe ekstrafuzalne, i neurony ruchowe gamma, unerwiające komórki mięśniowe intrafuzalne, w których
znajdują się zakończenia pierścieniowato-spiralne (terminatio nervi anulospiralis), czyli A-receptory. Skurcz komórek mięśniowych zwiększa pobudliwość zakończeń pierścieniowato-spiralnych. Aferentne impulsy nerwowe, przewodzone od tych zakończeń przez neurony czuciowe, są
przekazywane do neuronów ruchowych alpha przez połączenia monosynaptyczne. Jednocześnie za pośrednictwem gałęzi obocznych neurony
czuciowe wysyłają impulsy do ośrodków ruchowych dla mięśni antagonistycznych. Gałęzie oboczne kończą się w tych ośrodkach synapsami na
neuronach pośredniczących, hamujących. Neurony te wywołują postsynaptyczny potencjał hamujący (IPSP) w neuronach ruchowych dla
mięśni antagonistycznych. Dzięki tym wzajemnie zwrotnym stosunkom
pomiędzy ośrodkami dla mięśni antagonistycznych nie dochodzi w warunkach fizjologicznych do ich jednoczesnego pobudzenia.
Pobudliwość zakończeń pierścieniowato-spiralnych wzrasta nie tylko
w czasie skurczu mięśnia, lecz również w czasie jego biernego roz -
1 28
Czucie i ruch
ciągnięcia. Pod wpływem salwy aferentnej impułsacji zostają pobudzone j
neurony ruchowe alpha i mięsień kurczy się. Rozciągnięcie mięśnia
wywołuje jego krótkotrwały skurcz, jak to występuje w czasie badania
odruchów na rozciąganie, np. odruchu kolanowego lub o druchu ze
ścięgna piętowego (Achillesa).
Długotrwałe rozciąganie mięśnia powoduje stałą wzmożoną impulsację
aferentną wysyłaną przez zakończenia pierscieniowato-spiralne i mięsień
w wyniku odruchu monosynaptycznego stale się kurczy. Ośrodki dla
mięśni antagonistycznych zostają w tym czasie zahamowane.
PP-IA
Ryc. 65. Stosunki wzajemnie zwrotne pomiędzy ośrodkami
dla mięśni antagonistycznych goleni: prostownika —mięśnia
piszczelowego przedniego, i zginacza — mięśnia brzuchatego
łydki. Wrzecionka nerwowo-mięśniowe (A) rozciągniętego
mięśnia piszczelowego przedniego (PP) wysyłają impulsację
za pośrednictwem włókien nerwowych IA (PP-IA) do motoneuronów unerwiających ten mięsień (segment L5 w rdzeniu
kręgowym) i jednocześnie do neuronów pośredniczących
(segment Sl w rdzeniu kręgowym) hamujących motoneurony
mięśnia zginacza brzuchatego łydki. S — zakończenia synaptyczne nerwowo-mięśniowe.
Czucie
129
Bardzo silny skurcz mięśnia lub bardzo silne jego rozciągnięcie powoduje pobudzenie ciałek buławkowatych. czyli B-receptorów znajdujących się
w ścięgnach. Ciałko buławkowate {corpusculum bulboideum) ma około
100 razy wyższy próg pobudliwości niż zakończenie pierścieni owato-spiralne. Impulsy wysyłane przez B-receptory hamują neurony ośrodka dla
mięśnia kurczącego się i pobudzają neurony ośrodka dla mięśnia antagonisty cznego.
Hamowanie ośrodka dla mięśni nadmiernie kurczących się lub zbyt
silnie rozciągniętych odbywa się na zasadzie hamowania auto gennego (autogenic inhibition). Dzięki niemu nie dochodzi do uszkodzenia mięśnia przez rozerwanie jego komórek.
Wstrząs rdzeniowy
Po całkowitym przerwaniu ciągłości rdzenia kręgowego, po jego
zmiażdżeniu lub gwałtownym uciśnięciu, występuje u ludzi tzw. wstrząs
rdzeniowy (spinał shock), który objawia się zniesieniem wszystkich
odruchów rdzeniowych poniżej miejsca uszkodzenia. Nie występują zarówno odruchy na rozciąganie (mięśnie są wiotkie), jak i odruch zginania. Po pewnym czasie pierwszy powraca odruch zginania, ale w postaci odruchu patologicznego. U ludzi zdrowych podrażnienie skóry na
podeszwie wywołuje zgięcie podeszwowe palców, zgięcie grzbietowe stopy
i cofnięcie kończyny. Podrażnienie skóry na podeszwie po uszkodzeniu
rdzenia kręgowego wywołuje odruch patologiczny zwany odruchem
Babińskiego. Polega on na zgięciu podeszwowym czterech palców stopy
i jednoczesnym zgięciu grzbietowym palucha.
Po kilku tygodniach po uszkodzeniu rdzenia kręgowego odruchy,
początkowo bardzo słabe, stopniowo nasilają się. Dochodzi do wzmożenia odruchów (hyperreflexia), a po kilku miesiącach występuje również
tzw. odruch masowy (mass reflex). Podrażnienie skóry na podeszwie
wywołuje zgięcie palców i stopy, cofnięcie obu kończyn i jednocześnie
odruchowe oddawanie moczu i kału.
I Czucie
Pobudzenie receptorów wywołuje nie tylko przewodzenie impulsów
najkrótszą drogą przez łuk odruchowy do efektorów, lecz również
przewodzenie do ośrodków, w których skupione są neurony czuciowe
5 — Fizjologia człowieka
1 30
Czucie i ruch
wyższego rzędu. Pobudzenie neuronów w ośrodkach czuciowych jest
podstawą czucia (sensation). Pobudzenie jednego rodzaju receptorów
wywołuje czucie bodźca. Czucie jest to proste wrażenie zmysłowe polegające na subiektywnej ocenie bodźców pobudzających odpowiednie
receptory i na jednoczesnym przewodzeniu impulsacji czuciowej przez |
drogi czuciowe swoiste i nieswoiste.
Percepcja
Dzięki jednoczesnemu oddziaływaniu na kilka rodzajów receptorów
możliwe jest lepsze rozpoznanie bodźca. Pojęciem wyższego rzędu w stosunku do czucia jest pojęcie percepcji (perception). Obejmuje ona jednocześnie kilka rodzajów czucia i dzięki temu istnieje większa możliwość
rozpoznania zarówno samych bodźców, jak i ich źródła. Podstawą
percepcji jest więc złożone wrażenie zmysłowe.
W warunkach fizjologicznych wrażenia zmysłowe powstają tylko
wtedy, kiedy impulsacja wysyłana przez receptory jest jednocześnie
przewodzona przez neurony należące do dróg czuciowych swoistych
(specific sensory pathways) i dróg czuciowych nieswoistych (nonspecific
sensory pathways). Jednoczesne przewodzenie impulsacji czuciowej przez
drogi swoiste i nieswoiste jest warunkiem odbierania wrażeń zmysłowych.
Drogi swoiste przewodzą impulsy w czasie snu fizjologicznego i w czasie narkozy chirurgicznej. Przewodzenie przez drogi nieswoiste jest w tym
czasie zahamowane i wrażenia zmysłowe nie są odbierane. Przeciwnie,
po uszkodzeniu dróg swoistych i zachowaniu nie naruszonych dróg
nieswoistych wrażenia zmysłowe są odbierane, ale nie są rozpoznawane.
Podział czucia
W zależności od receptorów wyspecjalizowanych w odbieraniu określonych bodźców czucie dzieli się na:
•
•
•
•
teleceptywne;
eksteroceptywne;
prioprioceptywne;
interoceptywne.
Czucie teleceptywne
1 31
I Czucie teleceptywne
Czucie teleceptywne jest odbierane przez narządy zmysłów, a więc
przez narząd powonienia, wzroku i słuchu. Przez telereceptory są odbierane bodźce działające na organizm z pewnej odległości.
I Zmysł powonienia
Cząsteczki związków chemicznych wprowadzone do jamy nosowej
wraz z powietrzem wdychanym rozpuszczają się w śluzie pokrywającym
nabłonek błony śluzowej okolicy węchowej. Tą drogą działają na wypustki komórek nerwowo-zmysłowych węchowych w kształcie rzęsek.
Komórka nerwowo-zmysłowa węchowa (I neuron czuciowy) jednocześnie
odbiera bodźce i przewodzi impulsy nerwowe.
Wypustki około dwudziestu komórek nerwowo-zmysłowych węchowych tworzą włókna nerwowe węchowe, które po wejściu do dołu
przedniego czaszki kończą się synapsami w opuszce węchowej (bulbus
olfactorius) na komórkach nerwowych mitralnych (II neuron czuciowy).
Z opuszki węchowej aksony idą w trzech kierunkach przez:
• prążek węchowy przyśrodkowy {stria olfactoria medialis) i spoidło
przednie (commissura anterior) biegną do opuszki węchowej po stronie
przeciwnej;
Ryc. 66. Komórki nerwowo-zmysłowe, węchowe (I neuron czuciowy), w okolicy
węchowej jamy nosowej odbierające bodźce węchowe. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd.
Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
1 32
Czucie i ruch
m prążek węchowy pośredni (stria olfactoria intermedia) aksony neuronów opuszki osiągają istotę dziurkowaną przednią (substantia perforata anterior), skąd biegną do podwzgórza i innych struktur układu
rąbkowego (limbie system).
• prążek węchowy boczny (stria olfactoria lateralis), który zawiera
aksony kierujące się do III neuronu czuciowego w części korowej
i przyśrodkowej ciała migdałowatego (corpus amygdaloideum) oraz do
haka zakrętu hipokampa (uncus gyri parahippocampalis).
Próg pobudzenia (threshold) dla cząsteczek lotnych związków jest
bardzo różny. Stosunkowo słabo reagują komórki węchowe na eter
etylowy (próg 79 umol/L powietrza), silniej na kwas masłowy (próg
102 nmol/L powietrza) i najsilniej na merkaptan metylowy (próg 831
fmol/L powietrza).
Ryc. 67. Drogi przewodzące impulsację węchową. (Reprodukowano za zezwol eniem
z S. Różycki Anatomia mózgowia i rdzenia kręgowego, VI wyd. Państwowy Zakład
Wydawnictw Lekarskich, Warszawa 1960.)
Adaptacja (adaptation) do zapachu zależy od budowy chemicznej
cząsteczek wywołujących czucie zapachu. Do pewnych zapachów ludzie
adaptują się szybko, przestając je odczuwać, do innych bardzo wolno.
W przypadku działania dwóch zapachów odbierany jest ten, który działa
silniej lub do którego wolniej przebiega adaptacja. Człowiek może
odróżnić od 2000 do 4000 różnych zapachów.
Czucie teleceptywne
1 33
Pomiędzy komórkami nerwowo-zmysłowymi węchowymi nabłonka
węchowego znajdują się nagie zakończenia włókien nerwowych należących do nerwu trójdzielnego. Podrażnienie ich wywołuje odruch kichania
(sneezing reflex).
I Zmysł wzroku
W odbieraniu fal świetlnych i ich przekształceniu na wrażenie zmysłowe bierze udział narząd odbiorczy, czyli oko, oraz drogi i ośrod ki łączące oko z polem wzrokowym kory mózgu. Samo oko spełnia
złożoną funkcję. Zawiera układ optyczny, właściwe receptory — czyli
pręciki i czopki — oraz drogi dla impulsów nerwowych w samej siatkówce.
Gałka oczna {bulbus oculi) zbudowana jest z trzech zasadniczych
warstw. Warstwę zewnętrzną tworzy błona włóknista gałki ocznej (tunica
fibrosa bulbi), w części przedniej przezroczysta, czyli rogówka (cornea),
i w pozostałej części nieprzezroczysta, stanowiąca twardówkę (selera).
Warstwa środkowa jest utworzona z naczyniówki (choroidea), a w części
przedniej z ciała rzęskowego (corpus ciliare) i tęczówki (iris). Błona
wewnętrzna gałki ocznej (tunica interna bulbi) w przeważającej części
jest utworzona z siatkówki (retina).
Układ optyczny oka
Układ optyczny oka (eye optical system) utworzony jest ze struktur
i płynów przejrzystych, załamujących promienie świetlne. Są to w kierunku od zewnątrz gałki ocznej: rogówka (cornea), ciecz wodnista (humor
aąuosus) — wypełniająca komorę przednią gałki ocznej (camera anterior
bulbi) — soczewka (lens) i ciało szkliste (corpus vitreum).
Promienie równoległe padając na rogówkę, załamują się i skupiają
w ognisku leżącym na siatkówce. Oś układu optycznego oka (optic axis)
nie pokrywa się z osią patrzenia (visual axis). Oś patrzenia biegnie
przez rogówkę bardziej przyśrodkowo w stosunku do osi optycznej oka.
W soczewce obie osie krzyżują się i oś patrzenia pada na dołek środkowy (fovea centralis) plamki żółtej siatkówki (macula lutea retinae), gdzie
występuje największe zagęszczenie komórek receptorowych w kształcie
czopków.
1 34
Czucie i ruch
Ryc. 68. Przekrój poziomy prawej gałki ocznej. Linia przerywana — oś patrzenia. Strzałki
wskazują kierunek przepływu cieczy wodnistej z komory tylnej do komory przedniej.
(Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie
PZWL, Warszawa 1997.)
Siła załamywania układu optycznego oka człowieka w czasie patrzenia
w dal, a więc dla promieni równoległych, wynosi +66,7 dioptrii. Obraz,
jaki powstaje na siatkówce po przejściu promieni przez układ optyczny
oka, jest rzeczywisty, pomniejszony i odwrócony.
Akomodacja oka
Siła załamywania układu optycznego związana jest przede wszystkim
z krzywizną rogówki, a w mniejszym stopniu z krzywiznami soczewki.
W czasie patrzenia na przedmioty bliskie soczewka zmienia swój kształt,
co jest związane z akomodacja. Krzywizny soczewki zwiększają się:
w większym stopniu krzywizna przedniej powierzchni soczewki, a w mniejszym stopniu krzywizna tylnej powierzchni soczewki. Na skutek zmiany
kształtu soczewki zwiększa się siła załamywania układu optycznego oka.
Czucie teleceptywne
1 35
Układ optyczny oka, dzięki zmianie siły załamywania soczewki, jest
zdolny do skupienia w ognisku leżącym na siatkówce zarówno promieni
równoległych od odległych przedmiotów, jak i rozbieżnych od bliższych
przedmiotów, aż do najbliższych znajdujących się w najbliższym punkcie
widzenia (near point of vision). W tym zakresie układ optyczny oka
nastawia się, czyli akomoduje, do patrzenia na przedmioty znajdujące się
w różnej odległości od oka.
Zmiana siły załamywania soczewki następuje odruchowo. W czasie
patrzenia na przedmioty odległe mięsień rzęskowy {musculus ciliaris) jest
rozkurczony, a przyczepiona promieniście do torebki soczewki obwódka
rzęskowa (wiązadełka Zinna) jest napięta. Soczewka jest wtedy spłaszczona. W czasie patrzenia na przedmioty bliskie mięsień rzęskowy się
kurczy, obwódka rzęskowa rozluźnia i soczewka przybiera kształt
bardziej kulisty. Siła załamywania soczewki zwiększa się. Przy maksymalnym zwiększeniu krzywizn soczewki siła załamywania układu optycznego oka zwiększa się o około 12 dioptrii. Tak może zwiększać się siła
załamywania oko dziecka 8-letniego (66,7 dioptrii+12 dioptrii) i dzięki
temu jego najbliższy punkt widzenia znajduje się w odległości 8,6 cm
od rogówki. U dziecka w tym wieku patrzącego na przedmiot z tej
odległości promienie świetlne skupiają się w ognisku leżącym na siatkówce.
Ryc. 69. Zmiana krzywizny soczewki w czasie akomodacji oka: a — krzywizny soczewki
w czasie patrzenia na przedmioty odległe; b — zmniejszenie promieni krzywizn soczewki
w czasie patrzenia na przedmioty bliskie.
W miarę wzrostu i starzenia się organizmu torebka soczewki stopniowo
traci swoją sprężystość i właściwość nadawania soczewce bardziej kulistego
kształtu, co jest przyczyną stopniowego odsuwania się od rogówki
najbliższego punktu widzenia. W wieku 20 lat znajduje się on już
w odległości 10,4 cm, a w wieku 60 lat — w odległości 83,3 cm. Patrzenie na
bliskie przedmioty wymaga korekcji za pomocą szkieł.
1 36
Czucie i ruch
Konwergencja oczu
W czasie kierowania wzroku na przedmiot bardzo odległy osie patrzenia obu oczu są ustawione prawie równolegle. W miarę zbliżania się
obserwowanego przedmiotu gałki oczne odruchowo ustawiają się w ten
sposób, że osie patrzenia przecinają się. Nosi to nazwę konwergencji.
Im bliżej oczu znajduje się obserwowany przedmiot, tym większy jest
kąt, pod którym przecinają się osie patrzenia.
Kontrola wielkości źrenicy
Średnica źrenicy kontrolowana jest odruchowo i jej wielkość zależy
od natężenia promieni świetlnych padających na siatkówkę oraz od
odległości obserwowanego przedmiotu. Zwiększenie natężenia światła
i zbliżanie się obserwowanego przedmiotu wywołują skurcz mięśnia
zwieracza źrenicy (musculus sphincter pupillae) znajdującego się w tęczówce (iris). Przeciwnie — zmniejszenie natężenia światła lub oddalanie
się obserwowanego przedmiotu wywołują skurcz mięśnia rozwieracza
źrenicy (musculus dilatator pupillae) i źrenica się rozszerza.
Impulsy nerwowe w odruchowym zwężeniu źrenicy pod wpływem
światła i w czasie akomodacji, podczas patrzenia na bliskie przedmioty,
biegną przez dwa odmienne łuki odruchowe. W niektórych schorzeniach
(kiła układu nerwowego) źrenice w czasie akomodacji zwężają się i nie
reagują na światło.
Wady refrakcji oczu
Oko miarowe (emmetropia) jest to oko, którego układ optyczny skupia
na siatkówce promienie świetlne biegnące równolegle. W przeciwieństwie
do oka miarowego oko niemiarowe (ametropia) skupia promienie równoległe przed lub za siatkówką.
W oku niemiarowym krótkowzrocznym (myopia) układ optyczny zbyt
silnie załamuje promienie i obraz ostry tworzy się w ciele szklistym przed
siatkówką. Natomiast na siatkówce tworzy się obraz nieostry.
W oku niemiarowym nadwzrocznym (hypermetropia) promienie równoległe są słabiej załamywane przez układ optyczny i przecinają się za
siatkówką. Obraz tworzący się na siatkówce jest nieostry.
Korekcję wad refrakcji oczu przeprowadza się za pomocą szkieł.
W przypadku krótkowzroczności stosuje się szkła o soczewkach rozpra-
Czucie teleceptywne
1 37
Ryc. 70. Schemat dróg i ośrodków kontrolujących szerokość źrenic: linia przerywana
czerwona — drogi i ośrodki łuku odruchowego rozszerzania źrenicy, linia przerywana czarna — drogi i ośrodki łuku odruchowego odruchu zwężania źrenicy, linia z punktami —
część łuku odruchowego zwężania źrenicy w czasie patrzenia na przed mioty bliskie.
C8 — ósmy segment części szyjnej rdzenia kręgowego.
szających, w przypadku nadwzroczności zaś szkła o soczewkach skupiających.
Wadą refrakcji jest również niezborność oka (astigmatismus). W oku
prawidłowym rogówka ma nieco krótszy promień krzywizny w płaszczyźnie pionowej w porównaniu z promieniem krzywizny w płaszczyźnie
poziomej. W przypadku większej różnicy długości promieni krzywizny rogówki w różnych płaszczyznach promienie świetlne skupiają się
w dwóch lub więcej ogniskach. Obraz tworzący się na siatkówce nie jest
ostry. Korekcję tej wady przeprowadza się za pomocą szkieł o soczewkach
cylindrycznych.
1 38
Czucie i ruch
Ryc. 71. Oko miarowe (a) i wady refrakcji: b — oko
niemiarowe krótkowzroczne; c — oko niemiarowe krótkowzroczne po korekcji soczewką wklęsłą; d — oko
niemiarowe nadwzroczne; e — oko niemiarowe nadwzroczne po korekcji soczewką wypukłą; / — oko
niemiarowe niezborne; g — oko niemiarowe niezborne,
astygmatyczne, po korekcji soczewką cylindryczną.
Ryc. 72. Komórka wzrokowa pręcikonośna (po lewej) i komórka wzrokowa czopkonośna
(po prawej). (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Czucie teleceptywne
1 39
Czynność siatkówki
Zasadniczymi elementami czynnościowymi siatkówki są komórki nerwowe skupione w trzech warstwach. Warstwę zewnętrzną siatkówki
stanowią neurony wzrokowe o wypustkach w postaci czopków i pręcików, tzw. neurony wzrokowe czopkonośne i pręcikonośne. Warstwę środkową tworzą neurony dwubiegunowe, a warstwę wewnętrzną — sąsiadująca z ciałem szklistym — neurony wzrokowo-zwojowe.
Receptorami wrażliwymi na falę świetlną, czyli fotoreceptorami, są
pręciki {bacillus) i czopki (conus). Pod wpływem fali świetlnej w pręcikach
i czopkach dochodzi do przemian chemicznych i powstający impuls
przewodzony jest przez neurony dwubiegunowe, a następnie przez neurony wzrokowo-zwojowe, których aksony biegną w nerwie wzrokowym.
Pręciki
Pręcik jest to wypustka neuronu wzrokowego pręcikonośnego (cellula
optica bacillifera), w której występują spłaszczone pęcherzyki gładkiej
siateczki śródplazmatycznej w postaci dysków, wypełnione barwni kiem — rodopsyną (rhodopsin), czyli czerwienią wzrokową (visual purple).
Barwnik ten pochłania światło niebiesko-zielone, czyli fale świetlne
o maksimum pochłaniania w zakresie około 500 nm. Pod wpływem światła
pochodna rodopsyny — metarodopsyna I — zmienia się na metarodopsynę II i następnie metarodopsynę III, która rozpada się na całkowi-cietrans-retinal (all-trans retinal) i białko opsynę (opsin). Jednocześnie błona
komórkowa pręcików hiperpolaryzuje się, ponieważ zamykają się kanały
dokomórkowego prądu jonów Na +. W ciemności jony Na + stale
napływają do fotoreceptorów, w czym bierze udział cykliczny guanozynomonofosforan (cGMP).
Pod wpływem światła i przy udziale transducyny (transducin), należącej do białek błonowych G, zostaje zaktywowany enzym fosfodiesteraza, która metabolizuje cGMP. W ciemności dochodzi do resyntezy
rodopsyny z 11-cis-retinalu i opsyny, inaktywacji fosfodiesterazy, zwiększenia zawartości cGMP i do otwierania się kanałów dokomórkowego
prądu jonów Na +. Błona komórkowa fotoreceptorów powraca do spoczynkowej polaryzacji. Pręciki odbierają różnice w natężeniu promieni
świetlnych padających na różne miejsca siatkówki. Obraz odbierany przez
pręciki jest nieostry, ponieważ jeden neuron wzrokowo-zwojowy odbiera
impulsy od wielu neuronów dwubiegunowych, te zaś mają synapsy
z 30—50 neuronami wzrokowymi pręcikonośnymi.
1 40
Czucie i ruch
hiperpolaryzacja błony
komórkow ej fotoreceptor ów
Ryc. 73. Schemat rozpadu rodopsyny pod wpływem światła i jej resynteza.
Czopki
Neuron wzrokowy czopkonośny {cellula optica conifera), podobnie
jak neuron wzrokowy pręcikonośny, wypełniony jest barwnikiem składającym się z retinalu oraz z jednego z trzech różnych białek — opsyn.
W zależności od budowy cząsteczki białka czopki pochłaniają światło
niebieskie (maksimum pochłaniania około 420 nm), światło zielone
(maksimum pochłaniania około 530 nm) lub światło czerwono-żółte
(maksimum pochłaniania około 560 nm). Pod wpływem fal świetlnych
o określonej długości w jednym z trzech typów czopków dochodzi do
rozpadu barwnika na opsynę i całkowicie-trans-retinal, tak jak w pręcikach.
Czopki zapewniają nie tylko odbiór światła o różnej długości fali,
lecz również ostrość widzenia. Największa ostrość widzenia jest uzyskiwana podczas padania obrazów na okolicę dołka środkowego siatkówki
Czucie teleceptywne
1 41
(fovea centralis), a zwłaszcza w jego części środkowej zajmowanej przez
plamkę żółtą {macula lutea). W tej okolicy siatkówki występują tylko
czopki, które są rozmieszczone tak gęsto, że przylegają do siebie.
Neuronalna organizacja siatkówki
W warstwie splotowatej zewnętrznej stykają się ze sobą wypustki
neuronów pręcikonośnych z neuronami poziomymi (neurocytus horizontalis) i neuronami dwubiegunowymi {neurocytus bipolaris). W wyniku
wzajemnego oddziaływania na siebie tych trzech neuronów, neuron
dwubiegunowy depolaryzuje się i przewodzi impuls nerwowy do neuronu
wzrokowo-zwojowego {neurocytus ganglionaris).
W warstwie splotowatej wewnętrznej stykają się ze sobą neurony
dwubiegunowe z neuronami wzrokowo-zwojowymi oraz z neuronami
amakrynowymi {neurocytus amacrinus) i również tu dochodzi do wzajemnego oddziaływania na siebie tych neuronów.
Neuron wzrokowy czopkonośny przekazuje stan czynny do dwóch
neuronów dwubiegunowych. Każdy z tych neuronów dwubiegunowych
styka się z jednym neuronem wzrokowo-zwojowym. Promień światła,
padając na neuron wzrokowy czopkonośny, aktywuje jeden neuron
dwubiegunowy — „zapalający" („on celi"). Zmniejszenie się intensywności światła padającego na czopek aktywuje drugi neuron dwubiegunowy — „gasnący" („off celi"). Każdy z tych dwóch neuronów dwubiegunowych przekazuje stan czynny do innego neuronu wzrokowo-zwojowego. Dzięki temu przez oddzielne włókna nerwowe w nerwach wzrokowych biegną do mózgu impulsy wywołane zwiększeniem lub zmniejszeniem intensywności światła padającego na poszczególne punkty siatkówki. Takie kodowanie obrazu padającego na siatkówkę zapewnia ostre
widzenie konturów.
Pomiędzy warstwą splotowatą zewnętrzną i warstwą splotowatą wewnątrzną siatkówki występują neurony poziome, neurony amakrynowe
i neurony międzysplotowe. Neurony poziome tworzą synapsy w warstwie
splotowatej zewnętrznej z neuronami pręcikonośnymi i czopkonośnymi.
Neurony amakrynowe wysyłają swoje wypustki do warstwy splotowatej
wewnętrznej, tworząc synapsy z neuronami dwubiegunowymi i neuronami wzrokowo-zwojowymi.
Trzeci typ neuronów — neurony międzysplotowe — ma wypustki
do obu warstw siatkówki i przekazuje informacje od warstwy splotowej wewnętrznej do warstwy splotowej zewnętrznej, a więc w kierunku przeciwnym w stosunku do neuronów dwubiegunowych. Czyn-
1 42
Czucie i ruch
ność tych trzech typów neuronów wiąże się z modulacją w obrębie
siatkówki impulsacji wysyłanej przez fotoreceptory.
Pomiędzy neuronami siatkówki występują zarówno synapsy elektryczne, jak i synapsy chemiczne. Zasadniczymi transmitterami w synapsach
chemicznych szybko przewodzących stan czynny są glutaminiany, asparaginiany i acetylocholina. W synapsach szybko wyzwalających hiperpolaryzację błony postsynaptycznej uwalniane są kwas gamma-aminomasłowy i glicyna.
Warstwo barwnikowa
Warstwa^ pręcików i c
zopków
Warstwa jądrowa
zewnętrzna
Warstwa splofowata
zewnętrzna
Warstwa jqdrowa
wewnętrzna
Warstwa splotowata
wewnętrzna
Warstwa zwojowa
Warstwa graniczna
wewnętrzna
Ryc. 74. Neuronalna organizacja siatkówki. Schematycznie przedstawione są połączenia
pomiędzy komórkami nerwowymi siatkówki: neuron czopkonośny w spoczynku (1), neuron
czopkonośny pobudzony narastającym natężeniem światła (2), neuron czopkonośny pobudzony zmniejszającym się natężeniem światła (3), neuron pręcikonośny (4), neuron dwubiegunowy (5), neuron dwubiegunowy pobudzony narastającym natężeniem światła (6),
neuron dwubiegunowy pobudzony zmniejszającym się natężeniem światła (7), neuron
poziomy (8), neuron amakrynowy (9), neuron międzysplotowy (10), mały neuron zwojowy
(11), duży neuron zwojowy (12).
Czucie te/eceptywne
1 43
Poza transmitterami rolę modulatorów synaptycznych w siatkówce
spełniają aminy — dopamina i serotonina — oraz peptydy — przedsionkowy peptyd natriuretyczny (ANP), cholecystokinina (CCK), enkefaliny (Enk), amid (Phe-Met-Arg-Phe-NH2), glukagon, hormon uwalniający gonadotropiny (GnRH), neurotensyna, neuropeptyd Y (NPY),
somatostatyna (SRIF), Substancja P i wazoaktywny peptyd jelitowy
(VIP).
Pomiędzy neuronami siatkówki występują komórki glejowe (gliocytus
radialis) zajmujące całą szerokość siatkówki od warstwy granicznej
zewnętrznej {stratum limitans externum) do warstwy granicznej wewnętrznej {stratum limitans internum). W warstwie granicznej zewnętrznej komórki glejowe mają liczne wypustki otaczające czopki i pręciki. Pełnią funkcję podporową i odżywczą w stosunku do neuronów
siatkówki.
Pomiędzy naczyniówką i warstwą pręcików i czopków znajduje się
warstwa barwnikowa {stratum pigmentosum), która pochłania promienie
świetlne wpadające do gałki ocznej i zapobiega ich odbiciu i rozpr oszeniu. Dzięki pochłanianiu promieni świetlnych przez warstwę barwnikową obraz padający na siatkówkę ma ostre kontury. Brak barwnika
(albinosi) lub jego niedobór powoduje rozproszenie się promieni wewnątrz gałki i zacieranie konturów. Poprawę ostrości wzroku można
uzyskać w tym przypadku, nosząc ciemne okulary, które częścio wo
zapobiegają odbijaniu się i rozpraszaniu światła wewnątrz gałki ocznej.
Człowiek odbiera oddzielne obrazy, jeśli występują z częstotliwością
do 10 Hz. W przypadku większej częstotliwości obrazy nie są odbierane
oddzielnie, lecz jak jeden obraz, ponieważ resynteza barwnika w fotoreceptorach trwa 0,1 s. Zjawisko to jest wykorzystywane przez
film i telewizję. Natomiast działanie światła na siatkówkę może trwać
bardzo krótko. Można obserwować szczegóły na przedmiotach szybko
wirujących, jeśli są one oświetlone za pomocą lampy stroboskopowej na
bardzo krótki czas, około kilku milisekund.
Ostrość wzroku
Ostrość wzroku (acuity of vision), czyli siła rozdzielcza oka, jest
oceniana na podstawie obrazu padającego na plamkę żółtą. Odróżnienie
dwóch świecących punktów leżących blisko siebie zachodzi tylko wtedy,
kiedy dwa czopki zaktywowane przez promienie wysyłane przez te punkty
są przedzielone czopkiem nie zaktywowanym. Średnica czopka w plamce
1 44
Czucie i ruch
żółtej wynosi od 2 do 3 um, tym samym oko odbiera dwa punkty
świecące z odległości 10 m jako niezależne, jeśli są oddzielone od siebie
o 1 mm.
Adaptacja siatkówki
Fotoreceptory siatkówki adaptują się, czyli przystosowują, zarówno do
warunków odbierania promieni świetlnych o dużym, jak i o małym
natężeniu.
Adaptacja do dużego natężenia, czyli adaptacja do silnego światła (light
adaptation), osiąga swoje maksimum w czasie około 10 minut. W zaadaptowanej do światła siatkówce przeważająca część fotoreceptorów ma
rozłożony barwnik i tym samym nie wysyłają one impulsów nerwowych.
Barwnik w fotoreceptorach zostaje znacznie szybciej rozłożony niż jest
resyntezowany i większość fotoreceptorów jest niepobudliwa.
Przeciwne zjawisko zachodzi w czasie adaptacji do ciemności (dark
adaptation). W całkowitej ciemności dochodzi do resyntezy barwnika
we wszystkich fotoreceptorach, tak że minimalne natężenie promieni jest
odbierane przez maksymalną liczbę fotoreceptorów. Adaptacja do ciemności trwa ponad godzinę i w tym czasie pobudliwość siatkówki może
zwiększyć się do 100 tysięcy razy w porównaniu z siatkówką zaadaptowaną do silnego światła.
Pole widzenia
Pole widzenia (visual field) jest to obraz przestrzeni otaczającej padający na siatkówkę oka i wyzwalający wrażenia wzrokowe. Pole widzenia
dla promieni o różnej długości fali, czyli dla barwy białej, jest największe. Dla promieni monochromatycznych pole widzenia jest mniejsze,
co jest spowodowane nierównomiernym rozmieszczeniem fotoreceptorów
wrażliwych na poszczególne barwy w obrębie siatkówki. W obrębie krążka
nerwu wzrokowego {discus nervi optici) nie ma fotoreceptorów i nie są
odbierane z tej okolicy wrażenia wzrokowe. Brak receptorów w tej okolicy powoduje występowanie w polu widzenia tzw. plamki ślepej (blind
spot).
Pola widzenia dla prawego i lewego oka w środkowej większej części
pokrywają się ze sobą. Dzięki temu występuje widzenie obuoczne (binocular vision), które pozwala na ocenę odległości i wielkości oglądanych
Cz ucie telec e pty w ne
1 45
Ryc. 75. Pole widzenia prawego oka w odniesieniu do barwy białej, niebieskiej, czerwonej
i zielonej (a). Pole widzenia obu oczu (6).
przedmiotów. Obraz przedmiotu padający na siatkówki obu oczu nieznacznie się różni, co nosi nazwę zjawiska paralaksy i jest spowodowane
różnicą w kącie patrzenia obu oczu. Widzenie stereoskopowe zapewnia
zdolność oceny trójwymiarowej oglądanych przedmiotów, a tym samym
ich wielkości.
Elektrookulogram i elektroretynogram
Pomiędzy rogówką i siatkówką występuje stała różnica potencjału
elektrycznego. Rogówka ma ujemny, siatkówka zaś dodatni potencjał
elektryczny. Ruchy gałek ocznych, szczególnie w płaszczyźnie poziomej,
wywołują zmiany pola elektrycznego wytworzonego przez różnicę potencjału elektrycznego rogówkowo-siatkówkowego. Zmiany pola elektrycznego spowodowane ruchem gałek ocznych można odebrać przy użyciu
elektrod umieszczonych na skórze w okolicach oczodołów i zarejestrować
w postaci zapisu — elektrookulogramu — ECG (electrooculogram).
Pod wpływem światła padającego na pręciki i czopki następuje przechodzenie jonów przez błonę komórkową tych receptorów, a następnie
ruch jonów w płynie zewnątrzkomórkowym w siatkówce. Wahania
potencjału elektrycznego wywołane przemieszczaniem się jonów w siatkówce pod wpływem światła można odebrać przy użyciu aktywnej
1 46
Czucie i ruch
elektrody umieszczonej na powierzchni rogówki w postaci soczewki
kontaktowej.
Błysk światła padającego na pręciki i czopki wywołuje pojawienie się
ujemnego potencjału elektrycznego na powierzchni rogówki w postaci
fali a. Po fali a występuje fala b, o dodatnim ładunku, wywołana
przewodzeniem stanu czynnego przez neurony dwubiegunowe siatkówki
i zwiększeniem aktywności dużych podporowych komórek glejowych
(komórek Miillera). Po fali b pojawiają się fale oscylujące związane
z przewodzeniem stanu czynnego przez neurony zwojowe siatkówki.
Potencjały elektryczne wywołane światłem padającym na receptory siatkówki zapisywane są w postaci elektroretynogramu — ERG (electroretinogram).
Ryc. 76. Elektroretynogram (ERG — electroretinogram) z oczu zaadaptowanych do
ciemności (adaptacja skotopowa) wywołany: a — błyskiem światła monochromatycznego
(niebieskie — n); b — błyskiem światła monochromatycznego (czerwone — c): zaznacza się
fala b, — czopkowa i b 2 — pręcikowa; c — błyskiem światła polichromatycznego
(białe — p'); d — po adaptacji do światła dziennego błyskiem światła polichromatycznego (białe — p"). (Otrzymano od Olgierda Palacza z Katedry i Kliniki Pomorskiej Akademii
Medycznej w Szczecinie.)
Przewodzenie impulsów z siatkówki
Aktywacja neuronów wzrokowych czopkonośnych i pręcikonośnych
(I neuron czuciowy) jest przekazywana za pośrednictwem neuronów
dwubiegunowych (II neuron czuciowy) i neuronów wzrokowo-zwojowych (III neuron czuciowy) siatkówki do ciała kolankowatego bocznego
(corpus geniculatum laterale), gdzie znajduje się IV neuron czuciowy.
Czucie teleceptywne
1 47
Ryc. 77. Droga impulsów z siatkówki do kory mózgu okolicy bruzdy ostrogowej. Brakuje
całego lub części pola widzenia po uszkodzeniu: A — nerwu wzrokowego, B — włókien
skrzyżowanych w skrzyżowaniu wzrokowym, C — włókien nie skrzyżowanych w skrzyżowaniu wzrokowym, D — drogi wzrokowej, E, F i G — promienistości wzrokowej.
1 48
Czucie i ruch
Wypustki III neuronu czuciowego biegnące w nerwie wzrokowym (nervus
opticus) po wejściu do jamy czaszki dochodzą do skrzyżowania wzrokowego i część z nich krzyżuje się i dalej biegną w paśmie wzrokowym
(tractus opticus). Włókna biegnące z części bocznej siatkówki nie są
skrzyżowane, natomiast biegnące z części przyśrodkowej krzyżują się.
Impulsacja z siatkówki jest przekazywana z ciała kolankowatego
bocznego poprzez promienistość wzrokową (radiatio optica) do kory
mózgu, pola 17 według klasyfikacji Brodmanna, powyżej i poniżej bruzdy
ostrogowej (sulcus calcarinus), gdzie jest odbierana przez V neuron
czuciowy.
Zniszczenie dróg i ośrodków przewodzących impulsację z siatkówki
powoduje odpowiedni ubytek całego lub części pola widzenia jednego
oka lub części pól widzenia obu oczu w zależności od miejsca uszkodzenia.
Percepcja wrażeń wzrokowych
Odbiór i rozpoznawanie wrażeń wzrokowych jest procesem złożonym,
który przebiega zarówno w obrębie ciała kolankowatego bocznego
i wzgórka górnego blaszki pokrywy (colliculus superior laminae tecti),
jak i w korze mózgu płata potylicznego, w polu 17 oraz sąsiadujących
z nim polach 18 i 19, w częściach dolnych płata skroniowego i w płacie
ciemieniowym w części tylno-dolnej.
Ryc. 78. Widzenie stereoskopowe: a —
prawe i lewe oko odbiera nieco różny obraz
pudełka zapałek, co pozwala na ocenę jego
odległości od oczu i jego wielkości; b —
obraz pudełka odbierany przez lewe oko; c —
obraz pudełka odbierany przez prawe oko.
Czucie teleceptywne
1 49
W korze mózgu w polach wzrokowych przebiegają procesy nerwowe
spowodowane odbiorem konturów przedmiotów. Oko znajduje się w stałym ruchu i w związku z tym coraz to inne neurony korowe zostają
pobudzone przez salwy impulsów o dużej częstotliwości wywołane przez
przesuwający się na siatkówce obraz konturów przedmiotu. Widzenie
barwne i stereoskopowe oglądanych przedmiotów jest uzależnione od
połączeń neuronów ośrodków wzrokowych w korze mózgu z neuronami
ciała kolankowatego i wzgórka górnego blaszki pokrywy.
W korze mózgu powyżej i poniżej bruzdy ostrogowej (sulcus calcarinus), czyli w polu 17, mieści się pierwszorzędowy korowy ośrodek
wzrokowy. Z tego ośrodka impulsy nerwowe wywołane światłem padającym na siatkówkę są przekazywane do drugo- i trzeciorzędowego
ośrodka wzrokowego, w polu 18, a stąd do ośrodków korowych jeszcze
wyższego rzędu. W ośrodkach tych, znajdujących się w płacie skroniowym i ciemieniowym, zachodzą procesy związane z zapamiętywaniem
nowego obrazu padającego na siatkówkę, jak również z porównywaniem
nowego obrazu, aktualnie odbieranego przez siatkówkę, z wcześniej
zapamiętanym.
Pola korowe wzrokowe w dolnych częściach płata skroniowego sąsiadują z polami kory płatów skroniowych i potylicznych położonych
najbardziej przyśrodkowo na dolnej powierzchni mózgu. W tych polach
korowych zachodzą procesy związane z percepcją obrazów odbieranych
przez siatkówkę, szczególnie z rozpoznawaniem ludzi na podstawie
wyglądu ich twarzy.
Kontrola ruchów gałki ocznej
Do gałki ocznej przyczepionych jest sześć mięśni gałki ocznej unerwionych przez nerw okoruchowy — III {nervus oculomotorius), nerw
bloczkowy — IV (nervus trochlearis) i nerw odwodzący — VI (nervus
abducens). Dzięki tym mięśniom gałka oczna wykonuje ruchy w płaszczyznach poziomej i pionowej oraz ruchy obrotowe.
Gałki oczne wykonują cztery zasadnicze rodzaje ruchów:
• ruchy szybkie, występujące podczas przenoszenia wzroku z jednego
przedmiotu na drugi;
• ruchy wolne, takie jak w czasie obserwowania poruszającego się
przedmiotu;
• ruchy wyzwalane przez impulsację z przedsionków;
• ruchy konwergencyjne.
1 50
Czucie i ruch
Ryc. 79. Ruchy prawego oka
(widok na gałkę od przodu).
Mięśnie gałki ocznej: mięsień
skośny górny — m.s.g., mięsień
skośny dolny — m.s.d., mięsień
prosty górny — m.p.g., mięsień
prosty dolny — m.p.d., mięsień
prosty boczny — m.p.b., mięsień prosty przyśrodkowy m.p.p.
Spostrzeganie dowolnego przedmiotu wymaga, aby jego obraz na
siatkówce stale się przesuwał. Dlatego gałki oczne u człowieka są w stałym
ruchu, nawet w czasie snu.
Przy wzroku skierowanym na nieruchomy przedmiot, gałki oczne
wykonują nieznaczne powolne ruchy, po których następują szybkie
ruchy, powodujące powrót do początkowego ustawienia gałek ocznych.
Jest to oczopląs fizjologiczny (physiologic nystagmus). Ruchy gałek
ocznych w postaci oczopląsu występują również po podrażnieniu receptorów przedsionka (yestibulum).
Impulsacja z receptorów przedsionków kontroluje stale aktywność
neuronów ruchowych w jądrach ruchowych dla mięśni gałek ocznych.
Dzięki temu niezależnie od ruchów głowy wzrok pozostaje skierowany
na ten sam przedmiot.
W czasie przenoszenia wzroku z przedmiotów odległych na bliskie
gałki oczne wykonują ruch, tak że osie patrzenia początkowo ustawione
równolegle zaczynają się przecinać, co nosi nazwę konwergencji. Jednocześnie z ruchem gałek ocznych, dzięki skurczowi mięśnia rzęskowego,
występuje akomodacja soczewki oraz zwęża się źrenica.
I Zmysł słuchu
Właściwy receptor odbierający fale akustyczne znajduje się w uchu
wewnętrznym (auris interna), w narządzie spiralnym — Cortiego (orga-
Czucie teleceptywne
151
num spirale). Tam zachodzi przetworzenie fal akustycznych na impulsy
nerwowe.
Fale akustyczne przewodzone są przez powietrze znajdujące się w przewodzie słuchowym zewnętrznym (meatus acusticus externus). Na swej
drodze fale akustyczne napotykają błonę bębenkową {membrana tympani)
i wywierają na nią zmienne ciśnienie. Drgania błony bębenkowej pod
wpływem fal akustycznych przenoszone są w uchu środkowym {auris
media) na okienko przedsionka {fenestra vestibuli) przez dźwignię utworzoną z trzech kosteczek słuchowych {ossicula auditus): młoteczka {malleus), kowadełka {incus) i strzemiączka {stapes).
Powierzchnia błony bębenkowej jest około 22 razy większa od powierzchni podstawy strzemiączka {basis stapedis) zamykającej okienko
przedsionka. Dzięki temu energia fal akustycznych odebranych przez
większą powierzchnię błony bębenkowej jest przeniesiona za pośrednictwem dźwigni z kosteczek słuchowych na małą powierzchnię podstawy
strzemiączka. Drgania błony bębenkowej wywołują fale zmian ciśnienia
i ruch przychłonki wypełniającej schody przedsionka {scala vestibuli),
przenoszonej do schodów bębenka {scala tympani) przez szparę osklepka
{helicotrema) łączącą schody przedsionka ze schodami bębenka. Nacisk
podstawy strzemiączka na przychłonkę wywołuje uwypuklenie się błony
zamykającej okienko ślimaka {fenestra cochleae) do jamy bębenkowej
{cavum tympani). Następnie fala ciśnienia powraca w kierunku przeciwnym.
Łagiewka i plamka
Ryc. 80. Schemat budowy błędnika błoniastego składającego się z przewodu ślimakowego,
należącego do narządu słuchu, oraz woreczka, łagiewki i przewodów półkolistych, należących
do narządu równowagi. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd.
Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
152
Czucie i ruch
Ucho
Ucho
Ucho
wewnętrzne
zewnętrzne środkowe
TrąbHa słuchowa
Ryc. 81. Rezonans fal akustycznych. Odkształcanie się blaszki podstawnej pod wpływem
maksymalnej amplitudy fali ciśnienia przychłonki.
Ryc: 82. Schemat budowy przewodu ślimakowego (przekrój poprzeczny). (Reprodukowano
za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa
1997.)
Czucie te/eceptywne
1 53
Pomiędzy schodami przedsionka i schodami bębenka znajduje się
przewód ślimakowy (ductus cochleariś) wypełniony śródchłonką. Oddziela
go od schodów przedsionka cienka błona przedsionkowa (Reissnera),
a od schodów bębenka grubsza blaszka podstawna (lamina basilaris).
Ślimak ma długość 35 mm i jest skręcony 2 i 3/4 raza. W miarę oddalania
się od obu okienek ślimak rozszerza się. Blaszka podstawna, na której
znajduje się narząd spiralny, również staje się szersza.
Rezonans fal akustycznych
Ruch strzemiączka wywołuje falę ciśnienia w przychlonce, której amplituda zależy od intensywności, a miejsce, w którym występuje — od
częstotliwości odbieranych dźwięków. Fale ciśnienia przychłonki, tzw.
fale biegnące (travelling waves), przenosząc się od okienka przedsionka
do szpary osklepka, podwyższają swoją amplitudę aż do miejsca, w którym zachodzi zmiana ich fazy. W miarę zmniejszania się częstotliwości
dźwięków miejsce zmiany fazy oddala się od okienka przedsionka.
W miejscu zmiany fazy fala ciśnienia przychłonki przenosi się na śródchłonkę wypełniającą przewód ślimakowy i blaszka podstawna podlega
największemu odkształceniu.
Komórki zmysłowe wloskowate {cellulae sensoriae pilosae) narządu spiralnego są rozmieszczone na blaszce podstawnej w dwóch rzędach.
Komórki
Bottchera
Zewnętrzne
Zewnętrzna
Wewnętrzna
komórki talangowe
komórka filarowa
Ryc. 83. Schemat budowy narządu spiralnego (Cortiego) (przekrój poprzeczny). (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie
PZWL, Warszawa 1997.)
1 54
Czucie i ruch
W rzędzie po stronie wewnętrznej tunelu (Cortiego) jest 3500, a po
stronie zewnętrznej jest 20 000 komórek zmysłowych włoskowatych.
Komórki te są jak gdyby rozpięte pomiędzy blaszką podstawną i błoną
pokrywającą {membrana tectoria), do której są przylepione ich włoski.
Na swoim przebiegu, pomiędzy ciałem komórki zmysłowej włoskowatej
i błoną pokrywającą, włoski przechodzą przez błonę siatkowatą {membrana reticularis).
Na skutek ruchu blaszki podstawnej, w miejscu zmiany fazy fali
biegnącej, komórki zmysłowe włoskowate zostają pobudzone w wyniku
napinania się włosków pomiędzy błoną siatkowatą i błoną pokrywającą.
Odbiór bodźców słuchowych
Umownie przyjęto mierzyć natężenie fal akustycznych w decybelach.
Bodziec progowy, przy którym można osiągnąć u człowieka próg słyszenia, wynosi 0 decybeli (dB), co się równa ciśnieniu 20 uN/m2 = 20 |iPa.
Pomiędzy progiem słyszenia i bodźcem maksymalnym występuje różnica
w intensywności około 10 14 razy, czyli próg słyszenia — 0 dB, i bodziec
Ryc. 84. Audiogram prawego i lewego ucha.
Czucie teleceptywne
1 55
maksymalny — 140 decybeli. Dźwięki o natężeniu powyżej 140 dB poza
wrażeniem słuchowym wywołują również czucie bólu związane z uszkodzeniem narządu spiralnego.
Ludzie młodzi odbierają dźwięki o częstotliwości w zakresie od 20
do 20 000 Hz. W miarę starzenia się organizmu górna granica obniża się
i dźwięki o dużej częstotliwości nie są słyszalne. Najlepiej odbierane są
dźwięki w zakresie od 1000 do 3000 Hz.
Odruchy bębenkowe
W uchu środkowym znajdują się dwa małe mięśnie, których skurcz
wpływa na przenoszenie drgań błony bębenkowej przez kosteczki słuchowe. Są to mięsień napinacz błony bębenkowej {musculus tensor
tympani) i mięsień strzemiączkowy {musculus stapedius). Skurcz obu tych
mięśni zmniejsza przewodzenie dźwięków. Wzrasta napięcie błony bębenkowej i podstawa strzemiączka zostaje odciągnięta od okienka przedsionka. Odruch bębenkowy występuje w czasie działania silnych dźwięków i zabezpiecza narząd spiralny przed uszkodzeniem.
Potencjał mikrofonowy
Śródchłonka wypełniająca przewód ślimakowy jest bogata w jony
potasowe o dodatnim ładunku elektrycznym i zawiera mniej ujemnych
jonów chlorkowych. Dzięki temu śródchłonka ma ładunek dodatni
+ 80 mV w stosunku do przychłonki. W czasie odbierania dźwięków
w okolicach ślimaka występują wahania potencjału elektrycznego. Fale
akustyczne zostają zamienione bezpośrednio na wahania potencjału
elektrycznego, tak jak w mikrofonie. Jest to zjawisko towarzyszące
odbieraniu fal akustycznych, nie uczestniczące zaś bezpośrednio w przetwarzaniu tych fal na impulsy nerwowe przewodzone przez włókna
nerwowe.
Droga impulsów słuchowych
Każdą komórkę zmysłową włoskowatą oplata siatka włókien ner wowych będących wypustkami komórek dwubiegunowych zwoju spiral-
1 56
Czucie i ruch
Ryc. 85. Drogi przewodzące impulsację słuchową. Neurony przewodzące impulsację słuchową znajdują się w: zwoju spiralnym ślimaka (1), jądrze ślimakowym grzbietowym i brzusznym
(2), jądrach oliwki (3), wzgórku dolnym blaszki pokrywy (4) i ciałach kolankowatych
przyśrodkowych (5).
nego ślimaka (ganglion spirale cochleae). Za pośrednictwem komórek
nerwowych tego zwoju (I neuron czuciowy) i ich wypustek tworzących
nerw ślimakowy (nervus cochleariś), stanowiący część słuchową nerwu VIII
(pars cochleariś nerri vestibulocochlearis), impulsy nerwowe przewodzone
są do mostu do jąder ślimakowych brzusznego i grzbietowego (nuclei
cochleares ventralis et dorsalis), gdzie znajduje się II neuron czuciowy.
W nerwie ślimakowym biegnie około 28 000 włókien nerwowych, które
przewodzą impulsy nerwowe od poszczególnych komórek zmysłowych
włoskowatych. Tym samym poszczególne włókna przewodzą impulsy
nerwowe wywołane tonami o określonej wysokości. W miarę zwiększania się intensywności odbieranego dźwięku zwiększa się również
częstotliwość impulsów nerwowych przewodzonych przez poszczególne
włókna nerwu ślimakowego.
Czucie eksteroceptywne
1 57
W moście, w jądrze brzusznym i grzbietowym ciała czworobocznego
(nucleus ventralis et dorsalis corporis trapezoidei), lub powyżej mostu we
wzgórku dolnym blaszki pokrywy (colliculus inferior laminae tecti) znajduje
się III neuron czuciowy. Neurony jądra ślimakowego grzbietowego
wysyłają wypustki przechodzące na drugą stronę mostu, wypustki neuronów jądra ślimakowego brzusznego biegną zaś po tej samej i po przeciwnej
stronie pnia mózgowia.
Wstęga boczna (lemniscus lateralis) przewodzi impulsy słuchowe do
III lub IV neuronu czuciowego we wzgórku dolnym blaszki pokrywy
i do IV lub V neuronu czuciowego w ciele kolankowatym przyśrodkowym
(corpus geniculatum mediale). Z tej okolicy impulsy słuchowe przez
promienistość słuchową (radiatio acustica) biegną do pola słuchowego
w zakręcie skroniowym górnym (gyrus temporalis superior) w płacie
skroniowym. Według klasyfikacji Brodmanna jest to pole 41 kory mózgu.
Komórkom zmysłowym włoskowatym kolejnych odcinków narządu spiralnego odpowiadają skupienia neuronów czuciowych w polu słuchowym
w korze mózgu. Dzięki temu w polu słuchowym sąsiadują ze sobą
skupienia neuronów odbierające dźwięki wyższe o kolejne oktawy.
Poza czuciową reprezentacją narządu słuchu w polu 41 występują
w jego sąsiedztwie w korze mózgu drugo- i trzeciorzędowe reprezentacje
słuchu. Ich znaczenie polega na różnicowaniu i porównywaniu wysokości
i natężenia odbieranych dźwięków.
I Czucie eksteroceptywne
Właściwości receptorów
Z powierzchni skóry odbierane jest czucie dotyku, ucisku, ciepła,
zimna i bólu. Poza czuciem bólu, które odbierane jest przez nagie
zakończenia nerwowe, pozostałe rodzaje czucia skórnego mają wyspecjalizowane narządy odbiorcze.
Bodziec pobudzający narządy odbiorcze charakteryzuje: siła, czas
narastania i czas jego trwania. Dla powstania wrażenia zmysłowego
najważniejszy jest czas narastania siły bodźca. Im czas narastania jest
krótszy, tym intensywność wrażenia zmysłowego jest większa. W czasie
działania bodźca o tej samej sile występuje zjawisko przystosowania się
receptora do bodźca, czyli adaptacja. Receptory znajdujące się w skórze
adaptują się szybko.
1 58
Czucie i ruch
Ryc. 86. Receptory skórne czucia eksteroceptywnego: a — ciałka dotykowe (Meissnera);
b — ciała blaszkowate (Paciniego); c — łąkotki dotykowe (krążki Merkla). (Reprodukowano
za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa
1997.)
Pojedyncze aferentne włókno nerwowe przewodzi impulsy nerwowe
od jednego rodzaju receptorów. Receptory te zajmują pewną określoną
powierzchnię skóry.
Jednostką czucia (sensory unit) są wszystkie receptory połączone
z pojedynczym włóknem aferentnym. Obszary unerwione przez poszczególne jednostki czucia częściowo zachodzą na siebie, ponieważ receptory,
z których poszczególne włókna nerwowe odbierają impulsację, są często
wzajemnie przemieszczane.
Przewodzenie impulsów przez włókno nerwowe od jednego rodzaju
receptorów jest zgodne z prawem swoistej energii nerwów Johannesa
Miillera. Prawo to obejmuje całą projekcję czuciową od receptora aż
do ostatniego neuronu czuciowego w korze mózgu. Podrażnienie w dowolnym miejscu projekcji czuciowej, włókien lub komórek nerwowych
przewodzących impulsy od określonego receptora, wywołuje wrażenia
zmysłowe specyficzne dla tego receptora.
Czucie eksteroceptywne
1 59
I Czucie dotyku i ucisku
Receptorem czucia dotyku (touch sensation) są ciałka dotykowe
(Meissnera), czucia ucisku zaś ciałka blaszkowate (Paciniego). Impulsy
nerwowe wywołane pobudzeniem tych receptorów przewodzone są przez
zwoje rdzeniowe (I neuron), korzenie grzbietowe i wstępują do mózgowia w sznurze tylnym {funiculus posterior), w pęczku smukłym (fasciculus gracilis) i w pęczku klinowatym (fasciculus cuneatus) do II neuronu
czuciowego w jądrze smukłym (nucleus gracilis) i w jądrze klinowatym
{nucleus cuneatus). Wypustki II neuronu biegną we wstędze przyśrodkowej (lemniscus medialis) do jądra brzusznego tylno-bocznego wzgórza
{nucleus ventralis posterolateralis thalami) do III neuronu czuciowego.
Część impulsów przewodzona jest inną drogą do III neuronu czuciowego we wzgórzu za pośrednictwem II neuronu czuciowego w rogach
tylnych rdzenia kręgowego. Aksony II neuronów czuciowych przechodzą
w spoidle białym {commissura alba) i w spoidle szarym przednim (commissura grisea anterior) na drugą stronę rdzenia i wstępują ku górze
w sznurze bocznym {funiculus lateralis) w drodze rdzeniowo-wzgorzowej
Sekundy 0
1
2
3
i,
5
Ryc. 87. Impulsacja eferentna generowana, pod wpływem bodźca stale działającego, przez
różne rodzaje receptorów w mięśniach i w skórze oraz przez włókno nerwowe.
1 60
Czucie i ruch
Czucie eksteroceptywne
161
bocznej {tractus spinothalamicus lateralis) i w sznurze przednim (funiculus anterior) w drodze rdzeniowo-wzgórzowej przedniej {tractus spinothalamicus anterior). Następnie dochodzą do jądra brzusznego tyl-nobocznego wzgórza {nucleus ventralis posterolateralis thalami), gdzie
znajdują się III neurony czuciowe. Wypustki tego neuronu przebiegają
przez torebkę wewnętrzną, kończąc się na IV neuronie czuciowym
w zakręcie zaśrodkowym kory mózgu (gyrus postcentralis), w polu 1—3
według Brodmanna.
Czuciowe komórki nerwowe i ich wypustki nie są bezładnie przemieszane, ale są rozmieszczone zgodnie z somatotopową organizacją
neuronalną. Od receptorów określonej powierzchni skóry są przewodzone
impulsy w tych samych pęczkach włókien, a neurony czuciowe sąsiadują
ze sobą. Rozmieszczenie III i IV neuronów czuciowych we wzgórzu
i w korze mózgu jest zniekształconym odbiciem powłok całego ciała.
We wzgórzu III neurony w jądrach brzusznych tylno-bocznym i tylno-przyśrodkowym są skupione w przestrzeni trójwymiarowej. W korze
mózgu zaś IV neurony są rozmieszczone równolegle do powierzchni
zakrętu zaśrodkowego. Korowa reprezentacja głowy znajduje się najbardziej bocznie w zakręcie zaśrodkowym, reprezentacja dla kończyn dolnych
jest zaś umiejscowiona przyśrodkowo.
Gęstość, z jaką występują receptory w skórze, jest różna dla poszczególnych okolic. Największe zagęszczenie receptorów występuje w skórze
końca nosa, w opuszkach palców i w wargach. Najmniej receptorów
jest w skórze grzbietu, ud i ramion. Okolice o dużej gęstości receptorów
Ryc. 88. Przenoszenie się potencjału wywołanego drażnieniem skóry na palcach ręki. Salwa
impulsów eferentnych została odebrana w postaci potencjału elektrycznego z nerwu
posrodkowego w okolicy nadgarstka (A), przedramienia (B), łokcia (C), pachy (D), w punkcie
Erba (E), z rdzenia kręgowego na wysokości wyrostka ościstego 7 kręgu szyjnego (F i G)
i z kory mózgu kontralateralnego płata ciemieniowego (H). Przerywana linia pionowa
pomiędzy jasnymi trójkątami wyznacza początek potencjału bioelektrycznego odległego
pola. Przerywana linia pionowa pomiędzy czarnymi trójkątami wyznacza czas salw impulsów
aferentnych wstępujących do rdzenia kręgowego i do kory mózgu. Załamki dodatnie: P9> Pj 1,
P14 i załamki ujemne Nj,, N13, N18, N20. (Reprodukowano za zezwoleniem z J. E. Desmedt.
Non-invasive analysis of the spinał cord generators activated by somatosensory input in man:
near field and far field potentials. Experimental Brain Research. Springer-Verlag, 1984,
Suppl. 9, 45—62.)
6 — Fizjologia człowieka
162
Czucie i ruch
wykazują jednocześnie dużą wrażliwość na dotyk i mają dużą reprezentację korową. Przeciwnie, okolice mało wrażliwe na dotyk są reprezentowane przez małe pola w korze mózgu.
I Czucie ciepła i zimna
Spadek temperatury skóry jest odbierany przez kolbę końcową (Krausego), wzrost temperatury zaś przez położone głębiej w skórze ciałka
zmysłowe (Ruffiniego). Bodźcem progowym dla receptorów zimna jest
spadek temperatury skóry o 0,004°C na sekundę w przedziale od 10 do
41°C, dla receptorów ciepła zaś wzrost temperatury skóry o 0,001°C na
sekundę w przedziale od 20 do 45°C. Receptory ciepła i zimna odbierają
wzrost lub spadek temperatury tylko wtedy, kiedy temperatura otoczenia
różni się od temperatury powierzchni skóry. Przy jednakowej temperaturze powierzchni skóry i otoczenia receptory nie są pobudzane. Stan taki
nosi nazwę zera fizjologicznego.
I neuron czuciowy, przekazujący pobudzenie z receptorów ciepła
i zimna, tułowia i kończyn, znajduje się w zwojach rdzeniowych, II neuron — w rogach tylnych rdzenia kręgowego. Ich aksony przechodzą na
drugą stronę rdzenia i wstępują ku górze w drodze rdzeniowo-wzgórzowej bocznej {tractus spinothalamicus lateralis). III neuron znajduje się
w jądrze brzusznym tylno-bocznym wzgórza. Od receptorów ciepła
i zimna, występujących w obrębie głowy, impulsacja jest przewodzona za
pośrednictwem neuronów zwoju trójdzielnego (ganglion trigeminale),
jąder czuciowych nerwu trójdzielnego i wstęgi trójdzielnej do jądra
brzusznego tylno-przyśrodkowego wzgórza. IV neuron czuciowy znajduje
się w zakręcie zaśrodkowym kory mózgu.
I Czucie bólu powierzchownego
Czucie bólu powierzchownego (superficial pain) wywołują bodźce
uszkadzające skórę, co jest odbierane przez nagie zakończenia nerwowe.
Impulsacja bólowa przewodzona jest przez włókna aferentne z osłonką
mielinową, należące do grupy A, i bez osłonki mielinowej, należące do
grupy C d.r. W uszkodzonych tkankach dochodzi do aktywacji enzymów
proteolitycznych zwanych tkankowymi kalikreinami. Enzymy działają na
białka tkankowe — kininogeny, odczepiając od nich aktywne poli -
Czucie eksteroceptywne
1 63
Ryc. 89. Droga przewodząca impulsy z receptorów ciepła, zimna i bólu w skórze do pól
czuciowych kory mózgu.
1 64
Czucie i ruch
peptydy — kininy, które depolaryzują nagie zakończenia nerwowe i wyzwalają w aferentnych włóknach nerwowych salwy impulsów bólowych.
Kininy stanowią liczną grupę polipeptydów o zbliżonym fizjologicznym
działaniu. Spośród nich została poznana budowa chemiczna dziewięciopeptydu — bradykininy. Kininy nie tylko depolaryzują nagie zakończenia
nerwowe, lecz również rozszerzają naczynia krwionośne.
Pomiędzy bodźcem progowym i bodźcem maksymalnym wywołującym
czucie bólu zachodzi stosunek energii jak 1 :2. Maksymalne czucie bólu
występuje w czasie zadziałania energii tylko dwukrotnie większej od
progowej, co stanowi zabezpieczenie przed bodźcami uszkadzającymi
tkanki.
W ośrodkowym układzie nerwowym impulsacja bólowa biegnie dwiema drogami. Pierwsza — swoista — pokrywa się z drogą przewodzącą
impulsację od receptorów ciepła i zimna za pośrednictwem jąder w rogach tylnych rdzenia kręgowego (II neuron), jąder przekaźnikowych
wzgórza (III neuron) do kontralateralnego zakrętu zasrodkowego w korze
mózgu (IV neuron), czyli do pierwszej i drugiej korowej reprezentacji
czucia dotyku oraz do zakrętu obręczy (gyrus cinguli). Druga droga —
nieswoista — biegnie od II neuronu w rogach tylnych rdzenia poprzez
układ siatkowaty wstępujący, istotę szarą środkową śródmózgowia i jądra
śródblaszkowe wzgórza do wszystkich pól w korze mózgu.
Czucie eksteroceptywne
1 65
Hamowanie czucia bólu
Hamowanie przewodzenia impulsacji bólowej biegnącej od miejsca
zadziałania bodźca nocyceptywnego zachodzi przede wszystkim w obrębie
rogów tylnych rdzenia kręgowego oraz we wzgórzu i w układzie nieswoistym.
W rogach tylnych rdzenia kręgowego do II neuronu czuciowego przewodzą impulsację bólową włókna C o średnicy 0,4—1,2 um i włókna A
0 małej średnicy 2—5 um. Włókna A o większej średnicy 6—12 um
przewodzą natomiast impulsację od receptorów dotyku, która po wejściu
do rdzenia kręgowego wstępuje w sznurze tylnym do rdzenia pr ze
dłużonego i stąd do istoty szarej środkowej śródmózgowia (substantia
grisea centralis mesencephali). Włókna te mają również gałązki oboczne
kończące się w rogach tylnych rdzenia kręgowego na:
• zakończeniach włókien C, wywołując presynaptyczne hamowanie;
• neuronach pośredniczących hamujących.
Neurony istoty szarej środkowej śródmózgowia poprzez zstępujące
włókna pobudzają neurony pośredniczące hamujące w rogach tylnych
rdzenia kręgowego. Dzięki presynaptycznemu hamowaniu włókien C
1 pobudzeniu neuronów pośredniczących hamujących jest hamowane
przewodzenie impulsacji bólowej przez II neuron czuciowy.
Czucie bólu jest procesem złożonym. Jego wystąpienie warunkowane
jest obecnością aferentnej impulsacji bólowej i jednocześnie odpowiedniej
wrażliwości na tę impulsację ośrodków w rdzeniu kręgowym i przedłużonym, w śródmózgowiu, we wzgórzu, w podwzgórzu i w układzie
limbicznym. Neurony w tych ośrodkach, przewodzące impulsację bólową, mają w błonie komórkowej receptory wiążące się z transmitterami
pobudzającymi i hamującymi, jak również receptory opioidowe. Z tymi
receptorami wiążą się peptydy opioidowe, do których należą: enkefalina
metioninowa i leucynowa, beta-endorfina, alpha- i beta-neoendorfina,
dynorfina 1—8 i dynorfina 1—17 (patrz str. 81).
Działanie peptydów opioidowych nie ogranicza się do synaps, w obrębie których są uwalniane, ale poprzez płyn międzykomórkowy działają
na całe ośrodki i przechodzą do płynu mózgowo-rdzeniowego. Wykazano,
że u ludzi skarżących się na przewlekłe bóle zawartość peptydów opioidowych w płynie mózgowo-rdzeniowym jest mała.
Impulsacja bólowa przewodzona przez swoiste i nieswoiste drogi
i ośrodki w dłuższym czasie podlega wzmocnieniu lub tłumieniu dzięki
zmianom:
• w uwalnianiu transmitterów pobudzających i hamujących;
166
Czucie i ruch
Ryc. 91. Hamowanie czucia bólu w obrębie rogów tylnych rdzenia kręgowego. Neurony
czuciowe w rogach tylnych odbierają jednocześnie impulsację przekazywaną przez synapsy
pobudzające (jasne kółka) i przez synapsy hamujące (czarne kółka). Pobudzone receptory
skórne wysyłają impulsację: (1) — do pól czuciowych kory mózgu przez drogi wstępujące
swoiste (d.w.s.) i jądra przekaźnikowego wzgórza, (2) — do wszystkich pól w korze mózgu
przez drogi wstępujące nieswoiste (d.w.n.) i jądra nieswoiste wzgórza, (3) — do mięśni
poprzecznie prążkowanych i (4) — za pośrednictwem układu współczulnego do mięśni
gładkich. Impulsację bólową hamuje kora mózgu, istota szara środkowa śródmózgowia
i układ siatkowaty zstępujący poprzez neurony pośredniczące hamujące w rogach tylnych
rdzenia kręgowego, d.z. — droga zstępująca.
Czucie eksteroceptywne
1 67
• w uwalnianiu modulatorów synaptycznych, szczególnie peptydów opioidowych;
• w gęstości receptorów w błonach pre- i postsynaptycznych;
• wywołanym inkorporacją modu latorów do wnętrza neuronów, ich
wewnątrzkomórkowym transportem do perykarionów i ich oddziały
waniem na syntezę: transmitterów, modulatorów, receptorów błono
wych i enzymów (patrz tab. 3, str. 77 i tab. 4, str. 80).
I Czucie smaku
W jamie ustnej znajdują się skupione w kubkach smakowych (caliculi
gustatorii) receptory odbierające cztery podstawowe smaki: słodki, kwaśny, słony i gorzki.
Komórki receptorowe odbierające czucie smaku zaopatrzone są w wypustki w postaci mikrokosmków skierowanych do otworu smakowego
(porus gustatorius). Komórki smakowe zostają pobudzone przez substancje rozpuszczone w śluzie pokrywającym błonę śluzową w okolicy kubków smakowych i działające na mikrokosmki.
Ryc. 92. Schemat przekroju kubka
smakowego błony śluzowej języka.
(Reprodukowano za zezwoleniem
z W. Sawicki Histologia, II wyd.
Wydawnictwo Lekarskie PZWL,
Warszawa 1997.)
Substancje wywołujące czucie różnych smaków pobudzają te same
komórki smakowe, lecz w różnym stopniu. Pobudzenie komórek receptorowych zależy od wielkości cząsteczki substancji rozpuszczonej w śluzie.
Tym samym częstotliwość impulsów nerwowych we włóknach aferentnych zależy od rodzaju działającej substancji. Wrażliwość komórek
smakowych w kubkach rozmieszczonych w błonie śluzowej pokrywającej
język, krtań i gardło nie jest jednakowa.
1 68
Czucie i ruch
Dzięki różnorodnemu umiejscowieniu w błonie śluzowej kubków
smakowych i odmiennej ich wrażliwości na cząsteczki substancji rozpuszczonych w śluzie odbierane jest czucie czterech smaków. Próg pobudliwości komórek smakowych na jedną substancję jest różny i wynosi np. dla
wodnego roztworu chlorku sodowego od 2,71 do 43,1 mmol/L NaCl (od
0,016 do 0,25%).
Włókna nerwowe przewodzące impulsy z komórek smakowych biegną
w nerwach czaszkowych: twarzowym — VII (w strunie bębenkowej),
językowo-gardłowym — IX i błędnym — X. Pierwszy neuron czuciowy
znajduje się w zwojach tych nerwów, a drugi mieści się w jądrze samotnym
{nucleus solitarius) w rdzeniu przedłużonym. Aksony tych neuronów
przechodzą na drugą stronę rdzenia przedłużonego i — biegnąc we wstędze przyśrodkowej {lemniscus medialis) — kończą się na III neuronie
czuciowym w jądrze brzusznym tylno-przyśrodkowym wzgórza {nucleus
ventralis thalami posteromedialis). W korze mózgu, w zakręcie zaśrodkowym powyżej bruzdy bocznej, znajduje się IV neuron czuciowy. W tej
samej okolicy kory mózgu skupione są neurony czuciowe odbierające
pobudzenie od innych receptorów z okolicy jamy ustnej i gardła.
Ryc. 93. Rozmieszczenie receptorów w błonie
śluzowej języka odbierających różne wrażenie
smakowe.
I Czucie proprioceptywne
Proprioreceptory wysyłają informacje do ośrodkowego układu nerwowego o stanie układu kostno-stawowo-mięśniowego oraz ruchu całego
ciała.
Czucie proprioceptywne
1 69
Receptory układu ruchu
W układzie szkieletowym i mięśniowym znajdują się receptory pobudzane pracą statyczną i dynamiczną mięśni szkieletowych (kinesthetic
receptors). Znajdujące się tam receptory odbierają napięcie oraz rozciągnięcie mięśni i ścięgien, a także ucisk wywierany na powierzchnie
stawowe. Receptory te występują we wrzecionkach nerwowo-mięśniowych w postaci zakończeń pierścieniemato-spiralnych, ciałek zmysłowych
(Ruffiniego), w ścięgnach jako ciałka buła wko watę (Golgiego), na powierzchniach stawowych, zaś w okostnej jako ciałka blaszkowate (Paciniego)
oraz wolne zakończenia nerwowe.
Ruch ciała zarówno po prostej, jak i obrotowy jest odbierany przez
receptory znajdujące się w błędniku (labyrinthuś) stanowiącym narząd
równowagi.
Droga impulsów z układu ruchu
Impulsacja z proprioreceptorów układu ruchu jest odbierana przez
I neuron czuciowy znajdujący się w zwojach rdzeniowych, a następnie
jest przekazywana przez dwie różne drogi do kory mózgu i do kory
móżdżku.
Impulsacja przewodzona przez I neuron biegnie bezpośrednio do
mózgowia w pęczku smukłym (fasciculus gracilis) i pęczku klinowatym
(fasciculus cuneatus) do II neuronu w jądrze smukłym (nucleus
gracilis) i w jądrze klinowatym (nucleus cuneatus), a następnie przechodzi
na drugą stronę rdzenia przedłużonego i podąża we wstędze przyśrodkowej
(lemnis-cus medialis) do III neuronu w jądrze brzusznym tylno-bocznym
wzgórza (nucleus ventralis posterolateralis thalami). IV neuron czuciowy
znajduje się w zakręcie zaśrodkowym kory mózgu.
Część wypustek neuronów ze zwojów rdzeniowych (I neuron) kończy
się synapsami już w obrębie rogów tylnych rdzenia kręgowego na II
neuronie czuciowym. Aksony tych neuronów przechodzą na drugą stronę
rdzenia kręgowego, wstępując ku górze:
• w drodze rdzeniowo-wzgórzowej przedniej i bocznej (tractus spinothalamicus anterior et lateralis) do III neuronów w jądrze brzusznym
tylno-bocznym wzgórza, który z kolei wysyła wypustki do IV neuronu
w zakręcie zaśrodkowym kory mózgu;
• w drodze rdzeniowo-móżdżkowej przedniej i tylnej (tractus spinocerebellaris anterior et posterior) do kory móżdżku.
170
Czucie i ruch
W tych samych polach zakrętu zaśrodkowego kory mózgu znajdują się
neurony odbierające impulsację z proprioreceptorów układu ruchu oraz
eksteroreceptorów skóry.
W warunkach fizjologicznych nie jest odczuwana impulsacja z proprioreceptorów, która pozostaje poniżej progu świadomości. Jej występowanie można wykazać jedynie pośrednio. Mając zamknięte oczy,
człowiek bezbłędnie określa położenie kończyny, która nie dotyka niczego,
co mogłoby stanowić dodatkowe źródło informacji.
| Czynność błędnika
W błędniku znajdują się proprioreceptory narządu równowagi. Błędnik składa się z części kostnej — błędnika kostnego (labyrinthus osseus),
i leżącej wewnątrz części błoniastej — błędnika błoniastego (labyrinthus
membranaceuś). W skład błędnika wchodzą: przedsionek (vestibulum),
obejmujący łagiewkę (utriculus), i woreczek (sacculus) oraz trzy przewody półkoliste (ductus semicirculares).
Błędnik błoniasty wypełniony jest śródchłonką o dużej koncentracji
jonów K+ (150 mmol/L). W plamkach statycznych łagiewki (macula
utriculi) i woreczka (macula sacculi) oraz w grzebieniach bankowych
przewodów półkolistych (crista ampullae) znajdują się komórki nabłonka
walcowatego mające kosmki od strony śródchłonki. Są to komórki
zmysłowe włoskowate (cellulae sensoriae pilosa), czyli receptory narządu
równowagi (eąuilibrium receptors). Kosmki pokrywające powierzchnie
szczytowe tych komórek mają różną długość. Jeden najdłuższy kosmek jest
to kinocylium, kosmki krótsze zaś są to stereocylia.
W spoczynku komórka zmysłowa włoskowata stale uwalnia transmitter,
który wyzwala we włóknach nerwowych oplatających te receptory aferentne impulsy nerwowe o pewnej częstotliwości. Przechylenie się stereocyliów w stronę kinetocylium powoduje otwieranie kanałów dla dokomórkowego prądu jonów K+ w błonie komórkowej komórek włoskowatych.
Błona komórkowa depolaryzuje się i zwiększa się uwalnianie transmittera,
który wywołuje przyspieszenie częstotliwości aferentnych impulsów nerwowych we włóknach nerwowych stanowiących wypustki neuronów
zwoju przedsionkowego (ganglion vestibulare). Przechylenie się stereocyliów w przeciwną stronę — od kinetocylium — wywołuje zamykanie się
kanałów dla dokomórkowego prądu jonów K +, hiperpolaryzację błony
komórkowej, hamowanie uwalniania transmittera i zmniejszenie częstotliwości aferentnych impulsów nerwowych biegnących do zwoju przed-
Czucie proprioceptywne
171
sionkowego, a stąd do jąder przedsionkowych w moście {nuclei vestibulares
pontis).
W plamkach statycznych łagiewki i woreczka kosmki komórek zmysłowych włoskowatych są pokryte masą galaretowatą, czyli błoną kamyczkową {membrana statoconiorum). Na niej osadzone są kryształki soli
wapniowych zwane kamyczkami błędnikowymi {statoconia) lub otolitami.
Ruch liniowy głowy, przyspieszający lub zwalniający, powoduje przemieszczanie się kamyczków błędnikowych. Kosmki odchylają się w stronę
kinetocylium, zwiększa się wówczas częstotliwość impulsów nerwowych
biegnących do zwoju przedsionkowego, lub przechylają się w stronę
przeciwną i częstotliwość impulsów nerwowych zmniejsza się. Próg
pobudliwości na przyspieszenie liniowe (linear acceleration) dla komórek
receptorowych w plamkach przedsionka wynosi 12 cm/s.
Kamyki na błonie
statycznej
Ryc. 94. Schemat plamki przedsionka z komórkami receptorowymi: (1) w stanie spoczynku,
(2) zahamowanymi i (3) pobudzonymi. (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
W czasie przyspieszającego lub zwalniającego ruchu obrotowego głowy
następuje przepływ śródchłonki wewnątrz przewodów półkolistych i baniek błoniastych {ampullae membranaceae). Przepływ śródchłonki przez
bańkę odchyla osklepek {cupula) utworzony przez kosmki komórek
zmysłowych zlepione masą galaretowatą. Przechylanie się kosmków
172
Czucie i ruch
w stronę kinetocylium, a więc wtedy kiedy osklepek odchyla się w stronę
łagiewki, zwiększa częstotliwość impulsów w części włókien nerwowych
przewodzących do zwoju przedsionkowego. Przechylanie się kosmków ]
i osklepka w stronę przeciwną — od łagiewki — zmniejsza częstotliwość
impulsów we włóknach nerwowych.
Komórki zmysłowe włoskowate w grzebieniach bankowych reagują na
ruch obrotowy głowy przyspieszający, czyli na przyspieszenie kątowe
(angular acceleration). Bodźcem progowym jest przyspieszenie kątowe
równe 2—3° na sekundę.
Ruch obrotowy w prawo, czyli zgodnie z ruchem wskazówek zegara,
powoduje ruch śródchłonki w przewodach półkolistych przednich, w prawym przewodzie do łagiewki, w lewym przewodzie zaś od łagiewki.
W bańce prawego przewodu osklepek odchyla się w stronę łagiewki
i przyspiesza się częstotliwość impulsów we włóknach nerwowych biegnących do zwoju przedsionkowego. W tym samym czasie w bańce lewego
kanału osklepek odchyla się w stronę przeciwną od łagiewki i zwalnia się
częstotliwość impulsów biegnących do zwoju przedsionkowego. Neurony
zwojowe odbierają w tym samym czasie impulsację o różnym wzorze od
prawego i lewego błędnika.
Droga impulsów z błędnika
Włókna nerwowe oplatające komórki nabłonka zmysłowego przekazują pobudzenie do I neuronu czuciowego w zwoju przedsionkowym
(ganglion vestibulare). II neuron znajduje się w moście w jednym z czterech jąder przedsionkowych (nuclei vestibulares): górnym, dolnym, bocznym lub przy środkowym. Z tych jąder impulsacja jest wysyłana do:
• kory móżdżku przez konar dolny móżdżku (pedunculus cerebellaris
inferior);
• jąder ruchowych dla mięśni gałek ocznych — jąder nerwów czasz
kowych III, IV i VI za pośrednictwem pęczków podłużnych przyśrodkowych (fasciculi longitudinales mediales);
• rdzenia kręgowego przez drogę przedsionkowo-rdzeniową (tractus
vestibulospinalis).
Dzięki tym połączeniom zmiana położenia głowy w stosunku do
tułowia odebrana przez receptory błędnika wywołuje natychmiastową
odruchową korekcję napięcia mięśni kończyn i tułowia oraz odpowiednie
ustawienie gałek ocznych.
Czucie proprioceptywne
1 73
Ryc. 95. Drogi i ośrodki przewodzące
i odbierające impulsację nerwową biegnącą z błędnika: (1) zwój przedsionkowy,
(2) jądro przedsionkowe dolne, (3) jądro
przedsionkowe przyśrodkowe, (4) jądro
przedsionkowe górne, (5) jądro przedsionkowe boczne (Deitersa), (6) przez konar dolny móżdżku do kory móżdżku,
(7) przez drogę przedsionkowo-wzgórzową
do (8) wzgórza i (9) kory mózgu płata
ciemieniowego, (10) przez pęczki podłużne
przyśrodkowe do (11) jądra nerwu okoruchowego, (12) jądra nerwu bloczkowego,
(13) jądra nerwu odwodzącego i do rdzenia
kręgowego przez (14) drogę przedsionkowo-rdzeniową boczną i (15) drogę przedsionkowo-rdzeniową przyśrodkową.
Percepcja dotycząca ustawienia głowy w stosunku do tułowia jest
uzależniona od impulsacji nerwowej biegnącej od jąder przedsionkowych
poprzez jądra przekaźnikowe wzgórza do zakrętu zasrodkowego w korze
mózgu.
Oczopląs
Poza świadomością pozostaje kontrola ruchów gałek ocznych niezależnych od ruchów głowy. Na przykład w czasie obserwowania dowolnego przedmiotu z okna szybko jadącego pociągu występują ruchy gałek
ocznych o naprzemiennej fazie wolnej i szybkiej, które utrzymują obraz
tego przedmiotu na tym samym polu siatkówki. Takie ruchy gałek
ocznych noszą nazwę oczopląsu optokinetycznego (optokinetic nystagmus).
Ruchy gałek ocznych można również wywołać pobudzając receptory
błędnika. W tym celu stosuje się przyspieszenie (oczopląs podczasobro-
174
Czucie i ruch
towy) lub zwolnienie (oczopląs poobrotowy) obrotów głowy wraz z całym
ciałem u człowieka siedzącego na specjalnym fotelu. Ogrzewając lub
oziębiając błędnik, po wprowadzeniu ciepłej lub zimnej wody do zewnętrznego przewodu słuchowego, uzyskuje się oczopląs kaloryczny. Przyspieszenie lub zwolnienie obrotów głowy, jak również ciepło lub zimno,
wywołują ruch śródchłonki w przewodach półkolistych i pobudzenie lub
hamowanie receptorów błędnika.
I Czucie interoceptywne
Czucie trzewne (yisceral sensation) jest odbierane przez interoreceptory znajdujące się w narządach wewnętrznych, które pobudzane są
przez bodźce:
• mechaniczne, czyli wywołane rozciąganiem tkanek, w których się
znajdują np.: presoreceptory, baroreceptory w ścianach tętnic, mechanoreceptory w tkance płucnej;
• chemiczne, np.: prężność tlenu we krwi — chemoreceptory kłębuszka
szyjnego {glomus caroticum) i kłębka aortowego {glomus aorticum),
związki wytwarzane w niedokrwionych narządach wewnętrznych itp.
Impulsacja z interoreceptorów jest odbierana przez wypustki neuronów czuciowych (I neuron) w zwojach rdzeniowych. Wypustki tych
neuronów stanowią włókna aferentne zazwyczaj bez osłonki mielinowej
(C d.r.), biegnące od receptorów wraz z nerwami somatycznymi i autonomicznymi. Impulsacja z interoreceptorów jest przewodzona do mózgowia po stronie przeciwnej rdzenia kręgowego w sznurach tylnych,
bocznych i przednich. Neurony jąder brzusznych wzgórza tylno-bocznego
i tylno-przyśrodkowego przekazują impulsację do zakrętu zaśrodkowego
kory mózgu. Dzięki temu impulsacja z interoreceptorów nakłada się
w obrębie kory mózgu na impulsację z eksteroreceptorów i proprioreceptorów.
W warunkach prawidłowych impulsacja z interoreceptorów pozostaje
poniżej progu świadomości, jednak stale jest przewodzona przez trzewne
łuki odruchowe, uczestnicząc w regulacji czynności układu sercowo-naczyniowego, układu oddechowego, układu trawiennego i układu moczowo-płciowego.
Czucie interoceptywne
175
I Czucie bólu trzewnego
Proces chorobowy toczący się w obrębie jakiegoś narządu wewnętrznego wywołuje podrażnienie interoreceptorów. Impulsacja wywołana
przez interoreceptory nasila się poza fizjologiczną granicę częstotliwości.
Tym samym przez trzewny łuk odruchowy jest przewodzona zwiększona
w jednostce czasu liczba impulsów. Neurony czuciowe w rogach tylnych
rdzenia kręgowego stają się bardziej pobudliwe dzięki konwergencji —
przełączaniu się — impulsów z neuronów należących do trzewnych łuków
odruchowych na inne neurony przewodzące czucie z eksteroreceptorow
i proprioreceptorów.
Dzięki metamerycznemu, czyli odcinkowemu, unerwieniu skóry, mięśni
i narządów wewnętrznych impulsacja z interoreceptorów przełącza się
na drogi przewodzące czucie ze skóry i z mięśni unerwionych przez ten
sam odcinek (metamer) rdzenia kręgowego. Wywołuje to przeczulicę
skóry i promieniowanie bólu do okolic odległych od chorobowo zmienionego narządu wewnętrznego. Pojawiają się: promieniowanie trzewno-
Ryc. 96. Przełączanie się w rogach tylnych rdzenia kręgowego impulsacji biegnącej przez
trzewny łuk odruchowy na neurony przewodzące impulsację od eksteroreceptorow w skórze
i od propioreceptorów w mięśniach szkieletowych.
176
Czucie i ruch
Tabela 6
Drogi i ośrodki czuciowe swoiste
Czucie interoceptywne
\11
178
Czucie i ruch
-skórne (viscerocutaneous radiation), czyli przeczulica skóry, oraz odruchy trzewno-mięśniowe (visceromuscular reflex) — wzmożone napięcie
mięśni poprzecznie prążkowanych szkieletowych unerwionych przez ten
sam odcinek rdzenia kręgowego co narząd chorobowo zmieniony.
Aferentna impulsacja z interoreceptorów nie jest przekazywana wyłącznie przez włókna wstępujące do rdzenia kręgowego w obrębie jednego
czy dwóch sąsiednich metamerów. Może być przewodzona do rdzenia
również w obrębie kilku wyżej położonych metamerów.
Ryc. 97. Pola zakreskowane wyznaczają powierzchnię skóry objętej przeczulica i promieniowaniem bólu z narządów wewnętrznych:
mięśnia sercowego, żołądka i okrężnicy.
I
Drogi nieswoiste dla czucia
i percepcji
Drogę nieswoistą dla impulsów z receptorów stanowi wstępujący układ
siatkowaty (ascending reticular system). Włókna aferentne biegnące
w drogach swoistych wysyłają gałązki oboczne, czyli kolateralne, do
tworu siatkowatego pnia mózgowia (formatio reticularis trunci cerebri).
Przełączanie się impulsacji za pośrednictwem gałązek obocznych zachodzi
w większości przypadków pomiędzy II i III neuronem czuciowym dróg
swoistych. Twór siatkowaty pnia mózgowia przewodzi impulsy przez całą
sieć neuronów i synaps pomiędzy nimi. Wypustki neuronów tworu
Drogi nieswoiste dla czucia i percepcji
1 79
siatkowatego docierają do wszystkich pól kory mózgu oraz do ośrodków
podkorowych kontrolujących czynności poszczególnych narządów i układów oraz do ośrodków motywacyjnych kierujących zachowaniem się
człowieka.
Ryc. 98. Pola korowe według klasyfikacji Brodmanna. (Reprodukowano za zezwoleniem
z S. Różycki Anatomia mózgowia i rdzenia kręgowego, VI wyd. Państwowy Zakład
Wydawnictw Lekarskich, Warszawa 1960.)
1 80
Czucie i ruch
Warunkiem odebrania i rozpoznania bodźca działającego na receptory
jest jednoczesne pobudzenie:
• swoistych neuronów w polu czuciowym kory mózgu;
• całej kory mózgu;
• ośrodków podkorowych.
I Ruchy i postawa ciała
Ośrodki kontrolujące ruchy dowolne i postawę ciała znajdują się w:
• korze mózgu;
• jądrach podkorowych kresomózgowia;
• móżdżku.
W koordynacji czynności tych ośrodków z czynnością rdzenia kręgowego pośredniczy również twór siatkowaty pnia mózgowia.
W zakręcie zaśrodkowym (gyrus postcentralis), w polu korowym 1—3
według klasyfikacji Brodmanna, znajdują się pola czuciowe, odbierające
pobudzenie z ekstero-, proprio- i interoreceptorów. Impulsacja aferentna
biegnąca do zakrętu zaśrodkowego jest stąd przekazywana do zakrętu
przedśrodkowego {gyrus precentralis), czyli do pierwszorzędowej korowej
reprezentacji ruchu (pola 4), znajdującego się ku przodowi od bruzdy
środkowej (Rolanda) (sulcus centralis). Inne pola kory mózgu i ośrodki
podkorowe modulują pobudliwość neuronów ruchowych, znajdujących
się w pierwszorzędowej korowej reprezentacji ruchu. W wyniku tego
następuje torowanie lub hamowanie drogi dla eferentnych impulsów
nerwowych i w konsekwencji wyzwolenie lub zahamowanie określonego
ruchu.
W warstwie V kory mózgu zakrętu przedśrodkowego znajduje się
około 34 000 komórek nerwowych piramidalnych olbrzymich (Betza),
których aksony biegną do jąder ruchowych pnia mózgowia i rdzenia
kręgowego.
Komórki nerwowe piramidalne olbrzymie stanowią I neuron ruchowy
i są jednocześnie ogniwem w licznych łańcuchach neuronów. Neurony,
pomiędzy którymi krążą impulsy nerwowe, są to zarówno neurony
pośredniczące, znajdujące się w warstwie IV kory zakrętu przedśrodkowego, jak i neurony innych pól kory mózgu i ośrodków podkorowych.
Aksony komórek nerwowych piramidalnych olbrzymich mają największą średnicę — od 10 do 20 ^m. Stanowią jednak tylko około 1%
Ruchy i postawa ciała
1 81
włókien biegnących przez każdą drogę piramidową. Pozostałe 9% ma
średnicę od 5 do 10 um (najwięcej włókien), około 90%, ma jeszcze
mniejszą średnicę — od 1 do 4 um.
Korowe reprezentacje ruchu
W korze mózgu znajdują się ośrodki skupiające neurony dla poszczególnych mięśni szkieletowych po stronie przeciwnej ciała (kontralateralnej). Występują w zakręcie przedśrodkowym w pierwszorzędowej
korowej reprezentacji ruchu (primary motor area). Poczynając od szczeliny pomiędzy półkulami i kończąc w okolicy bruzdy bocznej mózgu
{sulcus lateralis cerebri), w zakręcie przedśrodkowym znajdują się kolejno
ośrodki dla ruchów: stopy, goleni, uda, tułowia, ramienia, przedramienia,
dłoni i palców, głowy, żuchwy, oka, warg, języka i gardła.
W korze mózgu poniżej pierwszorzędowej reprezentacji ruchu znajduje się pierwszorzędowe pole tłumiące ruchy po stronie kontralateralnej
ciała — PNMA (primary negative motor area).
af
ef
Ryc. 99. Schemat połączeń pomiędzy neuronami pośredniczącymi
i jedną komórką nerwową, piramidalną, olbrzymią (Betza) w korze
mózgu: af — akson aferetny, ef —
akson eferentny, o — synapsy pobudzające, • — synapsy hamujące.
1 82
Czucie i ruch
Dodatkowe pole czuciowo-ruchowe — SSMA (supplementary sensorimotor area) — zostało umiejscowione u ludzi po przyśrodkowej stronie
półkul mózgu, w korze mózgu części przyśrodkowej zakrętu czołowego
górnego (gyrus frontalis superior), płacika okołośrodkowego (lobulus
paracentralis), przedklinka (precuneus) i części zakrętu obręczy (gyrus
cynguli). To dodatkowe pole czuciowo-ruchowe wyzwala ruchy ciała
zintegrowane z impulsacją czuciową, szczególnie wzrokową, jak również
z impulsacją wysyłaną przez ośrodki motywacyjne. Ku przodowi od tego
pola znajduje się dodatkowe pole tłumiące ruchy ciała — SNMA (suplementary negative motor area).
Ryc. 100. Pola ruchowe i czuciowe w korze mózgu człowieka. Ml — pierwszorzędowa
korowa reprezentacja ruchu, Sl — pierwszorzędowa korowa reprezentacja czucia, PNMA —
pierwszorzędowe pola korowe hamujące ruchy, SNMA — dodatkowe pola korowe hamujące
ruchy, SSMA — dodatkowe pola korowe czuciowo-ruchowe. (Reprodukowane za zezwoleniem z S. H. Lim, D. S. Dinner, P. K. Pillay i wsp. Functional anatomy of the human
supplementary sensorimotor area: results of extraoperative electrical stimulation. Electroenceph. Clin. Neurophysiol., 1994, 91, 179—193.)
Ruchy i postawa ciała
1 83
Droga impulsów dla ruchu
Neurony ruchowe nadrzędne wysyłają wypustki biegnące przez istotę
białą półkul mózgu, torebkę wewnętrzną (capsula interna), konar mózgu
(pedunculus cerebri) i drogi: korowo-mostową (tractus corticopontinus),
korowo-opuszkową {tractus corticobulbaris) i korowo-rdzeniową boczną
oraz przednią {tractus corticospinalis lateralis et anterior).
Włókna przewodzące eferentną impulsację z kory mózgu biegną
w kolanie i odnodze tylnej torebki wewnętrznej. Najbardziej ku przodowi są położone włókna do jąder ruchowych III, IV, V, VI nerwów
czaszkowych. Następnie znajdują się włókna korowo-jądrowe do jąder
ruchowych VII, IX, X, XI i XII nerwów czaszkowych. Dalej ku tyłowi
biegną włókna drogi korowo-rdzeniowej do jąder ruchowych w rogach
przednich rdzenia kręgowego.
Włókna drogi korowo-jądrowej na odpowiednich poziomach krzyżują się, przechodząc na drugą stronę mózgowia. Włókna drogi korowo-rdzeniowej bocznej krzyżują się w piramidach (pyramis), a drogi korowordzeniowej przedniej w obrębie rdzenia kręgowego.
Prędkość przewodzenia impulsów w drodze korowo-rdzeniowej wynosi od 60 do 70 m/s. Impulsy wysyłane przez I neuron ruchowy nadrzędny w zakręcie przedśrodkowym docierają bezpośrednio do II neuronów ruchowych podrzędnych w jądrach ruchowych mostu, rdzenia
przedłużonego i rdzenia kręgowego. Część impulsów ma jednak do
pokonania neurony pośredniczące pomiędzy I i II neuronem ruchowym.
Przez drogi łączące neurony ruchowe I z II w czasie wykonywania ruchów
przebiegają salwy impulsów nerwowych. W zależności od częstotliwości
impulsów w salwie i częstotliwości występowania salw zachodzi torowanie
drogi dla impulsów ruchowych w obrębie pnia mózgowia i rdzenia
kręgowego.
Droga korowo-rdzeniowa boczna utworzona jest z 80% włókien nerwowych, droga korowo-rdzeniowa przednia zaś tylko z 20% włókien
nerwowych biegnących z kory mózgu do rdzenia kręgowego. Droga
korowo-rdzeniowa boczna jest drogą filogenetycznie młodszą i przewodzi impulsy nerwowe do II neuronów ruchowych podrzędnych unerwiających mięśnie dystalne kończyn. Dzięki impulsacji przewodzonej
przez te drogi możliwe są precyzyjne ruchy kończyn, na przykład ruchy
pisarskie.
Impulsy nerwowe przewodzone przez drogę korowo-rdzeniową przednią
napotykają na swej drodze neurony ruchowe pośredniczące w rogach
przednich rdzenia kręgowego. Po przejściu impulsacji ruchowej przez te
neurony zostają pobudzone II neurony ruchowe podrzędne unerwiające
1 84
Czucie i ruch
Ryc. 101. Przekrój poziomy przez prawą półkulę mózgu, poniżej spoidła wielkiego mózgu,
oraz prawą i lewą torebkę wewnętrzną z drogami ruchowymi (lewa torebka
wewnętrzna) i z drogami czuciowymi (prawa torebka wewnętrzna).
mięśnie tułowia i mięśnie proksymalne kończyn. Dzięki tym mięśniom
utrzymywana jest postawa wyprostna ciała. Utrzymanie odpowiedniej
pozycji i postawy ciała dzięki skurczom tych mięśni umożliwia wykonywanie precyzyjnych ruchów kończyn. Współdziałanie impulsacji biegnącej
przez obie drogi korowo-rdzeniowe warunkuje płynne i precyzyjne ruchy
dowolne (voluntary movement).
Ruchy i postawa ciała
1 85
I Czynność układu pozapiramidowego
Neurony kory mózgu wysyłają impulsy nerwowe do mięśni szkieletowych również drogą pośrednią, w czym biorą udział jądra podkorowe.
Aksony neuronów kory mózgu biegną do jąder podkorowych, gdzie
kończą się synapsami. Z kolei neurony jąder podkorowych wysyłają
swoje aksony, które — z pominięciem dróg korowo-rdzeniowych — docierają do drugich neuronów ruchowych w jądrach ruchowych.
Do układu pozapiramidowego zalicza się następujące struktury podkorowe: jądro ogoniaste {nucleus caudatus), jądro soczewkowate (nucleus
lentiformis) — składające się ze skorupy (putameń) i gałki bladej (globus
pallidus) — jądro brzuszne przednio-boczne wzgórza (nucleus ventralis
anterolateralis thalami), jądro niskowzgórzowe (nucleus subthalamicus),
istotę czarną (substantia nigra) i jądro czerwienne (nucleus ruber).
Zasadniczą funkcją układu pozapiramidowego jest współdziałanie w
wyzwalaniu ruchów dowolnych i regulowanie napięcia mięśni szkieletowych. Funkcje te układ pozapiramidowy spełnia dzięki:
• zwrotnej projekcji dającej odrzut impulsów z jąder podkorowych
z powrotem do kory mózgu;
• impulsacji bezpośredniej biegnącej poprzez drogi pozapiramidowe od
jąder podkorowych do jąder ruchowych rdzenia kręgowego;
• impulsacji wysyłanej przez jądra podkorowe do ją der ruchowych
rdzenia kręgowego za pośrednictwem neuronów tworu siatkowatego
pnia mózgowia i rdzenia kręgowego.
Zwrotna projekcja kierująca impulsy z powrotem do kory mózgu wiąże
się z trzema drogami:
• impulsy z kory mózgu po przejściu przez jądro ogoniaste, skorupę, gałkę
bladą i jądro brzuszne przednio-boczne wzgórza powracają do kory
mózgu i zmieniają pobudliwość jej neuronów;
• impulsy z kory mózgu krążą pomiędzy jądrami mostu (nuclei pontis),
korą móżdżku (cortex cerebelli), jądrem zębatym móżdżku (nucleus
dentałus), jądrem brzusznym przednio-bocznym wzgórza i korą mózgu;
• łańcuch neuronów jest zamknięty w jądrach podkorowych, impulsy nie
powracają do kory mózgu. Impulsy z kory mózgu biegną do jądra
ogoniastego i stąd do istoty czarnej, która ponownie kieruje impulsy
do jądra ogoniastego.
1 86
Czucie i ruch
Ryc. 102. Sprzężenia zwrotne pomiędzy
korą mózgu i ośrodkami podkorowymi
należącymi do układu pozapiramidowego.
Impulsy nerwowe przekazywane są pomiędzy korą mózgu, jądrem ogoniastym, istotą czarną śródmózgowia, jądrem brzuszno-bocznym wzgórza i powracają do kory
mózgu. Pomiędzy jądrem ogoniastym i istotą czarną śródmózgowia w przewodzeniu impulsów pobudzających pośredniczą
neurony SP-ergiczne, w przewodzeniu impulsów nerwowych wywierających działanie hamujące — neurony GABA-ergiczne.
Istota czarna śródmózgowia zwrotnie działa na neurony jądra ogoniastego za pośrednictwem neuronów dopaminergicznych.
Jądro ogoniaste wysyła do istoty czarnej impulsy wywołujące zarówno
pobudzenie za pośrednictwem neuronów peptydergicznych uwalniających
na swych zakończeniach Substancję P, jak również hamowanie przy udziale
neuronów GABA-ergicznych uwalniających na swych zakończeniach
kwas gamma-aminomasłowy. Neurony istoty czarnej zwrotnie oddziałują
hamująco na neurony jądra ogoniastego dzięki neuronom dopaminergicznym i dopaminie uwalnianej na zakończeniach tych neuronów w jądrze
ogoniastym.
Ruchy i postawa ciała
1 87
I Czynność móżdżku
Móżdżek odbiera informację wysyłaną przez wszystkie receptory całego
ciała, przetwarza je i gromadzi na ułamek sekundy, a następnie kontroluje
układ ruchowy. Móżdżek pełni funkcję dystrybutora siły skurczów mięśni
poprzecznie prążkowanych, umożliwiając poruszanie się człowieka, utrzymanie postawy wyprostnej i wykonywanie płynnych ruchów kończyn.
W korze móżdżku są reprezentowane wszystkie rodzaje receptorów,
zarówno receptory błędników, proprioreceptory, eksteroreceptory, jak
i telereceptory. Wykazano w niej dwie somatotopowe reprezentacje receptorów całego ciała. Pierwsza prezentacja proprioreceptorów i eksteroreceptorów znajduje się w korze górnej powierzchni móżdżku, druga
reprezentacja zaś w korze dolnej powierzchni móżdżku. Obie reprezentacje odbierają impulsy z receptorów znajdujących się po tej samej stronie
ciała. Pomiędzy pierwszą i drugą somatotopową reprezentacją występuje
reprezentacja wzroku i słuchu.
Receptory błędnika są reprezentowane w korze móżdżku starego
(archeocerebellum) po tej samej stronie ciała — ipsilateralnie — i po
przeciwnej stronie ciała — kontralateralnie.
Ryc. 103. Czynnościowe wzajemne powiązania pomiędzy korą prawej i lewej półkuli
mózgu z kontralateralną korą odpowiednich półkul móżdżku. Pierwszorzędowa
korowa reprezentacja ruchu lewej półkuli
mózgu jest czynnościowo powiązana z prawą półkulą móżdżku.
1 88
Czucie i ruch
Ryc. 104. Zasadnicze połączenie pomiędzy korą mózgu, korą móżdżku i rdzeniem kręgowym.
Z pola ruchowego w prawej półkuli mózgu biegną impulsy do kory móżdżku lewej półkuli
w drogach korowo-mostowej, korowo-opuszkowej i korowo-rdzeniowej za pośrednictwem
jądra czerwiennego, jąder mostu, jądra siatkowatego bocznego rdzenia przedłużonego i jądra
dolnego oliwki. Neurony gruszkowate kory móżdżku wysyłają impulsy hamujące do jąder
móżdżku łączących się z korą mózgu za pośrednictwem jądra brzuszno-bocznego wzgórza.
Na schemacie są pominięte połączenia móżdżku z jądrami przedsionka. Włókna kiciaste
(w.k.), włókna pnące (w.p.) i aksony neuronów gruszkowatych (a.n.g.) (Na podstawie
G. J. Allen, N. Tsukahara Cerebrocerebellar communication systems. Physiological Reviews,
1974, 54, 957—1006.)
Ruchy i postawa ciała
1 89
Impulsacja aferentna jest przewodzona do kory móżdżku przez:
• drogi szybko przewodzące, kończące się w postaci tzw. włókien kiciastych (mossy fiber);
• drogi przewodzące z opóźnieniem kilkunasto- i kilkudziesięciomilisekundowym, których zakończeniem są włókna pnące (climbing fiber).
Impulsacja przewodzona przez włókna kiciaste do kory móżdżku za
pośrednictwem małych i dużych neuronów ziarnistych i neuronów koszyczkowatych jest przekazywana do neuronów groszkowatych (komórek
Purkinjego). Neurony gruszkowate, mające duże rozgałęzione dendryty,
gromadzą informację biegnącą do kory móżdżku. Ilość nagromadzonej
informacji stale zmienia się pod wpływem hamującego oddziaływania
neuronów gwiaździstych i koszyczkowatych na neurony gruszkowate.
Pobudliwość neuronów gruszkowatych jest stale modulowana przez
impulsy aferentne biegnące z kory mózgu za pośrednictwem jąder przekaźnikowych mostu, z rdzenia kręgowego, z jąder przedsionkowych i z tworu
siatkowatego rdzenia przedłużonego.
Włókna pnące stanowią drugi rodzaj zakończeń dróg aferentnych
w korze móżdżku, przewodzą impulsy od jąder dolnych oliwki. W korze
móżdżku włókna pnące otaczają dendryty neuronów gruszkowatych,
pokrywając je licznymi synapsami. Dzięki nim są utworzone monosynaptyczne połączenia pomiędzy włóknami pnącymi przewodzącymi aferentna
impulsację do kory móżdżku i neuronami gruszkowatymi wraz z ich
aksonami, które przewodzą eferentną impulsację od kory móżdżku.
Impulsacja aferentna przewodzona z opóźnieniem przez włókna pnące
wywiera silne, pobudzające działanie i wyzwala wyładowania neuronów
gruszkowatych. Czynność zakończeń synaptycznych włókien pnących
można porównać do działania zastawek, które nagle uwalniają nagromadzoną informację w neuronach gruszkowatych.
W korze robaka móżdżku (vermis cerebelli) znajdują się neurony
gruszkowate odbierające impulsację wysyłaną przez receptory błędnika
i przez siatkówkę. Impulsacja z siatkówki przekazywana jest przez
włókna pnące z opóźnieniem kilkudziesięciu milisekund. Neurony gruszkowate konwergujące impulsację biegnącą z błędnika z impulsacja pochodzącą z siatkówki zapewniają odpowiednie rozłożenie napięcia mięśniowego, dzięki któremu możliwa jest stała obserwacja nieruchomego
przedmiotu mimo wykonywanego ruchu głowy.
Od kory móżdżku impulsy eferentne przewodzone są wyłącznie przez
aksony neuronów gruszkowatych, które biegną do jąder móżdżku i do
jąder przedsionkowych od kory móżdżku starego. Neurony gruszkowate
kory móżdżku wysyłają swe aksony do czterech symetrycznych jąder
1 90
Czucie i ruch
móżdżku: jądra zębatego {nucleus dentatus), jądra wierzchu (nucleus
fastigii), jądra kulkowatego {nucleus globosus) i jądra czopowatego (nucleus emboliformis). Na zakończeniach aksonów neuronów gruszkowatych
uwalnia się transmitter hamujący, czyli hiperpolaryzujący błonę komórkową neuronów jąder móżdżku i jąder przedsionkowych. Neurony jąder
móżdżku stale wysyłają eferentną impulsację pobudzającą do kory mózgu
i rdzenia kręgowego. Impulsacja pobudzająca jest modulowana przez
hamujące oddziaływanie kory móżdżku.
Impulsy eferentne z lewej półkuli móżdżku biegną do prawej półkuli
mózgu i przeciwnie — z prawej półkuli móżdżku do lewej półkuli mózgu.
Pola górnej powierzchni kory móżdżku, w której znajduje się pierwsza
somatotopowa reprezentacja receptorów całego ciała, pozostają w łączności z kontralateralną pierwszorzędową reprezentacją ruchową w korze
mózgu.
Aksony neuronów jąder móżdżku tworzą drogi aferentne biegnące
przez konary móżdżku do jąder przekaźnikowych. Są to: położone
kontralateralnie: jądro brzuszno-boczne wzgórza, jądro czerwienne i jądro
dolne oliwki, oraz położone ipsilateralnie jądro siatkowate boczne rdzenia przedłużonego. Aksony neuronów gruszkowatych kory móżdżku
starego, stosunkowo nieliczne u człowieka, w większości biegną bezpośrednio do jąder przedsionkowych, zwrotnie je hamując.
I Układ siatkowaty pnia mózgowia
Układ siatkowaty pnia mózgowia (brain stem reticular formation) jest
pojęciem czynnościowym. Neurony tego układu przewodzą impulsy
nerwowe w ośrodkowym układzie nerwowym poza swoistymi drogami
czuciowymi i ruchowymi. Czynność tego układu warunkuje wzbudzenie
(arousal) i uwagę (attention) oraz kontrolę czynności somatycznych i autonomicznych.
Do podłoża anatomicznego tego układu zalicza się skupienia komórek
nerwowych występujących w tworze siatkowatym mostu, rdzenia przedłużonego i kręgowego, w tym jądra szwu {nuclei raphes) i jądro miejsca
sinawego {nucleus loci cerulei), jądra siatkowate śródmózgowia, istotę
szarą środkową śródmózgowia {substantia grisea centralis), jądro siatkowate wzgórza (nucleus reticularis thalami), jądra śródblaszkowe wzgórz
(nuclei intralaminares thalami) i jądro brzuszne przednio-boczne wzgórza
(nucleus ventralis anterolateralis thalami).
Układ siatkowaty tworzą różnorodne neurony przekazujące pobudzenie lub hamowanie za pośrednictwem większości dotychczas poznanych
Ruchy i postawa ciała
191
transmitterów i modulatorów zarówno pobudzających, jak i hamujących.
Neurony układu siatkowatego tym się charakteryzują, że ich aksony
rozdzielają się na gałązki wstępujące i zstępujące o licznych rozgałęzieniach. Dzięki tym rozgałęzionym wypustkom układ siatkowaty przekazuje impulsy licznym komórkom nerwowym, położonym zarówno
w wyższych, jak i w niższych piętrach ośrodkowego układu nerwowego.
Układ siatkowaty dzieli się na:
• zstępujący, w którym wyróżnia się część pobudzającą i hamującą;
• wstępujący, który wywiera wpływ pobudzający i hamujący (ukła d
rekrutujący wzgórza).
Układ siatkowaty zstępujący
Układ siatkowaty zstępujący (descending reticular formation) kontroluje czynność odruchową rdzenia kręgowego, napięcie mięśni szkieletowych, czynność ośrodków kontrolujących krążenie i oddychanie.
W zależności od wpływu, jaki wywierają neurony układu siatkowatego
zstępującego dzieli się go na:
• część hamującą (reticular inhibitory area), zwalniającą lub bloku
jącą czynność innych neuronów;
• część pobudzającą (reticular facilitatory area), torującą lub przyspie
szającą czynność neuronów rdzenia przedłużonego i rdzenia kręgo
wego.
Układ siatkowaty wstępujący
Układ siatkowaty wstępujący kontroluje czynność neuronów jąder
ruchowych mostu, jąder ruchowych mięśni gałek ocznych oraz przewodzi impulsy aferentne od różnych receptorów i tym samym stanowi dla
nich drogę nieswoistą.
Układ siatkowaty wstępujący pobudzający — RAS (reticular activating system) — przewodzi impulsy pobudzające do:
• wszystkich pól kory mózgu;
• ośrodków podkorowych kierujących zachowaniem, czyli ośrodków
motywacyjnych;
• ośrodków kontrolujących układ autonomiczny i gruczoły dokrewne.
Na przedłużeniu tworu siatkowatego pnia mózgowia znajdują się jądra
wzgórza skupiające neurony o działaniu antagonistycznym w stosunku
1 92
Czucie i ruch
do neuronów układu siatkowatego wstępującego, pobudzającego. Jądra
te tworzą układ rekrutujący (recruting system) synchronizujący czynność
bioelektryczną kory mózgu, a tym samym zwalniający czynność bioelektryczną jej neuronów. Układ rekrutujący obejmuje jądra wzgórza:
przyśrodkowe {nuclei mediales thalami), śródblaszkowe {nuclei intralaminares thalami), brzuszne przednio-boczne {nucleus ventralis anterolateralis thalami), przednio-przyśrodkowe {nucleus ventralis anteromedialis
thalami) oraz siatkowate wzgórza {nucleus reticularis thalami).
Neurony uwalniające różnorodne transmittery i
modulatory
Neurony cholinergiczne, syntetyzujące acetylocholinę (ACh) i uwalniające ją na swych zakończeniach, występują we wszystkich strukturach
ośrodkowego układu nerwowego. Znajdują się w rdzeniu kręgowym,
w korze mózgu, w hipokampie, w prążkowiu i w podwzgórzu. Na
zakończeniach aksonów neuronów ruchowych, w obrębie synaps nerwowo-mięśniowych, uwalnia się ACh. Układ siatkowaty wstępujący
pobudzający (RAS) tworzą również neurony cholinergiczne.
Neurony noradrenergiczne, ich aksony i zakończenia synaptyczne,
znajdują się w pniu mózgowia, w podwzgórzu, we wzgórzu, w korze
móżdżku, w opuszkach węchowych, w hipokampie i w korze mózgu.
Wykazano dwa skupienia neuronów noradrenergicznych, których aksony
tworzą dwa wstępujące pęczki. W jądrze miejsca sinawego {nucleus loci
cerulei), pod dnem IV komory mózgu, znajduje się skupienie ciał neuronów, których aksony tworzą pęczek grzbietowy noradrenergiczny (noradrenergic dorsal bundle). Aksony tego pęczka kierują się do kory
mózgu i do hipokampa. Ciała neuronów, których aksony tworzą pęczek
brzuszny noradrenergiczny (noradrenergic ventral bundle), skupione są w
kilku okolicach pnia mózgowia. Zakończenia aksonów pęczka
brzusznego znajdują się w rdzeniu przedłużonym, w moście, w
śródmózgowiu i w mię-dzymózgowiu.
Neurony noradrenergiczne, których ciała znajdują się w jądrze grzbietowym nerwu błędnego i w jądrze samotnym wysyłają swoje aksony
do rdzenia kręgowego. Neurony adrenergiczne występujące w rdzeniu
przedłużonym wysyłają wstępujące aksony do podwzgórza i zstępujące
do rdzenia kręgowego. Transmisja noradrenergiczna i adrenergiczna
działa pobudzająco na neurony w ośrodkach motywacyjnych w międzymózgowiu i na neurony peptydergiczne w podwzgórzu, hamując jednak
Ruchy i postawa ciała
193
Ryc. 105. Układ noradrenergiczny mózgowia składający się z układu noradrenergicznego:
a — mostu; b — rdzenia przedłużonego. Neurony układu noradrenergicznego mostu (A) mają
projekcję zstępującą do: rdzenia kręgowego i móżdżku oraz projekcję wstępującą do: blaszki
pokrywy, wzgórza, przegrody, kory mózgu, hipokampa, ciała migdałowatego i opuszki
węchowej. Neurony układu noradrenergicznego rdzenia przedłużonego (B) mają tylko
wstępującą projekcję do: wzgórza, przegrody, podwzgórza, okolicy przedwzrokowej i ciała
migdałowatego.
7 — Fizjologia człowieka
1 94
Czucie i ruch
w podwzgórzu neurony wazopresynoergiczne i oksytocynoergiczne, w ko- ]
rze móżdżku zaś neurony gruszkowate.
Ciała neuronów dopaminergicznych znajdują się w śródmózgowiu
w jądrze międzykonarowym {nucleus interpeduncularis), w istocie czarnej
{substantia nigra) i w podwzgórzu. Aksony neuronów dopaminergicznych
śródmózgowia wstępują w znacznej przewadze do jąder kresomózgowia,
kończąc się w prążkowiu, w guzkach węchowych, w jądrze wtrąconym
{nucleus accumbens) i w jądrze środkowym ciała migdałowego {nucleus
centralis corporis amagdaloidei).
Neurony dopaminergiczne występujące w obrębie podwzgórza zawierają również neurotensynę (NT). Ich aksony biegną do:
• naczyń włosowatych sieci pierwotnej przysadkowych naczyń wrotnych.
Dopamina uwolniona do krwi jest przenoszona do przedniego płata
przysadki, działając hamująco na wydzielanie prolaktyny (PRL) przez
komórki laktotropowe;
• części pośredniej przysadki, hamując wydzielanie hormonu melanotropowego (MSH) przez komórki części pośredniej;
• tylnego płata przysadki, modulując uwalnianie wazopresyny (AVP)
i oksytocyny (OXY) z aksonów neuronów wazopresynoergicznych
i oksytocynoergicznych;
• neuronów peptydergicznych podwzgórza, wytwarzających podwzgórzowe hormony, modulując ich wydzielanie.
Ciała neuronów serotoninergicznych w większości znajdują się w linii
środkowej pnia mózgowia, w jądrach szwu {nuclei raphe). Aksony tych
neuronów wstępują do prążkowia, kory mózgu, kory móżdżku, ciała
migdałowatego, wzgórza i podwzgórza, zstępujące aksony biegną zaś
do rdzenia kręgowego.
W neuronach serotoninergicznych występuje również Substancja P.
Działanie neuronów serotoninergicznych na inne neurony prawdopodobnie zależy od wzajemnego stosunku uwalnianych jednocześnie cząsteczek serotoniny i Substancji P. Neurony serotoninergiczne jąder szwu
uczestniczą w procesach związanych z utrzymaniem stanu czuwania
i stanu snu.
Sole kwasów glutaminowego i asparaginowego są głównymi transmitterami w drogach czuciowych i ruchowych. Działają dwufazowo — początkowo pobudzają błonę postsynaptyczną, a następnie działają hamująco.
Transmitterem hamującym, który uwolniony na synapsach hiperpolaryzuje błonę postsynaptyczną, jest kwas gamma-aminomasłowy (GABA).
Jest on syntetyzowany z kwasu glutaminowego, jak również wychwyty-
Ruchy i postawa ciała
1 95
wany z płynu zewnątrzkomórkowego przez komórki nerwowe. Neurony
zawierające GABA, czyli neurony GABA-ergiczne, znajdują się we
wszystkich strukturach ośrodkowego układu nerwowego. Najwięcej występuje ich w korze mózgu, korze móżdżku, prążkowiu, gałce bladej,
opuszce węchowej, we wzgórzu i w istocie szarej rdzenia kręgowego,
szczególnie w rogach tylnych. Neurony gruszkowate kory móżdżku są
to również neurony GABA-ergiczne i uwalniając GABA na zakończeniach swych aksonów, hamują czynność neuronów jąder przedsionkowych w moście. Aksony neuronów GABA-ergicznych prążkowia i gałki
bladej biegną do istoty czarnej, hamując aktywność neuronów dopaminergicznych skupionych w tej strukturze.
Neurony GABA-ergiczne i glicynoergiczne, znajdujące się w istocie
szarej rdzenia kręgowego, hamują przewodzenie aferentnych impulsów
nerwowych przez neurony czuciowe w obrębie rogów tylnych rdzenia
kręgowego oraz uwalniane przez neurony pośredniczące działają hamująco na neurony ruchowe w rogach przednich rdzenia kręgowego. Neurony pnia mózgowia należące do układu siatkowatego hamującego są
neuronami glicynoergicznymi.
W komórkach nerwowych występują jednocześnie dwa i więcej aktywnych biologicznie związków (coexistence of neuronal messengers), za
pośrednictwem których przenoszona jest informacja. Są to neurony
zawierające:
• transmitter i neuropeptyd;
• dwa transmittery;
• transmitter, neuropeptyd i adenozynotrifosforany (transmitter purynergiczny);
• dwa i więcej neuropeptydów;
• neuropeptyd i leukotrieny.
Większość modulatorów synaptycznych to neuropeptydy występujące
w neuronach w podwzgórzu (patrz tab. 4, str. 80). Są one wydzielane
na synapsach wraz z różnymi transmitterami, działając modulująco na
błonę postsynaptyczną i presynaptyczną innych neuronów, lub są wydzielane do naczyń włosowatych sieci pierwotnej przysadkowych naczyń
wrotnych, stanowiąc podwzgórzowe hormony (patrz tab. 10, str. 228).
Poza
podwzgórzem
znajdują
się
skupienia
komórek
peptydergicznych i pęczki aksonów peptydergicznych, w których
występuje jeden lub jednocześnie kilka neuropeptydów. Mają one
działanie modulujące, tj. wzmacniające lub tłumiące, w zależności od
tego, z jakimi transmitterami i modulatorami występują jednocześnie w
neuronach.
nodulatorów w komórkach nerwowych
Czucie i ruch
)
Tabela 7
Koegzystencja przekaźników (transmitterów i
i w ośrodkowym układzie nerwowym
1 96
11
28
Galanina
Peptyd pochodny kalcytoninowego genu (CGRP)
30
Cholecystokinina-8
(CCK-8)
Neurotensyna (NT)
Neuropeptyd Y (NPY)
Wazopresyna
argininowa (AVP)
Tyreoliberyna (TRH)
Substancja P i
tyreoliberyna
Motylina
Somatostatyna (SRIF)
Peptydy opioidowe
37
+
+
+
+
+
+
13
+
+
+
3
11+3
22
14
5 + 31
+
+
8
36
9
glicyna
+
+
kwas gamma-aminomasłowy
noradrenalina
dopamina
+
+
+
serotonina
5
Substancja P (SP)
Wazoaktywny peptyd
jelitowy (VIP)
Transmittery
adrenalina
Enkefaliny
Liczba
aminokwasów w
cząsteczce
acetylocholina
Neuropeptydy
— modulatory
+
+
+
+
+
+
+
+
+
+
+
Grupa neuropeptydów opioidowych ma działanie tłumiące. Są to neuropeptydy takie, jak enkefalina metioninowa lub enkefalina leucynowa,
zbudowane z pięciu aminokwasów, lub też neuropeptydy opioidowe
o dłuższym łańcuchu aminokwasowym, w których w N-końcu łańcucha
peptydowego występuje ta sama sekwencja aminokwasów jak w enkefalinie metioninowej (beta-endorfina) lub enkefalinie leucynowej (alpha-neoendorfina, beta-neoendorfina, dynorfina 1—8 i dynorfina 1—17).
Czuwanie i sen
1 97
Modulatory wychwytywane przez zakończenia aksonów mogą być
wstecznie transportowane do ciał neuronów, wpływając na aktywność
enzymów w perykarionie, lub po wniknięciu do jądra komórkowego
mogą oddziaływać na syntezę matrycowego RNA i na potranskrypcyjną
modyfikację mRNA.
I Czuwanie i sen
U ludzi dorosłych występują cyklinicznie w ciągu doby dwa podstawowe stany fizjologiczne, tj. czuwanie i sen. Około 2/3 doby przypada
na czuwanie i około 1/3 na sen. Czuwanie (wakefulness) jest to stan
aktywności układu somatycznego. Sen (sleep) natomiast jest przede
wszystkim spoczynkiem dla tego układu. W czasie snu dochodzi również
do zmian w czynności innych układów: sercowo-naczyniowego, oddechowego, trawiennego i gruczołów dokrewnych.
Występowanie w ciągu doby jednego okresu czuwania i jednego
okresu snu wiąże się z dojrzewaniem mózgowia. Noworodek śpi około
16 godzin na dobę, budząc się w ciągu doby od kilku do kilkunastu
razy. Wraz ze wzrostem organizmu i dojrzewaniem mózgowia czas snu
skraca się, czas czuwania zaś wydłuża. Tworzą się dwie fazy aktywności układu somatycznego, faza snu i faza czuwania.
W procesie snu i czuwania uczestniczą substancje chemiczne, tak
zwane czynniki snu (sleep factors), których jest ponad dwadzieścia, i kilka
czynników czuwania (waking factors). Do czynników snu i czuwania zalicza się substancje chemiczne powstające zarówno w mózgowiu, jak
również na obwodzie i wraz z krwią dopływające do mózgowia.
Do czynników snu należy melatonina wydzielana w ciemności z szyszynki do krwi (patrz str. 263), jak również kwas gamma-aminomasłowy (GABA), transmitter interneuronów hamujących, i interleukina 1
(IL-1) wytwarzana przez komórki glejowe w mózgowiu. W procesie
zasypiania oddziaływanie czynników snu przeważa nad czynnikami czuwania, następuje tłumienie aferentnej impulsacji przewodzonej przez
układ siatkowaty wstępujący pobudzający — RAS (reticular activating
system) (patrz str. 191) i człowiek zasypia. W czasie 7—8-godzinnego
snu nocnego zostaje zmetabolizowany nadmiar czynników snu w mózgowiu i poza nim — człowiek budzi się.
Wykazano zależności pomiędzy regulacją temperatury ciała i snem.
We śnie temperatura ciała stopniowo obniża się o około 0,6°C, aby przed
przebudzeniem wzrosnąć.
1 98
Czucie i ruch
*v> v *yrf* w *^wv^^
Ryc. 106. Elektroencefalogram człowieka: a — fale alpha (1), blokowanie fal alpha po
otwarciu oczu (2); b — fale beta (1); c — czuwanie (w), stadia snu NREM (1, 2, 3),
REM — faza snu o szybkich ruchach gałek ocznych. (Zapisy otrzymane od Hanny
Wilczak-Szadkowskiej z Pracowni EEG i Neurofizjologii Kliniki Psychiatr ycznej AM
w Warszawie.)
Czynność bioelektryczna mózgu charakterystyczna dla stanu czuwania
zmienia się po zaśnięciu, jak również w czasie trwania snu nocnego.
Podczas snu nocnego czynność bioelektryczna mózgu zmienia się w sposób
typowy. Pozwala to podzielić sen na fazy i stadia.
Czuwanie i sen
1 99
Czuwanie — W (wakefulness) — charakteryzuje się występowaniem
w EEG fal alpha i beta u ludzi pozostających w spoczynku i z zamkniętymi powiekami. Sen (sleep) dzieli się na dwie fazy: na sen o wolnych
ruchach gałek ocznych (non rapid eye movement sleep — NREM) i na sen
o szybkich ruchach gałek ocznych (rapid eye movement sleep — REM).
W czasie zasypiania, w pierwszym stadium snu o wolnych ruchach gałek
ocznych (lst stage NREM) zanikają fale alpha, pojawiają się fale o małej
amplitudzie i częstotliwości od 6 do 7 Hz. Stadium 1 NREM nie przekracza 5% całego czasu przypadającego na sen nocny.
W drugim stadium snu o wolnych ruchach gałek ocznych (2nd stage
NREM) amplituda fal nieco podwyższa się i od czasu do czasu pojawiają
się skupienia ostrych fal o częstotliwości od 12 do 14 Hz i ampli tudzie do 50 |iV tworzące tzw. wrzeciona senne (sleep spindles), zwane
również falami sigma, oraz zespoły fali ostrej z falą wolną, tzw. zespoły K.
Stadium 2 NREM zajmuje łącznie ponad połowę, to jest 50—55% czasu
przypadającego na sen nocny.
W trzecim stadium snu o wolnych ruchach gałek ocznych (3rd stage
NREM) występują fale o wyższej amplitudzie niż 75 uV i o częstotliwości poniżej 2 Hz. Fale te zajmują do 50% analizowanego elektroencefalogramu.
W czwartym stadium snu o wolnych ruchach gałek ocznych (4th stage
NREM) fale o wyższej amplitudzie niż 75 (iV i o częstotliwości poniżej
2 Hz zajmują ponad 50% analizowanego elektroencefalogramu. Stadium
3 i 4 NREM, nazywane również snem delta, dominuje w pierwszych
godzinach snu.
W czasie 8-godzinnego snu występuje kilka razy, zazwyczaj od 4 do 6,
faza snu o szybkich ruchach gałek ocznych — faza REM. Każdorazowo
faza REM trwa kilkanaście do kilkudziesięciu minut. Szybkim ruchom
gałek ocznych towarzyszą zmniejszone napięcie mięśni szkieletowych,
ruchy żuchwy i desynchronizacja czynności bioelektrycznej kory mózgu.
Faza snu REM zajmuje łącznie 20— 30% całkowitego czasu snu nocnego — TSR (total stage REM).
Na sen o wolnych ruchach gałek ocznych i synchronicznej czynności
bioelektrycznej (NREM) przypada od 70 do 80% całkowitego czasu
snu nocnego — TSN. W czasie tej fazy snu znacznie zmniejsza się
aktywność bioelektryczna neuronów adrenergicznych, serotoninoergicznych i histaminoergicznych w obrębie pnia mózgowia. Umożliwia to
krążenie impulsów nerwowych pomiędzy wzgórzem i korą mózgu i warunkuje pojawienie się w EEG fal o małej częstotliwości — czyli snu
wolnofalowego (slow wave sleep). Sen ten wiąże się z występowaniem
jednego z czynników snu w płynie mózgowo-rdzeniowym wypełniającym
200
Czucie i ruch
komory mózgu — peptydu wywołującego sen delta (delta sleep inducing
peptide).
Faza snu REM związana jest z aktywnością neuronów mostu. W czasie
tej fazy snu, poza desynchronizacją czynności bioelektrycznej mózgu,
stwierdzono największy przepływ krwi przez jego tętnice środkowe,
przyspieszenie częstości skurczów serca i inne objawy wyzwalane za
pośrednictwem autonomicznego układu nerwowego, choć aktywność
neuronów adrenergicznych, serotoninoergicznych i histaminoergicznych
jest całkowicie stłumiona.
Ryc. 107. Hipnogram otrzymany na podstawie analizy elektroencefalogramu w czasie snu
trwającego 8 godzin. W — czuwanie, REM — pola zakratkowane, 1, 2, 3 i 4 — stadia snu
NREM, 0 — początek snu.
Niezwykłość fazy REM spowodowało pojawienie się wielu hipotez,
którymi próbowano objaśnić fizjologiczną rolę tej fazy snu. Od fazy
REM miała zależeć konsolidacja pamięci, wybiórcze zapominanie, inteligencja i marzenia senne. Hipotezy nie zostały doświadczalnie potwierdzone. Na przykład człowiek obudzony zarówno w fazie REM, jak
i NREM relacjonuje marzenia senne.
Ośrodki kierujące zachowaniem
201
I Ośrodki kierujące zachowaniem
Ośrodki motywacyjne
W organizmach zwierzęcych, a więc również w organizmie człowieka,
w rozwoju filogenetycznym wykształciły się dwa podstawowe mechanizmy nerwowe kierujące ich aktywnością somatyczną. Są to:
• mechanizm unikania (avoidance system), który zabezpiecza organizm
przed działaniem czynników lub bodźców szkodliwych;
• mechanizm konsumowania (approach system), który tak kieruje za
chowaniem, aby potrzeby organizmu były zaspokojone.
Pojawienie się pobudki (motivation) prowadzi do zachwiania równowagi pomiędzy mechanizmem unikania i mechanizmem konsumowania.
Pobudzenie odpowiedniego ośrodka wyzwala aktywność somatyczną,
której wynikiem jest przywrócenie równowagi pomiędzy mechanizmem
unikania i konsumowania.
Do ośrodków wyzwalających aktywność somatyczną zalicza się:
ośrodek głodu (hunger center), ośrodek pragnienia (thirst center), ośrodek agresji (attack center), ośrodek ucieczki (fear center) oraz ośrodki
rozrodczy i macierzyństwa (sexual and maternal behavior centers).
Ośrodki kierujące zachowaniem znajdują się w międzymózgowiu.
Neurony należące do poszczególnych ośrodków są skupione lub rozsiane
na większych obszarach, sąsiadujących z neuronami różnych ośrodków.
Neurony te wykazują swoistą wrażliwość na zmiany w koncentracji
niektórych składników krwi, np. glukozy, NaCl i hormonów.
Ośrodek pokarmowy
Ośrodek pokarmowy (feeding center) obejmuje dwa antagonistyczne
ośrodki:
• ośrodek głodu (hunger center) znajdujący się w bocznej części podwzgórza;
• ośrodek sytości (satiety center) znajdujący się w jądrze brzuszno-przyśrodkowym podwzgórza.
W skład ośrodka głodu i ośrodka sytości wchodzą neurony wrażliwe
na zawartość glukozy we krwi, spełniające funkcję detektorów. Apetyt
zostaje pobudzony po zmniejszeniu się stężenia glukozy we krwi i po
202
Czucie i ruch
zwiększeniu stężenia insuliny we krwi obwodowej wydzielonej przez
komórki B wysp trzustkowych. Peptydy opioidowe działają pobudzająco
na ośrodek głodu. Zwiększenie stężenia cholecystokininy we krwi hamuje
apetyt.
Ośrodki pokarmowe w podwzgórzu kierują zachowaniem organizmu
tak, aby było zaspokojone jego zapotrzebowanie na energię i inne związki
niezbędne do życia, oraz kontrolują wydzielanie soku żołądkowego.
Ośrodek pragnienia
W części środkowej podwzgórza skupione są neurony ośrodka pragnienia wyzwalające aktywność ruchową związaną z poszukiwaniem i piciem wody. Ośrodek ten jest ściśle związany z ośrodkiem termoregulacji
w przedniej części podwzgórza i z neuronami jądra nadwzrokowego
(nucleus supraopticus hypothalami) syntetyzującymi wazopresynę. Wzrost
temperatury krwi lub wzrost ciśnienia osmotycznego krwi prowadzi do
wzmożonego pragnienia i picia wody.
T
e
m
p
e
r
a
f
u
r
a
k
rwi dopływającej ao podwzgórza
podwyższona
1
Ośrodek
głodu
zahamowany
Ośrodek
sytości
pobudzony
Ośrodek
picia
pobudzony
Ryc. 108. Schemat zależności pomiędzy ośrodkiem termoregulacji i ośrodkami głodu, sytości
i pragnienia.
Ośrodki agresji i ucieczki
W podwzgórzu znajdują się skupienia neuronów wyzwalające silne
reakcje somatyczne związane z mechanizmem konsumowania, reprezentowanym przez ośrodek agresji, oraz mechanizmem unikania reprezentowanym przez ośrodek ucieczki. Neurony, z których składają się te
dwa ośrodki, nie mają ścisłego umiejscowienia, lecz są rozsiane prawie
w całym podwzgórzu i w sąsiadujących z nim okolicach
śródmózgowia i wzgórza.
Ośrodki kierujące zachowaniem
203
Ośrodek rozrodczy i ośrodek macierzyństwa
Ośrodek rozrodczy kieruje aktywnością somatyczną organizmu związaną z zachowaniem gatunku. Czynność tego ośrodka jest odmienna
u płci męskiej i żeńskiej. Różnicowanie się w kierunku ośrodka męskiego
lub żeńskiego zachodzi u płodów ludzkich już w czasie życia wewnątrz
macicy w organizmie matki. Pod wpływem krążących we krwi androgenów wykształca się męski ośrodek rozrodczy.
Ośrodek męski po osiągnięciu dojrzałości płciowej przez organizm
wyzwala, pod wpływem hormonów płciowych, męski popęd płciowy
w kierunku płci żeńskiej i powoduje stałe, niecykliczne, wytwarzanie
podwzgórzowego hormonu uwalniającego z części gruczołowej przysadki
hormony gonadotropowe — Gn-RH (gonadotropin-releasing hormone).
Ośrodek żeński pod wpływem hormonów płciowych żeńskich wyzwala
popęd płciowy w kierunku płci męskiej i jednocześnie kontroluje cykliczne
wydzielanie podwzgórzowego hormonu uwalniającego hormony gonadotropowe z przysadki — Gn-RH.
I Układ limbiczny
Układ limbiczny, czyli rąbkowy (limbie system), obejmuje kilka struktur: hipokamp (hippocampus), ciało migdałowate (corpus amygdaloideum), przegrodę przezroczystą (septum pellucidum) oraz drogi je łączące:
sklepienie (fornix), prążek krańcowy {stria terminalis) i inne. Do tego
układu zalicza się również filogenetycznie najstarsze części kory mózgu:
zakręt obręczy (gyrus cinguli) oraz sąsiadujące ze sobą pola wyspy
(insula), płat skroniowy po stronie przyśrodkowej i zakręty oczodołowe
{gyri orbitales).
Układ rąbkowy kontroluje ośrodki podwzgórza zarówno związane
z aktywnością somatyczną, jak i autonomiczną oraz wewnątrzwydzielniczą.
Struktury należące do układu limbicznego mają połączenia z korą
mózgu. Jądra ciała migdałowatego łączą się bezpośrednio z polami
w korze mózgu swoistymi dla różnych rodzajów czucia oraz z polami
kojarzeniowymi. Hipokamp ma pośrednie połączenia z korą mózgu
bieguna przedniego płata skroniowego, z korą okołowęchową i z korą
zakrętu parahipokampa. Dzięki tym połączeniom z korą mózgu oraz
jądrami ciał suteczkowatych i przedniego wzgórza układ limbiczny uczestniczy w procesach uczenia się i zapamiętywania.
204
Czucie i ruch
I Uczenie się i zapamiętywanie
Jedną z podstawowych czynności ośrodkowego układu nerwowego
jest przechowywanie śladów po odebranych bodźcach, czyli ich zapamiętywanie. Czynność ta zapewnia coraz lepsze utrzymywanie homeostazy organizmu ponieważ umożliwia zmianę zachowania dzięki zdobytemu doświadczeniu. Jest to zdolność do uczenia się (learning).
Pola kojarzeniowe
kory mózgu
i
Zakręt obręczy
Jądro pr zednie
wzgórza
■
Zakręt hipokampa
i
Jądra ciała
suteczkowatego
podwzgórza
Hipokamp
Ryc. 109. Schemat struktur mózgowia uczestniczących w tworzeniu
się pamięci świeżej.
Bodźce odebrane przez receptory zostają w nich przetworzone na
salwy impulsów nerwowych generowanych zgodnie ze wzorcem zależnym
od bodźca. Salwy impulsów nerwowych poprzez drogi swoiste i nieswoiste przewodzone są do kory mózgu, do układu limbicznego i do
innych struktur przodomózgowia. Pomiędzy neuronami w strukturach
mózgowia zaczynają krążyć impulsy nerwowe, co warunkuje wystąpienie
pamięci natychmiastowej (immediate memory). Przechowanie śladów po
odebranym bodźcu, na krótki czas — kilkudziesięciu sekund do kilkudziesięciu minut, stanowi pamięć świeżą, czyli krótkotrwałą (short term
memory). W tym czasie impulsy nerwowe wywołane odebranym czuciem
bodźca krążą pomiędzy korą mózgu i strukturami układu limbicznego.
Warunkiem trwałego zapamiętywania, czyli pamięci trwałej (lond-term
memory), jest konsolidacja pamięci (consolidation memory). Proces konsolidacji musi trwać dostatecznie długo, od kilku do kilkudziesięciu
minut, tak aby mogły powstać trwałe zmiany w połączeniach pomiędzy
Uczenie się i zapamiętywanie
205
neuronami w postaci nowych połączeń synaptycznych i mogło dojść do
ekspresji określonych genów w jądrach komórkowych (patrz str. 81).
W czasie procesu konsolidacji następuje około 100-krotne zmniejszenie
informacji zapamiętanej w stosunku do informacji w pełni percepowanej.
U człowieka w ciągu doby, w czasie której 16 godzin przypada na czuwanie,
percepowanych jest średnio 20 bitów informacji na sekundę, czyli około
1 x 1016 bitów informacji na dobę. Ta ilość informacji wprowadzona do
pamięci świeżej jest około 100 razy większa od pojemności pamięci trwałej,
która warunkowana jest liczbą komórek nerwowych i połączeń między
nimi.
i Pamięć trwała
Czynności związane z pamięcią trwałą przebiegają w czterech fazach.
Jest to zapamiętywanie bodźców, przechowywanie śladów pamięciowych,
przypominanie śladów pamięciowych i rozpoznawanie nowych bodźców
przez ich porównanie z poprzednio zapamiętanymi bodźcami.
PAMIĘĆ TRWAŁA
PAMIĘĆ NIEOPISOWA
PAMIEC OPISOWA
-proceduralna
- słowne opisywanie
zapamiętanych słów i faktów
WPRAWA
PAMIĘĆ
w wykonywaniu
-staw ne
czynności
odtw arzanie
ruchowych
SEMANTYCZNA
zopamie, tanych
UŁATWIENIE
wykonywania stów, praw }
twierdzeń
czynności
ruchowych
WARUNKOWANIE asocjacja pomiędzy
bodźcami działającymi
w przeszłości
HABITUACJA
UWRAŻLIWIENIE nieasocjacyjne
zmniejszenie lub
zwiększenie reakcji
na powtarzający się
bodziec
Ryc. 110. Rodzaje pamięci trwałej.
PAMIĘĆ
EPIZODYCZNA
-słow ne
opisywanie
zdarzeń
i faktów
z przeszłości
206
Czucie i ruch
Pamięć trwała dzieli się na pamięć proceduralną i opisową. Pamięć
proceduralna (procedural memory) związana jest z uzyskiwaniem wprawy
(skills) w wykonywaniu czynności ruchowych, ułatwianiem w ich wykonywaniu (priming), warunkowaniem (conditioning) i z innymi czynnościami związanymi z zapamiętywaniem.
Odruchy warunkowe
Odruchy warunkowe — CR (conditioned reflexes), czyli odruchy
nabyte, powstają wtedy, kiedy bodziec obojętny wyprzedza działanie
bodźca bezwarunkowego — US (unconditioned stimulus). Po jednym,
kilku lub wielu powtórzeniach bodziec warunkowy zaczyna wyzwalać
taką samą reakcję jaka występuje w czasie działania bodźca bezwarunkowego.
Siła odruchu warunkowego jest proporcjonalna do siły bodźca warunkowego tylko w pewnych granicach. Po przekroczeniu tej granicy silniejszy bodziec warunkowy wyzwala słabszy odruch warunkowy.
W życiu osobniczym stale się tworzą nowe odruchy warunkowe,
a poprzednio utworzone mogą zanikać. Nowy bodziec, działając po raz
pierwszy jednocześnie z bodźcem warunkowym (CS), osłabia odruch
warunkowy. Zjawisko to nosi nazwę hamowania zewnętrznego odruchów
warunkowych (external inhibition).
Hamowanie wewnętrzne odruchów
warunkowych
Hamowanie wewnętrzne odruchów warunkowych (internal inhibition)
wiąże się z:
• wygasaniem odruchów (extinction) na skutek braku wzmocnienia
bodźca warunkowego przez bodziec bezwarunkowy;
• hamowaniem warunkowym, kiedy bodziec warunkowy, występując
jednocześnie z innym bodźcem, nie jest wzmocniony;
• hamowaniem opóźniającym, kiedy działanie bodźca bezwarunkowego
znacznie się opóźnia w stosunku do bodźca warunkowego;
• hamowaniem różnicującym, kiedy dwa bodźce warunkowe różnią się
nieznacznie od siebie. Tylko pierwszy z tych bodźców jest wzmacniany
przez bodziec bezwarunkowy, reakcja na drugi bodziec zaś jest częś
ciowo lub całkowicie zahamowana.
Uczenie się i zapamiętywanie
207
Różna szybkość tworzenia się i wygasania odruchów warunkowych
stanowi według Pawłowa kryterium podziału układów nerwowych na
cztery typy, czyli temperamenty:
•
•
•
•
typ o słabych procesach nerwowych (melancholik);
typ silny, niezrównoważony (choleryk);
typ silny, zrównoważony, ruchliwy (sangwinik);
typ silny, zrównoważony, powolny (flegmatyk).
Wytwarzanie się łuku odruchowego odruchu warunkowego jest uzależnione od odpowiedniej pobudliwości wielu ośrodków. Przede wszystkim zależy od:
• przewodzenia impulsów aferentnych przez drogi nieswoiste, czyli przez
wstępujący układ siatkowaty (ascending reticular system);
• pobudliwości ośrodków motywacyjnych w międzymózgowiu i w ukła
dzie rąbkowym;
• kory mózgu.
Wyższa czynność nerwowa
Wyższa czynność nerwowa człowieka wiąże się z filogenetycznie
najmłodszymi polami kory mózgu. Są to pola kojarzeniowe, czyli asocjacyjne, których wyróżnia się trzy:
• czołowa okolica kojarzeniowa (frontal association area) zajmuje najbar
dziej ku przodowi położoną część płata czołowego i okolicę oczodo
łową kory mózgu;
• skroniowa okolica kojarzeniowa (temporal association area) znajduje się
w części przedniej płata skroniowego;
• potyliczno-skroniowo-ciemieniowa okolica kojarzeniowa (occipito-temporo-parietal association area) obejmuje pogranicze tych trzech pła
tów.
Czołowa okolica kojarzeniowa związana jest z funkcją zapamiętywania, abstrakcyjnym myśleniem, przewidywaniem, planowaniem postępowania oraz hamowaniem popędów i emocji. Skroniowa oko lica kojarzeniowa, dzięki licznym połączeniom z hipokampem leżącym
w głębi płata skroniowego, jest „magazynem" trwale zapamiętywanych
wrażeń zmysłowych. W potyliczno-skroniowo-ciemieniowej okolicy kojarzeniowej znajduje się nadrzędny ośrodek mowy w dominującej półkuli
mózgu.
208
Czucie i ruch
Ośrodki mowy
W korze mózgu człowieka w dominującej półkuli (dominant he misphere), lewej u znacznej większości ludzi, znajdują się:
• ośrodek ruchowy mowy Broca (Broca's speech center) związany z ar
tykulacją dźwięków w postaci słów;
• ośrodek ruchów pisarskich ręki (ability to write area) koordynujący
skurcze mięśni w czasie pisania;
• ośrodek słuchowy mowy (word deafness area) umożliwiający rozróż
nienie słyszanych słów;
• ośrodek wzrokowy mowy (word blindness area) pozwalający rozpo
znawać znaki pisarskie.
Ryc. 111. Umiejscowienie ośrodków mowy w korze mózgu człowieka. 1 — ośrodek ruchowy
mowy, 2 — ośrodek ruchów pisarskich ręki, 3 — ośrodek słuchowy mowy, 4 — ośrodek
wzrokowy mowy.
Ośrodki te są kontrolowane przez nadrzędny ośrodek mowy znajdujący się w dominującej półkuli mózgowej w okolicy kojarzeniowej potyliczno-skroniowo-ciemieniowej. Ośrodek nadrzędny połączony jest z innymi ośrodkami mowy za pośrednictwem poduszki wzgórza (puhinar
thalami).
Uczenie się i zapamiętywanie
209
Tabela 8
Umiejscowienie funkcji w korze mózgu człowieka
Funkcja
Płat
Czucie
Pole wg
Brodmanna
powonienie
skroniowy i
ciało
migdałowate
hak zakrętu
hipokampa
wzrok
potyliczny
kora otaczająca
bruzdę ostrogową
17
słuch
skroniowy
skroniowy górny
41
dotyk
ciemieniowy
zaśrodkowy
1,2, 3
temperatura
ciemieniowy
zaśrodkowy
1, 2,3
ból
ciemieniowy
zaśrodkowy
1, 2,3
proprioceptywne
ciemieniowy
zaśrodkowy
1,2,3
interoceptywne
ciemieniowy
zaśrodkowy
1, 2,3
czołowy
przedśrodkowy
zapamiętywanie
czołowy
(część
przednia)
czołowy górny
czołowy środkowy
czołowy dolny
oczodołowe
trwała pamięć
skroniowy
(część
przednia)
skroniowy górny
skroniowy środkowy skroniowy
dolny
mowa ośrodek
nadrzędny
potyliczno-skroniowociemieniowy
teleceptywne
eksteroceptywne
Ruch: pierwszorzędowa reprezentacja
Kojarzenie
Zakręt
4
9, 10, 11, 12
38
39,40
210
I
Czucie i ruch
Zespoły czynnościowe ośrodkowego
układu nerwowego
Ośrodkowy układ nerwowy charakteryzuje się złożoną budową zarówno makroskopową, jak i mikroskopową. Podstawowe elementy czynnościowe, jakimi są komórki nerwowe, różnią się znacznie pod względem morfologicznym i funkcjonalnym. Ciała neuronów często znajdują się w jednej strukturze, zakończenia tych neuronów występują zaś
w innej strukturze, nawet dość odległej. W obrębie jednej struktury
sąsiadują ze sobą neurony pobudzające i hamujące, a więc uwalniające
na swych synapsach przekaźniki chemiczne depolaryzujące lub hiperpolaryzujące postsynaptyczną błonę komórkową.
Liczne i różnorodne zależności występujące pomiędzy neuronami
jednej struktury i neuronami pozostałych struktur ośrodkowego układu
nerwowego znacznie utrudniają, a nawet uniemożliwiają, wybiórcze
poznanie prawidłowej funkcji jednej struktury. Szczególne trudności
występują w czasie prób umiejscowienia pamięci w określonych strukturach mózgowia.
Odstępując od zasady przypisywania każdej strukturze mózgowia
określonej funkcji, można w oderwaniu od struktury przedstawić czynność ośrodkowego układu nerwowego w postaci zespołów czynnościowych. Takie postępowanie przyjęte jest powszechnie w technice. Schemat
blokowy, np. komputera, pozwala znacznie szybciej zgłębić zasadę jego
działania niż szczegółowy schemat połączeń pomiędzy wszystkimi jego
elementami.
Wszystkie receptory w organizmie spełniają funkcję przetworników.
W ich obrębie zachodzi zmiana bodźców ze środowiska zewnętrznego
i wewnętrznego organizmu na salwy impulsów nerwowych. Ten sam
bodziec działający w tych samych warunkach zostaje zamieniony w przetwornikach na taką samą salwę impulsów. Salwy impulsów różnią się
między sobą nie tylko czasem trwania, lecz również przebiegiem narastania i zmniejszania się częstotliwości impulsów. Salwy impulsów nerwowych tworzą określone wzorce w każdym pojedynczym włóknie
aferentnym, a więc w obrębie jednej jednostki czuciowej. Bodziec działający na liczne receptory zostaje jednocześnie przetworzony na określone
wzorce impulsów w odpowiednio licznych jednostkach czuciowych.
Salwy impulsów nerwowych stanowiące odwzorowanie bodźca są
przewodzone równolegle przez liczne jednostki czuciowe do ośrodkowego
układu nerwowego i zostają tam poddane wstępnej analizie w pierwszym
zespole czynnościowym. Analiza odwzorowanego bodźca przeprowadzo-
Zespoły czynnościowe ośrodkowego układu nerwowego
211
na jest w sprzężeniu z drugim zespołem zawierającym zmagazynowane
lub zapamiętane wzorce bodźców, które uprzednio — tj. w ciągu całego
życia osobniczego — działały na receptory.
Z pamięci zostaje wywołany jeden wzorzec bodźca. W zespole komparatora zapamiętany wzorzec bodźca jest przyrównany do bodźca
aktualnie działającego na receptory. Stąd impulsacja zostaje przekazana
do zespołu sprawującego kontrolę reakcji orientacyjnej. Charakter tej
reakcji, której pierwszym zadaniem jest przestrojenie przetworników
w układzie ruchowym, zależy od znaczenia odbieranego bodźca.
Ośrodkowy układ nerwowy
Układ ruchowy
Ryc. 112. Schemat blokowy zespołów czynnościowych ośrodkowego układu nerwowego
kontrolujących układ ruchowy.
Zespół rozrusznika aktywności somatycznej obejmuje szereg sprzężo-
nych ze sobą ośrodków motywacyjnych, stanowiących mechanizm unikania i mechanizm konsumowania. Ten sam bodziec działający na
receptory wyzwala określoną reakcję ruchową w zależności od wypadkowej pobudliwości ośrodków motywacyjnych. Przewaga jednego z mechanizmów motywacyjnych, tj. unikania lub konsumowania, determinuje
określoną reakcję somatyczną. W czasie kiedy równowaga obu mechanizmów motywacyjnych jest zachowana, wówczas — mimo działającego
bodźca — nie dochodzi do reakcji somatycznej. Ruch jest kontrolowany
przez zespół koordynacji ruchów i wyzwalany bezpośrednio przez zespół
kontroli ruchów ciała.
Wszystkie zespoły czynnościowe ośrodkowego układu nerwowego są
utrzymywane w stanie gotowości dzięki układowi siatkowatemu wstępu-
212
Czucie i ruch
jącemu pobudzającemu — RAS. Cały ośrodkowy układ nerwowy jest
również stale zasilany za pośrednictwem krwi w tlen i w produkty
energetyczne. Brak impulsacji z układu siatkowatego wstępującego pobudzającego, co występuje w czasie snu, blokuje odwracalnie czynność
zespołów. Zatrzymanie krążenia krwi na pewien czas lub inne zaburzenia
wewnątrzkomórkowej przemiany materii prowadzą do nieodwracalnego
zaniku czynności niektórych lub wszystkich zespołów.
4
Kontrola środowiska
wewnętrznego
Zachowanie równowagi homeostatycznej
Zmiany w środowisku zewnętrznym zakłócają równowagę pomiędzy
środowiskiem zewnętrznym i wewnętrznym organizmu, czyli równowagę
homeostatyczną (homeostatic eąuilibrium). Ponowne osiągnięcie równowagi homeostatycznej wymaga zazwyczaj współdziałania wielu układów
w organizmie. To najszerzej pojęte dążenie do przywrócenia równowagi
homeostatycznej jest kontrolowane i regulowane zarówno przez ośrodki
nerwowe i podporządkowany im układ nerwowy autonomiczny, jak
również na drodze humoralnej przez gruczoły dokrewne oraz liczne
komórki wydzielające przekaźniki chemiczne.
Ośrodki nerwowe znajdujące się na różnych piętrach ośrodkowego
układu nerwowego odbierają informację od receptorów całego ciała,
przetwarzają je i programują odpowiedź adekwatną do zapotrzebowania.
Czynności takie wykonywane są przede wszystkim przez ośrodki znajdujące się w podwzgórzu. Zaprogramowana w ośrodkach odpowiedź
przekazywana jest do tkanek za pośrednictwem:
• układu nerwowego autonomicznego;
• gruczołów dokrewnych.
Równowaga homeostatyczną przywracana jest również dzięki informacjom, w postaci przekaźników chemicznych, wydzielanych i przesyłanych na drodze humoralnej przez komórki wielu tkanek w organizmie.
Przekaźniki chemiczne wytwarzane przez zaktywowane komórki krążą
w płynach ustrojowych i — wiążąc się ze specyficznymi receptorami
w innych, często odległych komórkach — przyspieszają lub zwalniają ich
metabolizm. Biorą w tym udział często te same przekaźniki chemiczne,
21 4
Kontrola środowiska wewnętrznego
które uwalniane są z zakończeń synaptycznych neuronów, jako transmittery synaptyczne. Są one wydzielane również przez inne komórki,
stanowiąc komunikację autokrynną, parakrynną i endokrynną (patrz
str. 54).
W komunikacji pomiędzy komórkami przywracającej homeostazę
uczestniczą nawet tak wyspecjalizowane komórki, jak kardiocyty mięśnia
przedsionków serca. Zwiększenie objętości płynu zewnątrzkomórkowego
(ECF), szczególnie tej jego części, którą stanowi osocze krwi, powoduje
wydzielanie przedsionkowego peptydu natriuretycznego (ANP) przez
kardiocyty mięśnia przedsionków. Peptyd ten — krążąc we krwi —
działa na nerki, zwiększając filtrację kłębuszkową i wydalanie jonów
sodowych z moczem. Prowadzi to do zmniejszenia objętości płynu
zewnątrzkomórkowego (ECF) i przywraca równowagę homeostatyczną
w zakresie przemiany wodno-elektrolitowej w organizmie.
I Układ nerwowy autonomiczny
Układ nerwowy autonomiczny tworzą komórki nerwowe, które za
pośrednictwem swoich wypustek przewodzą impulsy nerwowe do narządów wewnętrznych. Układ ten stanowi część eferentną trzewnych łuków
odruchowych. Efektorem w łukach odruchowych autonomicznych są
wszystkie inne komórki i tkanki organizmu poza komórkami mięśniowymi mięśni poprzecznie prążkowanych, szkieletowych, unerwionych
przez układ nerwowy somatyczny.
Najprostszy łuk odruchowy autonomiczny ma co najmniej dwa neurony przewodzące eferentną impulsację. Ciała pierwszych neuronów —
przedzwojowych (preganglionic neurons), wysyłających impulsy do narządów wewnętrznych, znajdują się w obrębie ośrodkowego układu nerwowego, a ich wypustki tworzą włókna nerwowe przedzwojowe (preganglionic axons), kończące się synapsami na drugich neuronach zwojowych
(ganglionic neurons) w zwojach autonomicznych. Wypustki tych drugich
neuronów, czyli zazwojowe aksony (postganglionic axons), tworzą włókna
zazwojowe kończące się synapsami na komórkach narządów wewnętrznych.
Układ autonomiczny (systema nervosum autonomiami) dzieli się zarówno pod względem anatomicznym, jak i czynnościowym na dwie części:
• część współczulną (pars sympathica);
• część przywspółczulną (pars parasympathica).
Układ nerwowy autonomiczny
21 5
I Część współczulna układu autonomicznego
Pierwsze neurony współczulne znajdują się w istocie szarej rdzenia
kręgowego w rogach bocznych (cornu laterale) w części piersiowej i górnej lędźwiowej w segmentach od Thl do L3. Ich wypustki, czyli włókna
przedzwojowe, opuszczają rdzeń kręgowy w korzeniach brzusznych (radices ventrales) i za pośrednictwem gałęzi łączących białych (rami communicantes albi) wchodzą do zwojów pnia współczulnego (ganglia trunci
sympathici), gdzie kończą się synapsami na drugich neuronach współczulnych. Wypustki tych neuronów, czyli włókna zazwojowe, przez gałęzie
łączące szare (rami communicantes grisei) powracają do nerwów rdzeniowych, wraz z którymi biegną do mięśni i skóry. Włókna te unerwiają
mięśnie gładkie naczyń krwionośnych oraz gruczoły i komórki mięśni
gładkich skóry.
WłóknC
eferentne
do trzewi
"
W łókna
aferenine
od trzewi
Ryc. 113. Część ośrodkowa autonomicznego łuku odruchowego: linia czarna ciągła —
aferentne włókno nerwowe, linia czerwona ciągła — eferentne włókno nerwowe współczulne
przedzwojowe, linia czerwona przerywana — eferentne włókno nerwowe współczulne
zazwojowe.
216
Kontrola środowiska wewnętrznego
Część wypustek pierwszych neuronów wspólczulnych nie kończy się
synapsami w zwojach pnia współczulnego, lecz biegnie dalej:
• w części szyjnej do zwoju szyjno-piersiowego, czyli gwiaździstego
(ganglion cervicothoracicum sive stellatum), do zwoju szyjnego środ
kowego (ganglion cewicale medium) i do zwoju szyjnego górnego
{ganglion cervicale superius);
• w części piersiowej i brzusznej do zwoju trzewnego {ganglion splanchnicum), do zwoju krezkowego górnego {ganglion mesentericum superius)
i do zwoju krezkowego dolnego {ganglion mesentericum inferius).
Neurony w zwojach współczulnych w części szyjnej, piersiowej i brzusznej wysyłają wypustki w postaci włókien zazwojowych do wszystkich
tkanek i narządów głowy, szyi, klatki piersiowej, jamy brzusznej, włącznie z narządami miednicy mniejszej i narządami płciowymi.
Niewielka liczba wypustek pierwszych neuronów współczulnych (włókna przedzwojowe) nie kończy się synapsami w zwojach współczulnych,
lecz bezpośrednio unerwia naczynia krwionośne w mięśniach szkieletowych oraz rdzeń nadnerczy {medulla glandulae suprarenalis).
Na zakończeniach włókien przedzwojowych, czyli włókien nerwowych
typu B, wydziela się acetylocholina, a na zakończeniach włókien zazwojowych, czyli włókien typu Cs, uwalnia się inny transmitter — noradrenalina.
Transmitter noradrenergiczny
Noradrenalina (NA) uwalniająca się na zakończeniach włókien współczulnych zazwojowych częściowo jest syntetyzowana w zakończeniach
tych włókien i częściowo wychwytywana z krwi krążącej. Na zakończeniach włókien typu Cs jest magazynowana w pęcherzykach synaptycznych noradrenergicznych związana z adenozynotrifosforanami (ATP)
i z białkami — chromograninami.
Noradrenalina w obrębie synaps i poza synapsami jest syntetyzowana
z aminokwasu L-tyrozyny przekształconego w wątrobie z innego aminokwasu — fenyloalaniny. Z tyrozyny powstaje DOPA (L-3,4-dihydroksyfenyloalanina), która przekształca się w dopaminę, a ta zamienia się
w L-noradrenalinę.
Noradreanalina uwolniona z zakończeń do płynu międzykomórkowego dyfunduje i dostaje się do krwi. W czasie dyfuzji i przechodzenia do naczyń włosowatych NA jest częściowo rozkładana przez tlenową
metylotransferazę katecholową — COMT (catechol-O-methyltran-
Układ nerwowy autonomiczny
21 7
I -" ---------------- Oksydaza monoaminow alMAO}
Kw as di hydr oksymigdolow y
I — ---------------- Tlenowa metylotransferaza katecholowa IC0MT1
Kw as 3- metoks y-i - hy dr ok sy mi gda łow y
(kw as w anilinooksymigdałowy— VMA)
Ryc. 114. Przemiana aminokwasu fenyloalaniny w organizmie. Metabolity fenyloalaniny —
dopamina i noradrenalina — są zmagazynowane w pęcherzykach synaptycznych w zakończeniach adrenergicznych.
sferase) — na nieaktywną 3-metoksynoradrenalinę. Natomiast noradrenalina powracająca do zakończeń nerwowych podlega tam działaniu
oksydazy monoaminowej — MAO (monoamine oxidase), znajdującej
się w mitochondriach, i jest zamieniana na kwas 3-metoksy-4-hydroksymigdałowy — VMA (vanilmandelic acid), przechodzący następnie do
krwi.
Ostateczny wynik działania noradrenaliny na komórki narządów
wewnętrznych, a szczególnie na komórki mięśniowe gładkie, zależy od
stosunku ilościowego w błonie komórkowej receptorów wrażliwych na
NA.
Receptory adrenergiczne
W błonie komórkowej komórek narządów wewnętrznych występują
receptory adrenergiczne: alpha (ax, a2) i beta (P15 P2). Receptory alpha są
najbardziej wrażliwe na cząsteczki noradrenaliny, receptory beta zaś
najsilniej reagują pod wpływem izopropylonoradrenaliny (isoproterenol).
21 8
Kontrola środowiska wewnętrznego
Receptory adrenergiczne są to białka o długich łańcuchach polipeptydowych przeplatających się siedem razy przez błonę komórkową. Pętle
cytopłazmatyczne tych białek receptorowych łączą się z białkami G (patrz
str. 32). Związanie się noradrenaliny z receptorem alphaj adrenergicznym
wywołuje w komórce zwiększenie stężenia inozytolotrifosforanu (IP3 )
i diacy loglicerolu (DAG) oraz hamowanie odkomórkowego prądu jonów
potasowych. Działanie noradrenaliny poprzez receptor alpha 2 adrenergiczny wiąże się z hamowaniem tworzenia się w komórce cyklicznego
adenozynomonofosforanu (cAMP) i jednoczesnym otwieraniem kanałów dla odkomórkowego prądu jonów potasowych i zamykaniem kanałów dla dokomórkowego prądu jonów wapniowych.
Działanie noradrenaliny przez receptory adrenergiczne beta x i beta2
wiąże się z aktywacją adenylocyklazy w błonie komórkowej i zwiększeniem w komórkach stężenia cAMP, czyli ma działanie metabolitotropowe.
Istnieje liczna grupa preparatów farmakologicznych wybiórczo blokujących alpha- lub beta-receptory adrenergiczne. Dzięki temu można na
drodze farmakologicznej uzyskiwać przewagę procesów związanych z pobudzeniem jednego rodzaju receptorów adrenergicznych. Blokerem, receptorów alpha-adrenergicznych jest fenoksybenzamina, blokerem receptorów beta-adrenegicznych zaś propranolol.
Większość neuronów noradrenergicznych, zwojowych, zawiera neuropeptyd Y (NPY). Mniejszość neuronów adrenergicznych zawiera somatostatynę (SRIF) i enkefaliny (Enk-Leu i Enk-Met). Neuropeptydy te
uwolnione wraz z noradrenałiną działają modułująco na przewodnictwo
noradrenergiczne.
Przewodzenie w zwojach współczulnych
Acetylocholina uwolniona na zakończeniach neuronów przedzwojowych działa jednocześnie na receptory nikotynowe i receptory muskarynowe w błonie komórkowej neuronów zwojowych. Wiązanie się acetylocholiny z receptorem nikotynowym wywołuje skutek podobny do działania
nikotyny. Neuron zwojowy szybko się pobudza i równie szybko pobudzenie to ustępuje. Związki antagonistyczne w stosunku do nikotyny, np.
heksametonium, blokują receptory nikotynowe.
Pobudzenie neuronu zwojowego za pośrednictwem receptora muskarynowego jest podobne do działania muskaryny i może zostać zablokowane przez atropinę. Impulsy nerwowe przekazywane przy udziale receptora muskarynowego wolno pobudzają neuron zwojowy, ale jego pobu-
Układ nerwowy autonomiczny
21 9
dzenie trwa dłużej w porównaniu z pobudzeniem wywołanym za pośrednictwem receptora nikotynowego.
W zwojach współczulnych występują również neurony pośredniczące.
Cząsteczki acetylocholiny uwalniane z zakończeń neuronów przedzwojowych pobudzają neurony pośredniczące, które z kolei oddziałują hamująco
na neurony zwojowe. Na synapsach neuronów pośredniczących wydzielana jest dopamina, która hiperpolaryzuje błonę komórkową neuronów zwojowych i zmniejsza ich pobudliwość, działając poprzez receptor
dopaminergiczny D2. Haloperidol jest preparatem blokującym receptory
dopaminergiczne i przewodzenie przez neurony pośredniczące w zwojach
współczulnych.
Przewodzenie impulsów nerwowych przez zwoje współczulne jest procesem złożonym. Acetylocholina działa bezpośrednio na neurony zwojowe — pobudzająco, i przy udziale neuronów pośredniczących — hamująco. Ostateczny wynik zależy od wzajemnych stosunków pomiędzy receptorami nikotynowymi i muskarynowymi, za których pośrednictwem neuron zwojowy jest pobudzany, a receptorem dopaminergicznym — hamującym, na który działa dopamina uwalniana przez neuron pośredniczący.
I
Część przywspółczulna układu
autonomicznego
Pierwsze neurony przywspółczulne ośrodkowe skupione są w:
• części przywspółczulnej jąder nerwów czaszkowych: okoruchowego —
n. III, twarzowego — n. VII, językowo-gardłowego — n. IX i błędne
go — n. X;
• części krzyżowej rogów bocznych rdzenia kręgowego w segmentach
S2—S4.
Wypustki tych neuronów jako włókna przedzwojowe typu B biegną
w nerwach czaszkowych: okoruchowym — n. III, twarzowym — n. VII,
językowo-gardłowym — n. IX i błędnym — n. X oraz tworzą nerwy
trzewne miedniczne (nervi splanchnici pefoini) po wyjściu z części krzyżowej rdzenia kręgowego S2—S4.
Około 90% włókien nerwowych przywspółczulnych biegnie w nerwie
błędnym. Nerw ten przewodzi do większości narządów wewnętrznych
impulsy odgrywające zasadniczą rolę w układzie przywspółczulnym.
Przywspółczulne włókna przedzwojowe biegną do narządów wewnętrznych i w ich obrębie kończą się synapsami na drugich neuronach
220
Kontrola środowiska wewnętrznego
przywspółczulnych zwojowych. Neurony te rozsiane są w ścianach narządów wewnętrznych, a ich aksony rozgałęziają się w niewielkiej odległości
od ciała neuronu, kończąc się na komórkach: gruczołowych, mięśni
gładkich, mięśnia sercowego oraz na innych. Komórki zwojowe przywspółczulne unerwiają tkanki, w których same się znajdują, mają krótkie
aksony i nie tworzą dłuższych włókien zazwojowych, tak jak to występuje
w zwojach współczulnych.
~
j
r
~\/-
M. zwieracz i m.
rozwieracz źrenicy
Gruczoły łzowe i ślinowe
Tarczyca
Oskrzela i płuca
Ryc. 115. Schemat autonomicznego układu nerwowego. W części przywspółczulnej aksony
neuronów przedzwojowych (linie ciągłe czarne) biegną w nerwie okołoruchowym (n. III),
twarzowym (n. VII), językowo-gardłowym (n. IX), błędnym (n. X) i nerwie trzewnym
miednicznym. Przywspółczulne neurony zwojowe wysyłają swoje wypustki do narządów
(linie przerywane czarne). W części współczulnej aksony neuronów przedzwojowych (linie
ciągłe czerwone) biegną do zwojów współczulnych: zwoju szyjnego górnego, zwojów pnia
współczulnego, zwoju trzewnego, zwoju krezkowego górnego i do zwoju krezkowego
dolnego. Współczulne neurony zwojowe wysyłają swoje aksony (linie przerywane czerwone)
biegnące w nerwach współczulnych do narządów.
Układ nerwowy autonomiczny
221
Na zakończeniach zarówno neuronów przywspółczulnych przedzwojowych, jak i zazwojowych wydziela się acetylocholina.
Transmitter cholinergiczny
Acetylocholina (Ach) syntetyzowana jest w zakończeniach nerwowych
przy udziale enzymu — acetylotransferazy cholinowej, która przenosi
grupę acetylową z acetylo-koenzymu A (Acetyl-CoA) na cholinę.
Magazynowana jest w pęcherzykach synaptycznych w obrębie zakończeń synaptycznych. Impuls nerwowy uwalnia z zakończeń pewną liczbę
cząsteczek acetylocholiny, która działa na receptory cholinergiczne w błonie komórkowej narządów wewnętrznych. Acetylocholina wiąże się z tzw.
receptorem muskarynowym i nikotynowym.
Receptory cholinergiczne
Receptor nikotynowy w błonie komórkowej narządów wewnętrznych
jest podobny do receptora w błonie postsynaptycznej zakończenia synaptycznego nerwowo-mięśniowego. Białko stanowiące receptor nikotynowy
zbudowane jest z pięciu podjednostek, z których każda przechodzi przez
całą grubość błony komórkowej. Pięć podjednostek tworzy w błonie
komórkowej kanał o średnicy około 0,5 nm w stanie spoczynku. Związanie
się acetylocholiny z dwiema takimi samymi podjednostkami powoduje
zmianę konformacji białka i średnica kanału poszerza się. Przez ten kanał
napływają do komórki jony sodowe i powodują jej aktywację.
Wyróżnia się pięć typów receptorów muskarynowych: M,, M2, M3, M4 i
M5. Związanie się acetylocholiny z receptorem M,, M4 lub M5 powoduje
uwalnianie się z wewnętrznej powierzchni błony komórkowej do cytoplazmy inozytolotrifosforanu (IP 3) i diacyloglicerolu (DAG). Natomiast
po związaniu się acetylocholiny z receptorami M 2 lub M3 następuje
inaktywacja cyklazy adenylanowej w błonie komórkowej i zmniejsza się
stężenie cyklicznego adenozynomonofosforanu (cAMP) w komórkach.
Dodatkowo następuje zamykanie kanałów dla dokomórkowego prądu
jonów wapniowych.
Różne typy receptorów cholinergicznych występują w błonie komórkowej różnych tkanek i narządów. Duża gęstość receptorów cholinergicznych typu M 2 występuje w błonie komórkowej komórek mięśnia sercowego, typu M4 zaś w komórkach zewnątrz- i wewnątrzwydzielniczych
trzustki. Preparaty farmakologiczne wybiórczo blokują jeden z recep-
222
Kontrola środowiska wewnętrznego
torów, np. atropina blokuje receptor muskarynowy, heksametonium zaś
receptor nikotynowy.
Acetylocholina uwolniona na synapsach jest rozkładana przez swoistą
esterazę cholinową (specific cholinesterase), cząsteczki ACh dyfundujące
poza synapsy i dostające się do krwi są zaś rozkładane przez nieswoistą
esterazę cholinową (pseudocholinesterase, nonspecific cholinesterase).
W neuronach cholinergicznych zwojowych występuje wazoaktywny peptyd jelitowy (VIP), działając jako modulator. Neurony cholinergiczne
zwojowe zawierają po kilka neuropeptydów jednocześnie. W neuronach
splotu warstwy mięśniowej jelita (plexus myentericus) koegzystują neuropeptydy: wazoaktywny peptyd jelitowy (VIP), cholecystokinina (CCK-8),
dynorfiny (Dyn 1—8 i Dyn 1—17) i enkefaliny (Enk-Met i Enk-Leu).
Poza tymi modulatorami w innych cholinergicznych neuronach zwojowych stwierdzono występowanie: tachikinin (Substancji P i K), peptydu
uwalniającego gastrynę (GRP), somatostatyny (SRIF), neuropeptydu
Y (NPY), galaniny, peptydu pochodnego od genu kalcytoninowego
(CGRP) i neurotensyny.
Działanie na efektory impulsacji
współczulnej i przywspółczulnej
I
W obrębie komórek efektorów występuje stałe współzawodnictwo
pomiędzy impulsacją przywspółczulną, czyli cholinergiczną, a współczulną, czyli adrenergiczną. Impulsacją współczulna pobudza lub hamuje
aktywność komórek efektora w zależności od przewagi receptorów alpha
lub beta w ich błonie komórkowej. Impulsacją przywspółczulną ma
zawsze działanie przeciwne.
Na procesy przebiegające w komórkach efektorów układy cholinergiczny i noradrenergiczny wywierają swój wpływ za pośrednictwem wewnątrzkomórkowych przenośników informacji, którymi są cykliczny guanozynomonofosforan (cGMP) i cykliczny adenozynomonofosforan (cAMP).
Zwiększenie zawartości w komórce pierwszego związku — cGMP, hamuje tworzenie się drugiego związku — cAMP, i odwrotnie. Współzawodnictwo w komórkach efektorów pomiędzy cGMP a cAMP warunkuje ostateczny wynik współzawodnictwa pomiędzy układem cholinergicznym i adrenergicznym. Działanie układu współczulnego i przywspólczulnego na poszczególne narządy przedstawiono w tabeli 9.
Transmittery układu autonomicznego wpływają na metabolizm wewnątrzkomórkowy. Pod wpływem noradrenaliny w komórkach watro-
Układ nerwowy autonomiczny
223
Tabela 9
Działanie impulsacji współczulnej i przywspółczulnej
Układ autonomiczny
Część współczulna
Część
przywspołczulna
Transmitter uwalniany z zakończeń neuronów przedzwojowych:
Transmitter uwalniany z zakończeń neuronów zwojowych:
Transmitter wiąże się
z receptorem:
Oko:
mięśnie tęczówki
— szerokość źrenicy
— mięsień rzęskowy
Gruczoł łzowy:
— wydzielanie
Serce:
układ przewodzący
— częstość skurczów
— przewodnictwo
mięsień sercowy
— siła skurczów
naczynia wieńcowe
— przepływ krwi
Naczynia krwionośne:
— skóry i błon ślizowych
— mięśni szkieletowych
— mózgu
— płuc
— narządów jamy brzusznej
Oskrzela:
— mięsień oskrzelowy
Przewód pokarmowy:
ślinianki
— wydzielanie śliny
żołądek
— motoryka i napięcie
błony mięśniowej
— wydzielanie soku
acetylocholina (ACh)
noradrenalina
alpha-adrenergicznym
(NA)
beta-adrenergicznym
acetylocholina
(ACh)
acetylocholina
(ACh)
muskarynowym
i nikotynowym
rozszerza
—
—
rozkurcza
zwęża
kurczy
—
—
pobudza
—
—
przyspiesza
przyspiesza
zwalnia
zwalnia
—
zwiększa
zmniejsza
zmniejsza
zwiększa
zwiększa
zwęża
zwęża
nieznacznie
zwęża
zwęża
zwęża
—
rozszerza
rozszerza
rozszerza
—
rozszerza
rozszerza
rozszerza
rozszerza
—
—
rozkurcza
kurczy
pobudza wydzielanie gęstej
śliny
—
pobudza wydzielanie wodnistej śliny
zmniejsza
hamuje (?)
zmniejsza
hamuje (?)
wzmaga
pobudza
224
Kontrola środowiska wewnętrznego
cd. tab. 9
Układ autonomiczny
Część współczulna
Część
przywspółczulna
jelita — motoryka i napięcie
błony mięśniowej — wydzielanie
soku
trzustka — wydzielanie soku
pęcherzyk żółciowy — błona
mięśniowa
bańka wątrobowo-trzustkowa —
mięsień zwieracz bańki
zmniejsza
hamuje (?)
zmniejsza
hamuje (?)
wzmaga
pobudza
rozkurcza
pobudza
rozkurcza
kurczy
kurczy
Pęcherz moczowy: — mięsień
wypierający — mięsień zwieracz
wewnętrzny cewki moczowej
kurczy
rozkurcza
kurczy
rozkurcza
bowych zachodzi glikoliza i uwalnianie cząsteczek glukozy do krwi.
Natomiast w komórkach tkanki tłuszczowej następuje hydroliza triacylogliceroli i przechodzenie do krwi wolnych kwasów tłuszczowych
(FFA).
Z zakończeń nerwowych adrenergicznych i cholinergicznych wraz
z transmitterami i modulatorami uwalniany jest adenozynotrifosforan
(ATP). Działa on na błonę presynaptyczną neuronów noradrenergicznych poprzez receptor purynergiczny P, i P 2 i wraz z jonami Ca2+ nasila
uwalnianie transmittera — noradrenaliny. Na zakończenia presynaptyczne neuronów cholinergicznych również działa ATP za pośrednictwem
receptorów purynergicznych Pj i P 2, potęgując działanie acetylocholiny.
Prostaglandyny uwalniane z błon komórkowych modulują transmisję
adrenergiczną i cholinergiczną w neuronach zwojowych. Prostaglandyny
z grupy PGF hamują uwalniane noradrenaliny, nasilają zaś przewodnictwo cholinergiczne. Przeciwne działanie mają prostaglandyny z grupy
PGE nasilające przewodnictwo noradrenergiczne i hamujące przewodnictwo
cholinergiczne.
Czynność gruczołów dokrewnych
225
I Czynność gruczołów
I dokrewnych
Kontrola humoralna w organizmie wiąże się z przenoszeniem informacji
zakodowanych w postaci przekaźników chemicznych, do których należą
hormony.
Wspólną cechą wszystkich hormonów jest to, że wytworzone w jednym
narządzie, tkance lub komórce są przenoszone przez płyny
ustrojowe i oddziałują na czynność innych narządów, tkanek i
komórek.
Przekaźniki chemiczne wydzielane do płynów ustrojowych powstają w:
• wyspecjalizowanych narządach — gruczołach dokrewnych, są to hor
mony gruczołów dokrewnych;
• komórkach rozsianego układu wydzielania wewnętrznego, nazywane są
hormonami tkankowymi;
• komórkach o innej wyspecjalizowanej funkcji jako hormony o działaniu
ogólnym lub miejscowym.
Hormony o działaniu miejscowym oddziałują na sąsiednie komórki
(paracrine communication) poprzez płyn zewnątrzkomórkowy, ale niektóre z tych hormonów mogą być również przenoszone przez krew,
wywierając wpływ na inne odległe narządy i tkanki.
Hormony wytwarzane przez gruczoły dokrewne dzieli się w zależności
od miejsca powstawania oraz budowy chemicznej ich cząsteczki. Hormony wytwarzane w organizmie człowieka podzielone zostały na dwie
grupy:
(1) Zbudowane z aminokwasów lub związków pochodnych.
(2) Pochodne cholesterolu.
Pierwsza grupa — związki pochodne aminokwasów — powstaje w komórkach:
podwzgórza (patrz tab. 10, str. 228);
części gruczołowej przysadki (patrz tab. 11, str. 236);
części pośredniej przysadki (patrz tab. 12, str. 241);
szyszynki (melatonina);
gruczołu tarczowego (trijodotyronina, tyroksyna i kalcytonina);
gruczołów przytarczycznych (parathormon — PTH);
wysp trzustkowych (Langerhansa) (insulina, glukagon, somatosta tyna — SRIF, gastryna i trzustkowy polipeptyd);
• rdzenia nadnerczy (adrenalina, noradrenalina i enkefaliny).
8 — Fizjologia człowieka
226
Kontrola środowiska wewnętrznego
Druga grupa — hormony steroidowe pochodne cholesterolu — powstaje
w:
• korze nadnerczy (glikokortykoidy), mineralokortykoidy i androgeny — tab. 13, str. 244);
• jądrach w komórkach śródmiąższowych (Leydiga) (testosteron);
• jajnikach w komórkach pęcherzyków jajnikowych (Graafa) (estrogeny);
• jajnikach w komórkach ciałka żółtego (progesteron).
W czasie ciąży łożysko wytwarza również hormony należące do
pierwszej i do drugiej grupy.
Hormony zbudowane z aminokwasów i związków pochodnych, przede
wszystkim hormony peptydowe i białkowe, mają inny mechanizm działania na komórki narządów docelowych w porównaniu do mechanizmu
działania hormonów steroidowych. Hormony peptydowe i białkowe
wiążą się z receptorem w błonie komórkowej komórek docelowych,
zmieniają metabolizm wewnątrzkomórkowy, w tym również aktywność
enzymów. W wyniku tego dochodzi do zmian w procesie syntezy związków wytwarzanych przez komórki docelowe.
Czynność gruczołów dokrewnych
Aminokwasy
w cytoplazmie
227
i
Synteza
------- " P^ Pept ydow
w rybosomach
Ryc. 117. Mechanizm molekularny działania hormonów steroidowych na syntezę poli petydów w komórkach narządów docelowych.
Hormony steroidowe natomiast wnikają przez błonę komórkową do
wnętrza komórek docelowych i w połączeniu z receptorem cytoplazmatycznym wpływają na transkrypcję cząsteczek mRNA w jądrze komórkowym. Wywołuje to zmianę w syntezie białek strukturalnych, enzymatycznych i wydzielanych przez komórki na zewnątrz.
I Hormony podwzgórzowe
W podwzgórzu w jądrze przykomorowym (nucleus paraventricularis)
i w jądrze nadwzrokowym (nucleus supraopticus) są u człowieka syntetyzowane dwa pre-pro-hormony: pre-pro-wazopresyno-neurofizyna
druga (pre-pro-AVP-NP II) i pre-pro-oksytocyno-neurofizyna pierwsza
(pre-pro-OXY-NP I). Cząsteczki pre-pro-hormonów upakowane w perykarionie do pęcherzyków neurosekrecyjnych (neurosecretory vesicle),
zwanych również ziarnistościami neurosekrecyjnymi, przesuwane są wewnątrz aksonów do tylnego płata przysadki. W czasie transportu aksonalnego zachodzi w pęcherzykach potranslacyjna modyfikacja pre-prohormonów, w wyniku czego w zakończeniach aksonów w pę-
228
Kontrola środowiska wewnętrznego
cherzykach neurosekrecyjnych znajdują się oddzielne cząsteczki neurohormonów wazopresyny argininowej i neurofizyny II w neuronach
wazopresynoergicznych oraz oksytocyny i neurofizyny I w neuronach
oksy tocynoergicznych.
Uwalnianie neurohormonów — wazopresyny i oksytocyny — z części
nerwowej przysadki do krwi polega na egzocytozie pęcherzyków neurosekrecyjnych z zakończeń aksonów w czasie depolaryzacji ich błony
komórkowej. W procesie tym współdziałają jony wapniowe.
Tabela 10
Hormony podwzgórzowe
Skrót lub
Nazwa hormonu
symbol
Budowa chemiczna
AVP
wazopresyna argininowa
peptyd o 9 aminokwasach
OXY
oksytocyna
hormon uwalniający hormon
kortykotropowy =
kortykoliberyna
hormon uwalniający hormon
tyreotropowy = tyreoliberyna
hormon uwalniający hormony
gonadotropowe = gonadoliberyna
peptyd o 9 aminokwasach
hormon uwalniający hormon
wzrostu = somatokrynina
hormon hamujący uwalnianie
hormonu wzrostu = somatostatyna
peptyd o 44 aminokwasach
czynnik hamujący uwalnianie
prolaktyny = prolaktostatyna
dopamina
CRH
TRH
GnRH
GRH
SRIF
PIF
peptyd o 41 aminokwasach
peptyd o 3 aminokwasach
peptyd o 10 aminokwasach
peptyd o 14 aminokwasach
Wazopresyna
Wazopresyna argininowa — AVP (arginine vasopressin) — występująca u człowieka jest cyklicznym dziewięciopeptydem o masie cząsteczkowej
1084.
Rozerwanie pierścienia peptydowego w wazopresynie, które najłatwiej
zachodzi między 1 i 6 aminokwasem dzięki przerwaniu wiązania dwusiarczkowego, powoduje utratę właściwości fizjologicznych. Wydzielona
do krwi wazopresyna jest dość szybko inaktywowana, czas jej połowicznego rozpadu (T 1^) wynosi 18 minut.
Czynność gruczołów dokrewnych
229
Ryc. 118. Neuron neurosekrecyjny
podwzgórza (a) i jego zakończenia
w części nerwowej przysadki (b)
z uwzględnieniem struktur cytoplazmatycznych. 1 — jądro, 2 — mitochondrium, 3 — ziarnista siateczka śródplazmatyczna, 4 — aparat Golgiego,
5 — dendryt, 6 — akson, 7 — pęcherzyki neurosekrecyjne, 8 — pęcherzyki
synaptyczne. (Reprodukowano za zezwoleniem Michała Karaska.)
Wazopresyna kurczy mięśnie naczyń krwionośnych i zwiększa resorpcję
zwrotną wody w nerkach. W związku z tym jest również nazywana
hormonem antydiuretycznym — ADH (antidiuretic hormone). Za pośrednictwem receptora V2 i dzięki przyspieszonej syntezie cyklicznego
adenozynomonofosforanu (cAMP) w komórkach kanalików nerkowych
zwiększa się resorpcja zwrotna wody w częściach dalszych kanalików
nerkowych i w kanalikach zbiorczych.
Wzrost ciśnienia osmotycznego krwi pobudza osmodetektory znajdujące się w okolicy jądra nadwzrokowego podwzgórza. Powoduje to
uwalnianie niewielkich ilości wazopresyny z części nerwowej przysadki
do krwi i zahamowanie utraty wody przez organizm. Jednocześnie zostaje
pobudzony ośrodek pragnienia w podwzgórzu, który kieruje aktywnością
somatyczną człowieka prowadzącą do wypicia wody, a tym samym do
obniżenia ciśnienia osmotycznego krwi.
W warunkach fizjologicznych impulsacja z receptorów objętościowych
i baroreceptorów układu sercowo-naczyniowego stale hamuje uwalnianie
wazopresyny do krwi.
Natomiast obniżenie ciśnienia tętniczego na skutek utraty krwi wywołuje wydzielanie do krwi znacznych ilości wazopresyny. Wydzielanie wazo-
230
Kontrola środowiska wewnętrznego
Ryc. 119. Cząsteczka wazopresyny argininowej
(AVP).
presyny w czasie obniżonego ciśnienia tętniczego zachodzi na skutek
braku impulsacji aferentnej z baroreceptorow zatok tętnic szyjnych i łuku
aorty oraz zwiększenia zawartości we krwi angiotensyny II. W tych
warunkach wazopresyna wydzielana do krwi kurczy mięśnie gładkie
naczyń krwionośnych, działając za pośrednictwem receptora V 1A. Tym
Czynność gruczołów dokrewnych
231
samym zwiększa całkowity opór naczyń obwodowych, podwyższa ciśnienie tętnicze i poprawia warunki krążenia krwi.
Mniejsza część aksonów neuronów wazopresynoergicznych biegnie
z jądra nadwzrokowego, jądra przy komorowego i jądra ponad skrzyżowaniem wzrokowym nie do tylnego płata przysadki, lecz do kory mózgu,
wzgórza, układu limbicznego, śródmózgowia, rdzenia przedłużonego
i rdzenia kręgowego. Neurony te uczestniczą w procesach związanych
z konsolidacją pamięci i z modulacją pobudliwości ośrodków regulujących
układ sercowo-naczyniowy.
W neuronach wazopresynoergicznych występują również transmittery
noradrenalina i dopamina oraz neuropeptydy: pochodne pre-pro-enkefaliny B (enkefalina leucynowa, alpha- i beta-neoendorfina, dynorfiny
1—8 i 1—17), kortykoliberyna, angiotensyna II i galanina.
Oksytocyna
Oksytocyna — OXY (oxytocin) — jest cyklicznym dziewięciopeptydem o masie cząsteczkowej 1007. Wydzielana jest do krwi z części
nerwowej przysadki na drodze odruchowej. Podrażnienie receptorów
brodawki sutkowej {papilla mammae) wywołuje wydzielanie oksytocyny,
która kurczy mięśnie przewodów mlecznych (ducti lactiferes) sutka
i wydala mleko w nich nagromadzone.
Podrażnienie receptorów w szyjce macicy (cervix uteri) i w pochwie
(vagina) również powoduje wydzielanie oksytocyny na drodze odruchowej. Występuje to w czasie porodu i w czasie aktu płciowego. Rozciągnięcie szyjki macicy w czasie akcji porodowej na drodze odru chowej powoduje wydzielanie dużych ilości oksytocyny, która z kolei,
działając na błonę mięśniową macicy, wywołuje jej skurcze i postęp akcji
porodowej.
W czasie aktu płciowego błona mięśniowa macicy (uterus) i jajowodu
(tuba uterina) kurczy się pod wpływem oksytocyny wydzielanej na drodze
odruchowej i przyspiesza transport spermatocytów przez jamę macicy
(canim uteri) i jajowód do bańki jajowodu (ampulla tubae uterinae), gdzie
następuje zapłodnienie jaja.
Hormony jajnika zmieniają pobudliwość błony mięśniowej macicy na
działanie oksytocyny. Estrogeny zwiększają pobudliwość macicy, zwiększając jednocześnie umiarkowanie ujemny potencjał wewnątrz komórek
mięśnia macicy, który osiąga od —35 do —45 mV. Progesteron zmniejsza
pobudliwość macicy, tak że pod wpływem oksytocyny macica nie kurczy
się. Zmniejszenie pobudliwości wiąże się ze znacznym wzrostem ujemnego
232
Kontrola środowiska wewnętrznego
potencjału wewnątrzkomórkowego do —60 mV, czyli z hiperpolaryzacją
błony komórkowej komórek mięśniowych.
W czasie ciąży, a zwłaszcza w czasie porodu, zwiększa się we krwi kobiet
ciężarnych zawartość oksytocynazy — enzymu rozkładającego oksytocynę. Porcje oksytocyny wydzielone do krwi na drodze odruchowej wywołują skurcz macicy i zostają szybko rozkładane przez oksytocynazę.
Wydzielanie kolejnych porcji oksytocyny co kilka lub kilkanaście minut
trwa w ciągu całego porodu.
Ryc. 121. Cząsteczka oksytocyny (OXY).
W neuronach oksytocynoergicznych koegzystują z oksytocyną neuropeptydy pochodne pre-pro-enkefaliny A (enkefalina metioninowa i leucynowa), cholecystokinina (CCK-8) i kortykoliberyna.
Hormony podwzgórzowe uwalniające i
hamujące
W podwzgórzu syntetyzowane są przekaźniki chemiczne, które zostają
wydzielone do krwi pierwotnej sieci naczyń włosowatych (rete hemocapillare primarium) w obrębie guza popielatego {tuber cinereum). Naczynia
włosowate sieci pierwotnej łączą się, tworząc przysadkowe naczynia
wrotne (vasa por talia hypophysis) biegnące wzdłuż lejka {infundibulum) od
pod wzgórza do części gruczołowej przysadki {adenohypophysis). Przysadkowe żyły wrotne dzielą się w przysadce na naczynia włosowate o dużym
świetle — zatoki żylne, stanowiące wtórną sieć naczyń włosowatych {rete
hemocapillare secundarium). Przekaźniki chemiczne, czyli hormony podwzgórzowe, wydzielone do pierwotnej sieci naczyń włosowatych po
przepłynięciu przez przysadkowe naczynie wrotne dostają się do wtórnej
sieci naczyń włosowatych i działają na komórki gruczołowe przysadki.
C z y n n oś ć gr uc z o ł ó w d ok r e w ny c h
233
Ryc. 122. Podwzgórze wraz z przysadką u człowieka.
Hormony podwzgorzowe działają pobudzająco, powodując wydzielanie hormonów przez przysadkę, oraz hamująco — zmniejszając lub
całkowicie hamując biosyntezę i uwalnianie do krwi hormonów przysadki.
Hormony podwzgorzowe działające pobudzająco to:
•
•
•
•
kortykoliberyna — CRH (corticotropin-releasing hormone);
tyreoliberyna — TRH (thyrotropin-releasing hormone);
gonadoliberyna — GnRH (gonadotropin-releasing hormone);
somatokrynina — GRH (growth hormone-releasing hormone).
234
Kontrola środowiska wewnętrznego
Hormony podwzgórzowe działające hamująco to:
• somatostatyna — SRIF (somatotropin release-inhibiting factor);
• prolaktostatyna — PIF (prolactin-inhibiting hormone).
Czynnik hamujący uwalnianie prolaktyny (PIF) nie jest polipeptydem.
Stanowią go aminy katecholowe, przede wszystkim dopamina, która
uwalniana jest z zakończeń neuronów dopaminergicznych do pierwotnej
sieci naczyń włosowatych w podwzgórzu.
Biosynteza i wydzielanie podwzgórzowych hormonów do przysadkowego układu wrotnego są kontrolowane przez:
• transmittery wydzielane na synapsach otaczających neurony wydzielnicze;
• hormony wydzielane przez zależne gruczoły dokrewne (kora nad
nerczy, gruczoł tarczowy, gruczoły płciowe), oddziałujące na pod-
Ryc. 123. Schemat ośrodkowej kontroli środowiska wewnętrznego za pośrednictwem
podwzgórza. Kierunek kontroli nerwowej oznaczony jest linią ciągłą, kierunek kontroli
humoralnej — linią przerywaną.
Czynność gruczołów dokrewnych
235
wzgórze na drodze zewnętrznego sprzężenia zwrotnego (external feedback control);
• hormony części gruczołowej przysadki wpływające na podwzgórze
dzięki wewnętrznemu sprzężeniu zwrotnemu (internal feedback con
trol);
• wytwarzane w podwzgórzu prostaglandyny zwiększające przepływ
krwi przez przysadkowe naczynia wrotne;
• inne bodźce ze środowiska wewnętrznego i zewnętrznego.
Komórki mikrogleju w podwzgórzu wytwarzają interleukinę 1 (IL-1) —
przekaźnik chemiczny o budowie polipeptydowej, która działa nieswoiście
na komórki części gruczołowej przysadki. Pobudza wydzielanie hormonu
kortykotropowego (ACTH), hormonu luteinizującego (LH) i hormonu
wzrostu (GH), hamuje natomiast wydzielanie prolaktyny (PRL).
W podwzgórzu i w okolicach z podwzgórzem sąsiadujących znajdują
się detektory, czyli komórki nerwowe szczególnie wrażliwe na krążące
we krwi cząsteczki hormonów gruczołów zależnych od przysadki oraz
hormony części gruczołowej przysadki. Znajdują się tam również detektory szczególnie wrażliwe na inne czynniki chemiczne, np. stężenie
glukozy lub innych związków we krwi, czy fizyczne, np. temperatura
krwi dopływającej do mózgu. W podwzgórzu konwerguje podrażnienie
detektorów z impulsacją aferentną biegnącą od receptorów całego ciała
i wysyłaną przez inne ośrodki mózgowe. W wyniku konwergencji podrażnienia detektorów i impulsacji nerwowej dochodzi do zwiększonego
lub zmniejszonego wydzielania poszczególnych hormonów przysadki.
I Przysadka — część gruczołowa
W części gruczołowej przysadki (adenohypophysis) występują komórki
wydzielające hormony:
• hormon wzrostu (hGH) — komórki somatotropowe (somatotropy);
• prolaktynę (PRL) — komórki prolaktynowe (laktotropy);
• hormon kortykotropowy (ACTH) — komórki kortykotropowe (kortykotropy);
• hormon tyreotropowy (TSH) — komórki tyreotropowe (tyreotropy);
• hormony gonadotropowe (hormon folikulotropowy — FSH i hormon
luteinizujący — LH ) — komórki gonadotropowe (gonadotropy)
oraz komórki macierzyste dla komórek wydzielających hormony.
Tabela 11
Hormony wydzielane przez komórki płata przedniego przysadki
TyreoKomórki gruczołowe:
Kortykotropowe
tropowe
Hormon:
Liczba aminokwasów
w cząsteczce
kortyko- P-lipotropowy tropowy
(ACTH) (p-LPH)
39
91
T V2 we krwi (min)
10
Kontrolowany uwalniaCRH
przez podjmy
wzgorzowy
hamuhormon
jący
kora
tkanka
Tkanki docelowe
gruczołu tłuszczonadner- wa
czowego
Gonadotropowe
Somatotropowe
Prolaktynowe
melanotropowy
(y-MSH)
tyreotropowy
(TSH)
folikulotropowy
(FSH)
luteinizujący
(LH)
wzrostu
(hGH)
prolaktyna
(PRL)
12
211
210
204
191
198
60
170
60
20—30
20—30
GRH
TRH
VIP
SRIF
dopamina
TRH
GnRH
SRIF
melanocyty w
skórze
gruczoł
tarczowy
jajniki:
pęcherzyki
jajnikowe
jądra: cewki
nasienne
jajniki:
pęcherzyki
jajnikowe
dojrzałe
jądra:
komórki
śródmiąższowe
komórki
komórki
całego orga- całego organizmu,
nizmu
wątroba
Czynność gruczołów dokrewnych
237
Hormon wzrostu
Ludzki hormon wzrostu — hGH (human growth hormone) — stanowi
łańcuch polipeptydowy składający się ze 191 aminokwasów o masie
cząsteczkowej 22 kDa.
W okresie pierwszych czterech lat życia wydzielanie hormonu zwiększa
się i u dzieci przed osiągnięciem dojrzałości płciowej wynosi na dobę
około 90 ug hGH, a po osiągnięciu dojrzałości płciowej około 690 |xg
hGH na dobę. U dorosłych w wieku 21 do 41 lat przysadka wydziela
około 385 ug hGH na dobę.
Zawartość hGH we krwi podlega znacznym wahaniom w ciągu doby.
Największa zawartość hGH we krwi występuje w nocy w pierwszych
godzinach snu. U ludzi powyżej 50 roku życia wydzielanie hGH zmniejsza się w ciągu doby. Dotyczy to również pulsacyjnego wydzielania
hGH z przedniego płata przysadki do krwi w czasie snu nocnego.
Hormon wzrostu (hGH) pobudza wątrobę, inne narządy i tkanki do
wydzielania czynników wzrostowych (growth factors), dawniej nazywanych somatomedynami. Zasadniczym czynnikiem wzrostowym wydzielanym pod wpływem hGH u ludzi jest czynnik wzrostowy insulinopodobny I — IGF-I (insulin-like growth factor I) i w znacznie mniejszym
stopniu czynnik wzrostowy insulinopodobny II — IGF-II (insulin--like
growth factor II). Zawartość hGH i IGF-I we krwi zwiększa się wraz
ze wzrostem organizmu i zmniejsza się w następstwie starzenia się.
Odróżnienie bezpośredniego działania hGH od działania pośredniego
poprzez IGF-I jest często niemożliwe. Z tego względu skutki obwodowe
związane z wydzielaniem hGH nie są bezpośrednim działaniem tego
hormonu, lecz aktywacją kaskady zaczynającej się w podwzgórzu i biegnącej przez przedni płat przysadki do tkanek.
Hormon wzrostu bierze udział w:
• syntezie białek organizmu;
• przemianie węglowodanów;
• przemianie tłuszczów;
• przemianie mineralnej.
Pod wpływem hormonu wzrostu (hGH) i czynnika wzrostowego
IGF-I dochodzi do przewagi procesów anabolicznych nad katabolicznymi, czego dowodem jest dodatni bilans azotowy organizmu.
Hormon wzrostu wzmaga transport aminokwasów do wnętrza komórek i syntezę białka komórkowego. Pod wpływem hGH w okresie
wzrostu organizmu chrząstki przynasadowe kości długich poszerzają się
i kości się wydłużają.
238
Kontrola środowiska wewnętrznego
Hormon ten zwiększa stężenie glukozy we krwi na skutek zahamowania syntezy glikogenu w mięśniach szkieletowych i zmniejszonego
zużycia glukozy. W wątrobie — przeciwnie — dochodzi do wzmożonej
glikoneogenezy i do zwiększenia zawartości glikogenu wątrobowego.
Hormon wzrostu, zwiększając stężenie glukozy we krwi, powoduje wtórne
wzmożone wydzielanie insuliny.
Hormon wzrostu ma działanie lipolityczne, tak że w kilkadziesiąt
minut po zwiększeniu zawartości hGH we krwi dochodzi także do zwiększenia stężenia krążących we krwi wolnych kwasów tłuszczowych (FFA).
Hormon ten zatrzymuje większość kationów, zwłaszcza zaś jony
wapniowe w postaci soli kwasu fosforowego. Czynniki takie, jak ból,
zimno, znaczny wysiłek fizyczny, głód, zmniejszenie stężenia glukozy
we krwi i zwiększenie zawartości aminokwasów we krwi, szczególnie
argininy, wywołują zwiększone wydzielanie hGH. Duże stężenie glukozy
we krwi i glikokortykoidy hamują wydzielanie hGH.
Podwzgórze reguluje uwalnianie hGH z części gruczołowej przysadki
za pośrednictwem dwóch hormonów o działaniu przeciwnym wydzielanych do przysadkowych naczyń wrotnych:
• hormonu uwalniającego hormon wzrostu z przysadki — somatokryniny
(GRH);
• hormonu hamującego uwalnianie hormonu wzrostu z przysadki — somatostatyny (SRIF).
Wraz ze starzeniem się organizmu w podwzgórzu uzyskuje przewagę
hormon hamujący (SRIF) nad hormonem uwalniającym (GRH) ludzki
hormon wzrostu (hGH) i stopniowo zmniejsza się z biegiem lat wydzielanie hGH.
U kobiet w czasie ciąży wytwarzana jest w syncytiotrofoblaście
somatomammotropina kosmówkowa ludzka — hCS (human chorionic
somatomammotropin). Hormon ten krąży głównie we krwi matki i tylko
w niewielkich ilościach przechodzi w łożysku do krwi płodu.
Prolaktyna
Prolaktyna (PRL) ma masę cząsteczkową 23,4 kDa i pod względem
chemicznym jest białkiem zbudowanym ze 199 aminokwasów, zbliżonym
do hormonu wzrostu.
Prolaktyna, podobnie jak hormon wzrostu, wzmaga syntezę białka
komórkowego w całym organizmie u obu płci. Wydzie lanie jej jest
stale hamowane przez uwalniany z podwzgorza do przysadkowych naczyń
Czynność gruczołów dokrewnych
239
wrotnych podwzgórzowy hormon hamujący uwalnianie prolaktyny
— PIH (prolactin-inhibiting hormone), którym jest dopamina. Wydzielanie
prolaktyny staje się bardziej aktywne w czasie snu, wysiłku fizycznego,
stresu fizycznego i psychicznego.
U kobiet w czasie ciąży wydzielanie prolaktyny zwiększa się, osiągając
największe stężenie we krwi przed porodem. Po porodzie stężenie to
zmniejsza się do wartości poprzedzających ciążę. W okresie laktacji
drażnienie receptorów w brodawce sutkowej przez ssącego oseska powoduje każdorazowo znaczny krótkotrwały wzrost wydzielania prolaktyny i wzmożoną syntezę białka wydzielanego z mlekiem z gruczołów
sutkowych.
Ryc. 124. Komórka kwasochłonna (laktotropowa) przysadki. 1 — ziarno wydzielnicze, 2 —
jądro, 3 — ziarnista siateczka śródplazmatyczna, 4 — mitochondrium, 5 — aparat Golgiego
wytwarzający ziarna wydzielnicze. (Reprodukowano za zezwoleniem Michała Karaska.)
U kobiet karmiących po porodzie, czyli w okresie laktacji, przez
PRL hamowane jest wydzielanie hormonów gonadotropowych (FSH i LH)
i nie dochodzi do owulacji, jak również nie ma cykli miesiączkowych.
Hamowanie uwalniania dopaminy z podwzgórza lub blokowanie
receptorów dopaminergicznych w błonie komórkowej komórek prolaktynowych przy użyciu preparatów farmakologicznych powoduje wydzielanie mleka przez gruczoły sutkowe u kobiet nie karmiących piersią.
240
Kontrola środowiska wewnętrznego
Hormon lipotropowy
W komórkach kortykotropowych części gruczołowej przysadki, w komórkach części pośredniej przysadki i w niektórych komórkach nerwowych w podwzgórzu dochodzi do ekspresji genu pre-pro-opiomelanokortyny — pre-POMC (pre-pro-opiomelanocortin), której łańcuch polipeptydowy zawiera 265 aminokwasów. W procesie potranslacyjnej enzymatycznej modyfikacji łańcuch polipeptydowy cięty jest na krótsze
łańcuchy stanowiące prekursory hormonów wydzielanych przez komórki.
Takim prekursorem dla innych hormonów jest hormon beta-lipotropowy
(beta-LPH — beta-lipotropin) zawierający 91 aminokwasów i hormon
gamma-lipotropowy — gamma-LPH — o jeszcze mniejszej liczbie aminokwasów.
Hormony lipotropowe (LPH) są więc prekursorami innych hormonów,
same prawdopodobnie nie stanowiąc nośnika informacji w organizmie
(tab. 12).
Hormony tropowe
Hormony tropowe: ACTH, TSH, FSH i LH wydzielane przez część
gruczołową przysadki kontrolują zależne gruczoły dokrewne. ACTH jest
małym polipeptydem mającym 39 aminokwasów w cząsteczce. TSH, FSH
i LH mają znacznie większe cząsteczki zbudowane z dwóch łańcuchów
polipeptydowych alpha i beta, w których łącznie występuje ponad 200
aminokwasów. Łańcuch alpha jest identyczny w cząsteczkach TSH, FSH
i LH, natomiast łańcuch beta jest inny w każdym z tych hormonów.
Hormony zależnych gruczołów dokrewnych: kory nadnerczy, gruczołu
tarczowego i gruczołów płciowych działają na detektory w podwzgórzu
i zwrotnie hamują sekrecję odpowiednich podwzgórzowych hormonów
uwalniających — CRH, TRH lub GnRH. Jest to mechanizm zewnętrznego sprzężenia zwrotnego.
Hormony kory nadnerczy, hormony płciowe i szczególnie hormony
gruczołu tarczowego działają również bezpośrednio na komórki wydzielnicze części gruczołowej przysadki.
Dzięki tym sprzężeniom zwrotnym i synergicznemu lub antagonistycznemu działanu hormonów na podwzgórze i część gruczołową przysadki
wydzielanie hormonów przez gruczoły dokrewne stale jest regulowane.
Hormony tropowe są wydzielane do krwi w sposób pulsacyjny, a więc
w niewielkich porcjach w odstępach od kilkunastu do kilkudziesięciu
minut i utrzymują zależne od przysadki gruczoły dokrewne na poziomie
Czynność gruczołów dokrewnych
241
czynności spoczynkowej. W czasie ostrego zapotrzebowania na hormony
przysadka wydziela duże ilości hormonów tropowych.
U kobiet od około 30 roku życia stopniowo zaczyna zwiększać się
stężenie hormonu folikulotropowego (FSH) w osoczu krwi, od około 35
roku życia zaś również zaczyna zwiększać się stężenie hormonu luteinizującego (LH) w osoczu krwi. U mężczyzn, w miarę starzenia się organizmu,
nie zaznacza się zwiększenie stężenia hormonów gonadotropowych (FSH
i LH) w osoczu krwi, ale występują zmiany w ich pulsacyjnym wydzielaniu
z części gruczołowej przysadki.
I Przysadka — część pośrednia
Komórki nabłonkowe części pośredniej przysadki (pars intermedia
hypophysis) wydzielają u ludzi hormony melanotropowe: alpha-MSH, beta-MSH, gamma-MSH (alpha-, beta-, gamma-melanocyte-stimulating hormone), fragment hormonu kortykotropowego (ACTH) o łańcuchu peptydowym od 18 do 39 aminokwasów, zwany również kortykotropowopodobnym peptydem części pośredniej przysadki — CLIP (corticotro-pinlike intermediate lobe peptide) — oraz alpha- lub beta-endorfinę.
Wszystkie te hormony powstają z pre-pro-opiomelanokortyny (pre-POMC).
Neurony dopaminergiczne podwzgórza wysyłają aksony biegnące
przez lejek do części pośredniej przysadki, stale hamując wydzielanie
Tabela 12
Potranslacyjna enzymatyczna modyfikacja prekursora pre-pro-opiomelanokortyny w ko-
W nawiasach podana jest liczba aminokwasów w cząsteczce polipeptydu.
mórkach części pośredniej przysadki
242
Kontrola środowiska wewnętrznego
hormonów. Natomiast brak hamowania ze strony neuronów dopaminergicznych wyzwala wydzielanie hormonów przez część pośrednią przysadki.
Hormon melanotropowy wywołuje u ludzi zmianę w rozmieszczeniu
melaniny w skórze oraz uwalnianie wolnych kwasów tłuszczowych z tkanki tłuszczowej.
U ludzi dorosłych część pośrednia przysadki wydziela tylko nieznaczne
ilości hormonów.
I Kora i rdzeń nadnerczy
Hormony kory nadnerczy
Hormony kory nadnerczy stanowią pochodne cholesterolu. Dzielą się
na trzy zasadnicze grupy różniące się pod względem budowy chemicznej,
roli w organizmie i mechanizmów kontrolujących ich wydzielanie. Hormony, których cząsteczki mają 21 atomów węgla, należą do grupy
glikokortykoidów i do grupy mineralokortykoidów. Trzecią grupę stanowią androgeny zawierające 19 atomów węgla w cząsteczce i grupę
ketonową przy 17 atomie węgla. W związku z tym grupę tę określa się
również jako 17-ketosteroidy.
W warunkach fizjologicznych kora nadnerczy wydziela tylko niewielkie ilości hormonów płciowych należących do grupy estrogenów i progestagenów.
ACTH wydzielane przez część gruczołową przysadki aktywuje adenylocyklazę w komórkach kory nadnerczy. W komórkach tych zwiększa
się zawartość cyklicznego AMP i dochodzi do przemian cholesterolu.
W procesie przemian cholesterolu bierze również udział kwas askorbinowy.
Glikokortykoidy
Zasadnicze hormony glikokortykoidowe to kortyzol i kortykosteron
Wydzielane do krwi wiążą się z białkami osocza. Kortyzol wiąże się
z białkiem należącym do frakcji alpha-globulin, czyli transkortyną. lub
globuliną wiążącą kortyzol — CBG (corticosteroid-binding globulin).
Tylko niewielki procent kortyzolu i kortykosteronu krąży we krwi
w postaci wolnej, nie związanej z białkiem. Występuje stała równowaga we
Czynność gruczołów dokrewnych
243
krwi pomiędzy hormonami związanymi z białkiem — nieczynnymi, jak
gdyby w ten sposób zmagazynowanymi, a hormonami wolnymi —
aktywnymi. Hormony nie związane z białkami osocza są fizjologicznie
aktywne.
Zawartość wolnych glikokortykoidów we krwi jest regulowana dzięki:
• wydzielaniu ACTH przez część gruczołową przysadki;
• wazopresynie uwalnianej do sieci pierwotnej przysadkowych naczyń
wrotnych i działającej za pośrednictwem receptora V 1B na komórki
kortykotropowe części gruczołowej przysadki;
• wytwarzaniu transkortyny przez wątrobę.
Zwiększona zawartość CBG we krwi zmniejsza stężenie wolnych
glikokortykoidów krążących we krwi i zwrotnie zwiększa wydzielanie
ACTH. Przy zmniejszonej zawartości CBG we krwi stężenie wolnych
glikokortykoidów zwiększa się i zwrotnie hamuje wydzielanie ACTH.
Glikokortykoidy metabolizowane są w wątrobie i wydalane z moczem
w przewadze jako metabolity związane z resztą kwasu glukuronowego
lub jako 17-ketosteroidy związane z resztą kwasu siarkowego.
Glikokortykoidy oddziałują na metabolizm węglowodanów, białek i tłuszczów w tkankach całego organizmu. W wątrobie przyspieszają syntezę
glikogenu i jednocześnie aktywują glukozo-6-fosfatazę. Dzięki temu
zawartość glukozy we krwi zwiększa się.
y
koidy
Glikokortykoidy
związane z białkiem
Ryc. 125. Regulacja stężenia wolnych glikokortykoidów krążących w osoczu krwi.
244
Kontrola środowiska wewnętrznego
Tabela 13
Hormony wydzielane przez korę gruczołu nadnerczowego
Grupa
Zawartość w 1 L
Okres poło- Ilość wydzielana
Hormon
hormonów
osocza krwi
wicznego
na dobę
obwodowej
rozpadu we
krwi TV,
(min)
Glikokortykoidy
kortyzol
kortykosteron
Mineralokortykoidy
aldosteron 11 deoksykortykosteron
dehydroepiandrosteron
Androgeny
Estrogeny
Progestydy
423 nmol/L (140 ug)
13 nmol/L
(4 ug)
20 pmol/L
(70 ng)
20 pmol/L (60 ng)
6,25 umol/L(l,75mg)
estradiol
progesteron
60—90
50
20
30 umol/d (10 mg)
9,8 umol/d (3 mg)
431 pmol/d(150ug)
622 pmol/d (200 ug)
mężczyźni: 58
umol/d (15 mg)
kobiety: 39 umol/d
(10 mg)
ilości śladowe
ilości śladowe
W nawiasach podane są te same wielkości wyrażone w miligramach i mikrogramach.
Glikokortykoidy utrzymują prawidłową pobudliwość mięśni poprzecznie prążkowanych szkieletowych, mięśni gładkich i mięśnia sercowego.
Zwiększają wydzielanie soku żołądkowego i zwiększają w nerkach przesączanie kłębuszkowe — GFR (glomerular filtration ratę), współdziałając
w wydalaniu wody z organizmu. Zmniejszają liczbę krążących we krwi
obwodowej granulocytów kwasochłonnych, dzięki zatrzymywaniu ich
w narządach wewnętrznych (płuca, śledziona), oraz limfocytów w wyniku
hamowania ich podziałów i przyspieszonego rozpadu. Węzły
chłonne i grasica pod wpływem glikokortykoidów zmniejszają swoje
rozmiary, ponieważ część komórek ginie w tkankach limfoidalnych,
szczególnie pod działaniem kortyzolu.
Mineralokortykoidy
Najważniejszy w grupie mineralokortykoidow jest aldosteron. W przeciwieństwie do glikokortykoidów wydzielanie jego jest regulowane w przewadze przez inne czynniki poza ACTH.
Czynniki zwiększające wydzielanie mineralokortykoidow to:
Czynność gruczołów dokrewnych
245
• zwiększenie zawartości we krwi angiotensyny II jako skutek zwięk
szenia zawartości we krwi reniny uwolnionej z aparatu przykłębuszkowego w nerkach po znacznym obniżeniu się ciśnienia tętniczego
krwi lub zmniejszeniu całkowitej objętości krwi krążącej;
• zwiększenie we krwi koncentracji jonów potasowych i zmniejszenie się
koncentracji jonów sodowych;
• zwiększenie wydzielania ACTH przez część gruczołową przysadki.
Mineralokortykoidy zwiększają w częściach dalszych kanalików nerkowych resorpcję zwrotną jonów sodowych z moczu pierwotnego i jednocześnie zwiększają wydzielanie jonów potasowych. Podobnie działają
na ślinianki i gruczoły w błonie śluzowej żołądka, zatrzymując jony
sodowe w organizmie i zwiększając objętość płynu zewnątrzkomórkowego. W komórkach mięśniowych i nerwowych mineralokortykoidy
zwiększają zawartość potasu i jednocześnie zmniejszają zawartość sodu.
Zwiększony aktywny transport sodu w nabłonku części dalszych
kanalików nerkowych, wbrew gradientowi stężenia, wymaga energii
dostarczanej przez ATP. Aldosteron przyspiesza syntezę w jądrze komórkowym mRNA, który z kolei po przesunięciu się do cytoplazmy wzmaga
syntezę białek w rybosomach. Wytworzone w rybosomach białka komórkowe przyspieszają resyntezę ATP z ADP. Komórka szybciej uzupełnia energię niezbędną do aktywnego transportu sodu, tym samym
proces ten szybciej przebiega.
Androgeny
Zasadniczym hormonem grupy androgenów wydzielanym do krwi
jest dehydroepiandrosteron — DHEA (dehydroepiandrosterone). Do jego
pochodnych należą również testosteron i estradiol wytwarzane w nieznacznych ilościach przez korę nadnerczy w warunkach prawidłowych.
W wyniku zaburzeń metabolicznych może dojść do wzmożonej syntezy
testosteronu lub estradiolu w korze nadnerczy.
Pod wpływem androgenów przyspieszeniu ulega synteza białek i wzrost
organizmu oraz rozwijają się niektóre drugorzędowe cechy płciowe
o typie męskim.
Wydzielanie dehydroepiandrosteronu (DHEA) — zarówno u kobiet,
jak i u mężczyzn — jest największe w wieku około 20 lat. W miarę
przybywania lat stężenie DHEA w osoczu krwi zmniejsza się u obu płci
o około 3% każdego roku.
246
Kontrola środowiska wewnętrznego
Hormony rdzenia nadnerczy
Rdzeń nadnerczy (medulla glandulae suprarenalis) wydziela aminy
katecholowe. Najwięcej, bo w około 80%, adrenaliny (A — epinephrine),
w około 20% noradrenaliny (NA — norepinephrine) i najmniej dopaminy. Adrenalina powstaje w cyklu przemian z aminokwasu fenyloalaniny poprzez tyrozynę, DOPA, dopaminę i noradrenalinę. W warunkach
przeciętnych życia organizmu w osoczu krwi stale znajduje się więcej
noradrenaliny (4,7 nmol/L osocza)w porównaniu z adrenaliną (0,5 nmol/L
osocza), ponieważ zasadniczym źródłem noradrenaliny są zakończenia
zazwojowe komórek nerwowych układu współczulnego.
Ryc. 126. Gromadzenie i wydzielanie z pęcherzyków rdzenia nadnerczy adrenaliny i noradrenaliny.
Adrenalina i noradrenalina są syntetyzowane i magazynowane w pęcherzykach komórek rdzenia nadnerczy, gdzie pozostają związane z adenozynotrifosforanami i białkiem — chromograniną A. W czasie pobudzenia komórki pęcherzyki, stykając się od wewnątrz z błoną komórkową, pękają i ich treść zostaje usunięta na zewnątrz na zasadzie
mechanizmu egzocytozy. Następnie puste pęcherzyki zamykają się i odsuwają od błony komórkowej. Następuje ponowne ich wypełnienie
hormonami.
W komórkach chromochłonnych rdzenia nadnerczy jest również syntetyzowany polipeptyd — pre-pro-enkefalina, w którego łańcuchu występuje 267 aminokwasów. W wyniku obróbki enzymatycznej z jednej
cząsteczki prekursora powstaje 6 cząsteczek enkefaliny metioninowej
i 1 cząsteczka enkefaliny leucynowej. Wydzielone enkefaliny wraz z aminami katecholowymi krążą we krwi i prawdopodobnie działają modulujące*
Czynność gruczołów dokrewnych
247
na drogi oraz ośrodki przewodzące i odbierające czucie bólu, choć
dotychczas nie wykazano przechodzenia enkefaliny przez barierę mózgową pomiędzy krwią i tkanką nerwową.
Wydzielanie hormonów rdzenia nadnerczy do krwi odbywa się pod
wpływem impulsacji biegnącej przez nerwy trzewne. Przedzwojowe neurony współczulne o zakończeniach cholinergicznych unerwiają rdzeń nadnerczy.
Pod wpływem acetylocholiny, wydzielonej na zakończeniach włókien
typu B, do wnętrza komórek rdzenia nadnerczy wnikają jony wapniowe,
które wywołują egzocytozę pęcherzyków i wydzielanie A i NA. Ośrodki
nerwowe bezpośrednio kontrolujące wydzielanie hormonów przez rdzeń
nadnerczy znajdują się w rogach bocznych istoty szarej w części piersiowej
rdzenia kręgowego, ośrodki nadrzędne zaś są w podwzgórzu.
Bodźcami zwiększającymi wydzielanie adrenaliny i noradrenaliny
przez rdzeń nadnerczy są: • obniżenie ciśnienia tętniczego krwi na skutek
utraty krwi i zmniejszenia
całkowitej objętości krwi;
Ryc. 127. Molekularny mechanizm wydzielania hormonów rdzenia gruczołu nadnerczowego:
adrenaliny (A) i noradrenaliny (NA).
248
Kontrola środowiska wewnętrznego
• zmniejszenie zawartości glukozy we krwi wywołane działaniem egzo
gennej insuliny;
• zmniejszenie prężności tlenu (Po2) we krwi tętniczej;
• obniżenie temperatury ciała;
• czynniki emocjonalne.
Hormony rdzenia nadnerczy krążące we krwi przenikają do tkanek,
w których są następnie metabolizowane, a ich pochodne związane z resztą
kwasu siarkowego lub glukuronowego są wydzielane z moczem. We
wszystkich tkankach występują enzymy, szczególnie aktywne w wątrobie:
tlenowa O-metylotransferaza katecholowa — COMT (catechol-O-methyltransferase), i oksydaza monoaminowa — MAO (monoamine oxidase).
Enzymy te metabolizują noradrenalinę i adrenalinę poprzez 3-metoksynoradrenalinę i 3-metoksyadrenalinę do kwasu 3-metoksy-4-hydroksymigdałowego — VMA (vanilmandelic acid).
Rola hormonów rdzenia nadnerczy
Hormony rdzenia nadnerczy wywierają swój wpływ na komórki innych
tkanek, działając poprzez alpha- i beta-receptory adrenergiczne znajdujące
się w ich błonie komórkowej. Adrenalina, która jest zasadniczym hormonem rdzenia nadnerczy, działa jednocześnie na alpha- i beta-receptory
i ostateczny wynik wywołany przez nią jest wypadkową jej działania na oba
rodzaje receptorów.
Czynność gruczołów dokrewnych
249
Adrenalina wydzielona do krwi przez rdzeń nadnerczy wywołuje:
rozszerzenie naczyń krwionośnych w mięśniach szkieletowych oraz
zwężenie naczyń w skórze, w błonach śluzowych i w narządach jamy
brzusznej;
przyspieszenie częstości skurczów serca, zwiększenie pojemności wyrzutowej serca i podwyższenie skurczowego ciśnienia tętniczego krwi;
rozkurcz mięśni gładkich w ścianach przewodu pokarmowego, oskrzeli
i pęcherza moczowego, działając na beta receptory adrenergiczne;
zwiększenie stężenia glukozy we krwi, działając glikogenolitycznie na
komórki wątrobowe poprzez cyklazę adenylanową, cykliczny AMP,
kinazę defosforylazy i fosforylazę;
przyspieszenie rozpadu triacylogliceroli w tkance tłuszczowej i przechodzenie do krwi wolnych kwasów tłuszczowych; pobudzenie
ośrodkowego układu nerwowego, szczególnie ośrodków w
podwzgórzu w wyniku aktywacji cyklazy adenylanowej w błonie
komórkowej neuronów i tworzenie się w komórkach nerwowych
cyklicznego AMP.
I Gruczoł tarczowy
Zasadniczym hormonem gruczołu tarczowego, czyli tarczycy, jest
tyroksyna — T 4 (thyroxine) — aminokwas zawierający w cząsteczce
cztery atomy jodu. Drugim hormonem wydzielanym w mniejszej ilości
jest trijodotyronina — T3 (triiodothyronine) — aminokwas z wbudowanymi trzema atomami jodu.
Ryc. 129. Pęcherzyk gruczołu tarczowego (a) i ultrastruktury komórki gruczołowej (6).
W komórce gruczołowej znajduje się dobrze rozwinięty aparat Golgiego i rozszerzone
cysterny ziarnistej siateczki śródplazmatycznej, na powierzchni komórki zwróconej do
wnętrza pęcherzyka widoczne zaś są mikrokosmki. (Reprodukowano za zezwoleniem
Michała Karaska.)
250
Kontrola środowiska wewnętrznego
Czynność gruczołu tarczowego związana jest z:
• wychwytywaniem jodu nieorganicznego i aminokwasu tyrozyny z krwi
krążącej przez komórki nabłonka pęcherzyków tarczycy;
• syntezą trijodotyroniny (T 3 ) i tyroksyny (T 4 ) wewnątrz komórek
nabłonka pęcherzyków tarczycy;
• magazynowaniem hormonów T 3 i T 4 wewnątrz pęcherzyków tarczycy
w postaci związanej z tyreoglobuliną;
• uwalnianiem do krwi hormonów T 3 i T 4 w wyniku odczepiania ich
od tyreoglobułiny w komórkach nabłonka pęcherzyków tarczycy.
Jod nieorganiczny pochodzący ze spożywanych pokarmów w ilości do
około 500 ug na dobę wchłaniany jest w przewodzie pokarmowym i krąży
we krwi utrzymując koncentrację 3 ng/L osocza. W około 80% jest on
wydalany przez nerki z moczem. Resztę jodu nieorganicznego wychwytują
komórki gruczołu tarczowego, gruczołów sutkowych, gruczołów błony
śluzowej żołądka, ślinianek i w niewielkich ilościach komórki innych
narządów. Wśród komórek wychwytujących z krwi jod nieorganiczny
tylko komórki gruczołu tarczowego wiążą go z cząsteczkami aminokwasów.
W PĘCHERZYKU
Ryc. 130. Biosynteza hormonów gruczołu tarczowego (T 3 i T 4 ) z tyrozyny i jodu (I) i
uwalnianie do krwi po degradacji tyreoglobułiny.
Czynność gruczołów dokrewnych
251
Wychwytywanie jodu nieorganicznego przez tarczycę jest procesem
aktywnym. Komórki nabłonka pęcherzyków tarczycy, których wnętrze
jest ujemne — 50 mV, osiągają 25 razy większe stężenie jodu w porównaniu z osoczem. Wychwytywanie ujemnie naładowanych anionów jodowych, wbrew gradientowi stężeń, określa się jako pompę jodową (iodide
pump).
W komórkach nabłonka, pęcherzyków jod jest utleniany i wbudowany
do cząsteczek tyrozyny — aminokwasu wchodzącego w skład tyreoglobuliny. Tyreoglobulina jest glikoproteidem o dużej masie cząsteczkowej
(około 660 kDa) tworzącym koloid wypełniający pęcherzyki.
W wyniku wbudowania jodu do tyrozyny powstaje monojodotyrozyna — MIT, która po przyłączeniu drugiego atomu jodu zmienia się
w dijodotyrozynę — DIT. Dwie cząsteczki DIT po połączeniu się ze
sobą i odczepieniu alaniny tworzą tetrajodotyroninę — T 4, czyli tyroksynę. Trijodotyronina powstaje z połączenia jednej cząsteczki monojodotyrozyny z jedną cząsteczką dijodotyrozyny.
Magazynowanie hormonów gruczołu tarczowego w koloidzie pęcherzyków polega na tym, że cząsteczki T 4 i T 3 wchodzą w skład dużej cząsteczki tyreoglobuliny.
Uwalnianie T4 i T3 z tyreoglobuliny zachodzi w komórkach nabłonka
pęcherzyków. Komórki te dzięki endocytozie wchłaniają do wnętrza
cząsteczki tyreoglobuliny. W lizosomach komórek nabłonka cząsteczki
tyreoglobuliny są trawione przez enzymy proteolityczne. Wolne aminokwasy, a wśród nich MIT, DIT, T 3 i T 4, przechodzą do cytoplazmy.
Trijodotyronina i tyroksyna wchodzą do krwi, monojodotyrozyna i dijodotyrozyna zaś zostają pozbawione jodu pod wpływem enzymu dehalogenazy jodotyrozynowej (iodotyrosine dehalogenase). Odczepiony jod
jest utleniany i w tej postaci powraca do wnętrza pęcherzyków.
Transport hormonów gruczołu
tarczowego
U dorosłego człowieka gruczoł tarczowy wydziel a do krwi około
100 ug tyroksyny na dobę i około 10 ug trijodotyroniny na dobę, które
wiążą się z białkiem osocza, tworząc tzw. jod związany z białkiem — PBI
(protein-bound iodine). Jod tyroksyny stanowi 95% jodu związanego
z białkiem.
Białka osocza wiążące hormony gruczołu tarczowego (PBI) są to: •
globulina wiążąca tyroksynę — TBG (thyroxine-binding globulin);
252
Kontrola środowiska wewnętrznego
• prealbumina wiążąca tyroksynę — TBPA (thyroxine-binding prealbumin);
• albumina.
Najwięcej tyroksyny związane jest z globuliną (TBG), znacznie mniej
z prealbumina (TBPA) i najmniej z albuminą.
Tyroksyna ma okres połowicznego rozpadu (T x/2) około 6—7 dni,
prawie cała jest związana z białkami osocza, tworząc nieaktywną PBI.
Trijodotyronina ma znacznie krótszy okres połowicznego rozpadu —
2 dni, i jest znacznie słabiej związana z białkami osocza. Działanie T 3 na
komórki organizmu jest silniejsze i krótkotrwałe, T 4 zaś słabsze i dhiżej
trwające.
We krwi krąży 52 pmol/L osocza wolnej tyroksyny i 115 nmol/L
osocza tyroksyny związanej z białkami krwi jako PBI. Wolna tyroksyna
bierze pośredni i bezpośredni udział w regulacji czynności gruczołu
tarczowego.
Maksymalne działanie związane z wprowadzeniem egzogennej tyroksyny do organizmu występuje po czterech dniach. Działanie trijodotyroniny jest znacznie szybsze. Maksymalne staje się już w ciągu 24 godzin
po wprowadzeniu.
Regulacja wydzielania hormonów gruczołu
tarczowego
Wydzielanie T 3 i T 4 przez gruczoł tarczowy jest regulowane bezpośrednio i pośrednio.
Do czynników zwiększających wydzielanie hormonów gruczołu tarczowego należą:
• hormon tyreotropowy (TSH) z części gruczołowej przysadki — wzmaga
wychwytywanie jodu przez gruczoł tarczowy i wiązanie jodu z biał
kiem oraz przyspiesza proteolizę tyreoglobuliny i uwalnianie do krwi
wolnych T3 i T4;
• podwzgórzowy hormon uwalniający hormon tyreotropowy z przysadki
(TRH) — działa na gruczoł tarczowy pośrednio poprzez TSH;
• zimno — nieznaczne obniżenie temperatury krwi dopływającej do
podwzgórza pobudza termodetektory w ośrodku termoregulacji. Pod
wpływem termodetektorów podwzgórze wydziela TRH, który —
uwalniając z przysadki TSH — działa w ten sposób na gruczoł tar
czowy;
Czynność gruczołów dokrewnych
253
wazopresyna, adrenalina i inne hormony oddziałujące na naczynia
krwionośne — uwalniają z gruczołu tarczowego T 3 i T 4. Wpływ ich
jest dwufazowy. W pierwszej fazie zwiększają wydzielanie T 3 i T 4
do krwi. Z kolei wolne T 3 i T 4 krążące we krwi hamują tworzenie
się TSH i tą pośrednią drogą hamują czynność gruczołu tarczowego.
Wydzielanie hormonów przez gruczoł tarczowy jest hamowane przez:
długotrwały wzrost średniej temperatury otoczenia i nieznaczny wzrost
temperatury krwi dopływającej do podwzgórza; zwiększenie
zawartości we krwi wolnych T 3 i T4 , które działają bezpośrednio
na część gruczołową przysadki lub pośrednio poprzez detektory w
podwzgórzu, hamując wydzielanie TSH; jod nieorganiczny
wprowadzony do organizmu w ilości ponad dziesięć razy większej niż w
warunkach prawidłowych (norma do 500 ug na dobę);
niedobór jodu nieorganicznego w pokarmach poniżej 20 ug na dobę;
inne aniony jednowartościowe, takie jak chlorany i azotany, które
współzawodnicząc wypierają aniony jodowe wychwytywane przez
gruczoł tarczowy z krwi;
związki egzogenne, np. propylotiouracyl, blokują wiązanie jodu z tyrozyną i powstawanie MIT i DIT.
254
Kontrola środowiska wewnętrznego
Rola hormonów gruczołu tarczowego
Wolne T 3 i T 4 po wniknięciu do wnętrza komórek w całym organizmie stymulują syntezę białek komórkowych, w tym również białek
enzymatycznych. Bezpośrednio z receptorem jądrowym wiąże się T 3,
której jest mniej, natomiast T 4 wnikająca do komórek w większej ilości
zostaje pozbawiona jednego atomu jodu i metabolizowana do trijodotyroniny — T3 (3, 5, 3'-triiodothyronine) lub do rewers trijodotyroniny — rT 3 (3, 3', 5'-triiodothyronine). Trijodotyronina (T 3) powstaje
wtedy, kiedy cząsteczka tyrozyny (T 4) zostaje pozbawiona atomu jodu
w pierścieniu aromatycznym dalszym w stosunku do łańcucha bocznego.
Rewers trijodotyronina (rT 3 ) tworzy się po odłączeniu atomu jodu
od pierścienia aromatycznego bliższego w stosunku do łańcucha bocznego.
Powstająca rT 3 w komórce nie tylko nie wykazuje aktywności T 3,
lecz hamuje powstawanie T 3 z T 4. W ten sposób w komórkach całego
organizmu zachodzi zwrotne hamujące przeciwdziałanie skierowane przeciwko hormonom gruczołu tarczowego. Czynniki powodujące metabolizowanie się T 4 do T 3 lub do rT 3 nie zostały dotychczas poznane.
Wyniki działania hormonów gruczołu tarczowego to:
• zwiększenie zapotrzebowania na tlen. W związku ze zwiększonym
zużyciem tlenu jego prężność (Po2) w tkankach słabnie;
• przyspieszenie spalania wewnątrzkomórkowego. Wiąże się z tym wy
twarzanie ciepła (calorigenic action) i wzmożona podstawowa prze
miana materii — BMR (basal metabolic ratę);
• zwiększone wydzielanie hormonu wzrostu w wyniku ich bezpośredniego
działania na część gruczołową przysadki;
• wzmożona synteza białek w organizmie wtedy, kiedy T 3 i T 4 są
wydzielane w ilościach fizjologicznych. Wydzielanie nadmierne tych
hormonów (nadczynność tarczycy) przyspiesza rozpad białek i prowa
dzi do ujemnego bilansu azotowego w organizmie;
• wzmożona resorpcja węglowodanów w jelitach, w komórkach zaś
przyspieszenie glikogenolizy;
• wzmożona synteza i rozpad cholesterolu w komórkach wątrobowych
oraz jego wychwytywanie z krwi. Tym samym T 3 i T 4 zmniejszają
stężenie cholesterolu w osoczu krwi;
• zwiększona przemiana wodno-mineralna. T3 i T4 przyspieszają wydala
nie z organizmu wody i soli mineralnych. W przypadku niedoboru
tych hormonów dochodzi do zatrzymania soli mineralnych, a szcze
gólnie jonów sodowych w organizmie.
Czynność gruczołów dokrewnych
255
Kalcytonina
W komórkach przypęcherzykowych gruczołu tarczowego (parafollicular cells) wytwarzany jest hormon peptydowy — kalcytonina (calcitonin). Zwiększona zawartość jonów wapniowych we krwi pobudza te
komórki do wydzielania kalcytoniny, zwrotnie zmniejszającej zawartość
jonów Ca 2+ we krwi.
Ludzka kalcytonina jest łańcuchem peptydowym o 32
aminokwasach i masie cząsteczkowej około 3,5 kDa.
Kalcytonina działa bezpośrednio na kości, zmniejsza w nich zawartość cyklicznego AMP, hamując tą drogą resorpcję wapnia i odwap nienie kości. Działa antagonistycznie w stosunku do hormonu gruczołów przytarczycznych — PTH, zwiększającego zawartość cyklicznego
AMP w kościach i przyspieszającego uwalnianie jonów wapniowych
z kości do krwi.
I Jajniki
Hormony jajnika (ovarium) dzielą się na dwie grupy: estrogeny i progesteron. Obie te grupy są pochodnymi cholesterolu.
Estrogeny powstają w pęcherzykach jajnikowych drugorzędowych
dojrzewających (Graafa), są syntetyzowane w komórkach warstwy ziarnistej (stratum granulosum) i w komórkach osłonki wewnętrznej (theca
interna) pęcherzyka jajnikowego (folliculus ovaricus) (patrz Rozród u płci
żeńskiej, str. 501).
Zasadniczymi hormonami należącymi do grupy estrogenów są: estradiol, estron i estriol. W zależności od fazy cyklu miesiączkowego wydzielanie estrogenów zwiększa się lub zmniejsza. Najwięcej estrogenów
wydziela się w fazie folikularnej, od 4 do 12 dnia cyklu miesiączkowego,
oraz w drugiej połowie fazy lutealnej. 19—25 dnia cyklu, osiągając od 200
do 500 ug na dobę.
Z całkowitej ilości estrogenów występujących w osoczu krwi tylko
około 2% stanowią wolne estrogeny. Pozostałe 98% estrogenów jest
związane z białkami osocza: w około 60% z albuminą i w około 38%
z globuliną wiążącą steroidy płciowe — GBG (gonadal steroid-binding
globulin). Wolne estrogeny przenikają przez błonę komórkową, wiążą się
z receptorem cytoplazmatycznym i oddziałują na aparat genetyczny jądra
komórkowego.
256
Kontrola środowiska wewnętrznego
Działanie estrogenów na:
błonę śluzową macicy — wywołują jej rozrost w fazie folikularnej cyklu
miesiączkowego, w fazie lutealnej zaś pobudzają gruczoły śluzowe
macicy do wydzielania śluzu;
błonę mięśniową macicy i jajowodu — powodują przerost mięśni
gładkich, zwiększają ich pobudliwość i ukrwienie. W czasie dojrzewania płciowego estrogeny wydzielone do krwi przez pęcherzyki jajnikowe drugorzędowe zwiększają ujemny potencjał wnętrza komórek
mięśniowych gładkich macicy. Zostaje przekroczony potencjał progowy błony komórkowej i mięsień macicy staje się wrażliwy na działanie oksytocyny.
drugorzędne cechy płciowe — wpływają na ich rozwój, powiększa się
pochwa, wzrastają gruczoły sutkowe, dochodzi do odkładania się
barwnika w naskórku wokół brodawek sutkowych i narządów płciowych zewnętrznych, gruczoły łojowe zwiększają wydzielanie rzadkiego
łoju;
ośrodkowy układ nerwowy — wyzwalają popęd płciowy w kierunku
płci męskiej działając na ośrodki motywacyjne w mózgowiu.
Ryc. 132. Regulacja wydzielania estrogenów w fazie folikularnej.
Czynność gruczołów dokrewnych
257
Progesteron jest wytwarzany przez komórki ciałka żółtego w jajnikach
w fazie lutealnej cyklu miesiączkowego. Powstaje pod wpływem LH,
w czym pośredniczy cykliczny AMP. Progesteron działa na macicę
antagonistycznie w stosunku do estrogenów. Zmniejsza pobudliwość
błony mięśniowej macicy, zwłaszcza w stosunku do oksytocyny, hiperpolaryzując błonę komórkową komórek mięśni gładkich. W obrębie
gruczołów sutkowych wywołuje w płacikach rozwój komórek gruczołowych.
Regulacja wydzielania hormonów jajnikowych
Dojrzewanie pęcherzyków jajnikowych drugorzędowych zachodzi pod
wpływem hormonu folikulotropowego (FSH) przysadki. Jego wydzielanie
jest zwrotnie hamowane przez inhibinę — hormon wytwarzany przez
komórki warstwy ziarnistej pęcherzyków jajnikowych wzrastających.
Owulacja występuje na skutek wzmożonego wydzielania przez przysadkę
jednocześnie obu gonadotropin: hormonu folikulotropowego (FSH) i hormonu luteinizującego (LH) ze znaczną przewagą tego ostatniego.
Zarówno estrogeny, jak i progesteron — krążące we krwi jako wolne
hormony — są w wątrobie utleniane lub łączone z resztą kwasu glukuronowego lub siarkowego. W tej postaci wydzielane są z żółcią do
dwunastnicy. Progesteron zamieniany jest w komórkach wątrobowych na
pregnandiol, który wiązany z resztą kwasu glukuronowego jest wydzielany
z żółcią do dwunastnicy. Metabolity estrogenów i progesteronu są
w jelitach częściowo wchłaniane do krwi, a następnie wydalane z moczem.
Zanikanie ciałka żółtego w końcu fazy lutealnej cyklu miesiączkowego
jest związane z wytwarzaniem się w macicy tzw. luteolitycznego czynnika
macicznego (uterine luteolysin). Ten uwalniany do krwi przez macicę
czynnik stanowią prostaglandyny, przede wszystkim prostaglandyna
PGF2a (patrz rozdz. 9).
I Jądra
Zasadniczym hormonem wydzielanym przez komórki śródmiąższowe
(Leydiga) w jądrach (testes) jest testosteron. Jego cząsteczka ma 19 atomów węgla i jest pochodną cholesterolu. Ogniwami pośrednimi w przemianie cholesterolu w testosteron są hydroksypregnenolon i progesteron
(patrz Rozród u płci męskiej, str. 499).
9 — Fizjologia człowieka
258
Kontrola środowiska wewnętrznego
Wydzielanie testosteronu jest kontrolowane przez hormon luteinizujący
(LH). Po związaniu się hormonu luteinizującego z receptorem błonowym
komórek śródmiąższowych jądra dochodzi do aktywacji cyklazy adenylanowej, zwiększenia stężenia cyklicznego AMP w tych komórkach
i zwiększonego wydzielania testosteronu. U dorosłych mężczyzn jądra
wydzielają od 14 do 31 umoli (4—9 mg) testosteronu w ciągu doby.
Testosteron krąży we krwi: w 65% związany z beta globuliną wiążącą
steroidy płciowe (GBG), w 33% związany z albuminą osocza i tylko
w około 2% w postaci wolnego hormonu. Zawartość całkowita testosteronu we krwi u mężczyzn wynosi średnio około 18,2 nmol/L (5,24 ug/L)
osocza. Testosteron występuje również we krwi u dorosłych kobiet, ale
w znacznie mniejszych ilościach, średnio około 1 nmol/L (0,3 ug/L)
osocza. Jest on wytwarzany przez korę nadnerczy.
Krążący we krwi testosteron dostaje się do wątroby, gdzie jest
zamieniany na 17-ketosteroidy wydalane z organizmu przez nerki wraz
z moczem. Dwie trzecie 17-ketosteroidów wydalanych z moczem są to
pochodne androgenów z kory nadnerczy, a tylko jedną trzecią 17-ketosteroidów stanowią metabolity testosteronu powstającego w jądrach.
Zwiększenie zawartości testosteronu we krwi powoduje pobudzenie
detektorów w podwzgórzu, zmniejszenie ilości wydzielanego do przy-
Czynność gruczołów dokrewnych
259
sadkowych naczyń wrotnych podwzgórzowego hormonu uwalniającego
gonadotropiny (GnRH) i hamowanie wydzielania hormonu luteinizującego (LH) przez przysadkę. Tą drogą zewnętrznego ujemnego sprzężenia zwrotnego regulowane jest wydzielanie testosteronu.
Spermatogeneza w cewkach nasiennych (tubuli seminiferi) jąder przebiega pod jednoczesną kontrolą hormonu folikulotropowego (FSH) i testosteronu. W cewkach nasiennych wytwarzane są hormony polipeptydowe
zwane inhibinami i aktywinami. Cząsteczki tych hormonów zbudowane są
z dwóch łańcuchów polipeptydowych połączonych ze sobą dwoma mostkami dwusiarczkowymi. Inhibiny są to heterodimery zbudowane z jednego łańcucha polipeptydowego alpha i jednego beta, które hamują
wydzielanie hormonu folikulotropowego (FSH). Natomiast aktywiny są
to homodimery, czyli cząsteczki zbudowane z dwóch takich samych
łańcuchów polipeptydowych, dwóch łańcuchów beta A lub betaB, które
pobudzają wydzielanie z przysadki hormonu folikulotropowego (FSH).
Rola testosteronu
Testosteron wydzielany jest do krwi osób obu płci, z tym że u mężczyzn
wydzielany jest w ilości kilkakrotnie większej niż u kobiet. Działanie
testosteronu na organizm kobiet jest również maskowane występowaniem
estrogenów, które mają w pewnym stopniu dzia łanie antagonistyczne
w stosunku do testosteronu.
Testosteron wywołuje:
• u mężczyzn — rozwój cech płciowych męskich obejmujący wykształ
cenie się zewnętrznych narządów płciowych, wzrost gruczołu kroko
wego (prostata), pęcherzyków nasiennych (vesiculae seminales), wy
kształcenie się budowy ciała i owłosienia typu męskiego;
• u płodów płci męskiej — różnicowanie się ośrodka rozrodczego
w pod wzgórzu w kierunku ośrodka typu męskiego. Ośrodek rozrodczy
typu męskiego jest niecykliczny, co wyraża się stałym pozio mem
wydzielania gonadotropin (FSH i LH);
• u obu płci — przyspieszenie syntezy białek, zatrzymanie wody i elektro
litów w organizmie, działając wraz z pozostałymi androgenami po
chodzącymi z kory nadnerczy.
260
Kontrola środowiska wewnętrznego
I Gruczoły przytarczyczne
Hormonem wydzielanym przez gruczoły przytarczyczne (glandulae
parathyroideae) jest hormon zwany parathormonem — PTH (parathyroid hormone). Jest to polipeptyd składający się z 84 aminokwasów
o masie cząsteczkowej 9,5 kDa i okresie połowicznego rozpadu we krwi
TV2 = 18 minut.
Parathormon zwiększa w osoczu krwi stężenie jonów wapniowych,
ponieważ pobudza:
• uwalnianie jonów Ca 2+ z tkanki kostnej do krwi;
• tworzenie się w nerkach witaminy D 3, która przyspiesza wchłanianie
jonów Ca 2+ do krwi w jelitach;
• resorpcję zwrotną jonów Ca 2+ w kanalikach nerkowych.
Zmniejszenie stężenia zjonizowanego wapnia w osoczu krwi pobudza
bezpośrednio komórki gruczołów przytarczycznych do wydzielania parathormonu, który, działając za pośrednictwem cyklicznego AMP na tkankę
kostną, uwalnia fosforan wapnia z tzw. magazynu wapnia wymiennego
w kościach (exchangeable bonę calcium reservoir). Jednocześnie parathormon zwiększa wchłanianie wapnia z przewodu pokarmowego. W tym
procesie działa synergicznie z witaminą D. Działanie parathormonu na
nerki sprowadza się do zwiększonego wydalania z moczem fosforanów
i do resorpcji zwrotnej jonów wapniowych.
Zwiększenie stężenia zjonizowanego wapnia w osoczu krwi bezpośrednio zwrotnie hamuje wydzielanie parathormonu przez gruczoły przytarczyczne.
Działanie antagonistyczne w stosunku do parathormonu ma kalcytonina wydzielana przez gruczoł tarczowy i zmniejszająca stężenie Ca 2 +
w osoczu krwi.
Czynność gruczołów dokrewnych
261
I Wyspy trzustkowe
Wyspy trzustkowe (Langerhansa) stanowią około 1% masy całej
trzustki. W wyspach trzustkowych {insulae pancreaticae) występują komórki A wydzielające glukagon, komórki B wydzielające insulinę i amylinę, komórki D wydzielające somatostatynę (SRIF) i komórki F —
trzustkowy polipeptyd (pancreatic polypeptide) o 36 aminokwasach. Glukagon jest polipeptydem składającym się z 29 aminokwasów o masie
cząsteczkowej 3,5 kDa. Insulina zaś jest hormonem białkowym składającym się z 2 łańcuchów polipeptydowych A i B, zawierających razem
51 aminokwasów o masie cząsteczkowej 5,7 kDa. Okres połowicznego
rozpadu insuliny we krwi (T 1/2) wynosi około 5 minut.
Insulina krążąca we krwi jest wychwytywana przez tkanki i maksymalny wynik jej działania występuje w czasie od 2 do 4 godzin. Komórki
wątrobowe są wystawione na działanie większego stężenia insuliny niż
komórki innych tkanek. Cała wydzielona przez komórki wysp trzustkowych insulina przepływa przez wątrobę we krwi żyły wrotnej. Tam
też przeważająca jej część jest rozkładana przez enzym wątrobowy
transhydrogenazę. Insulina zsyntetyzowana przez komórki B wysp trzustkowych jest w nich magazynowana w pęcherzykach cytoplazmatycznych.
Zwiększenie zawartości glukozy we krwi dopływającej do trzustki powoduje egzocytozę hormonu, polegającą na otwieraniu się błony komórkowej
w miejscu stykania się z pęcherzykami zawierającymi insulinę i wydzielaniu
ich zawartości na zewnątrz komórki.
Ryc. 135. Budowa schematyczna komórki
B wyspy trzustkowej (Langerhansa). (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
262
Kontrola środowiska wewnętrznego
Zwiększenie stężenia jonów Ca 2+ w cytoplazmie komórek B jest
czynnikiem wywołującym egzocytozę pęcherzyków zawierających insulinę. Insulina zmniejsza stężenie glukozy we krwi, co zwrotnie hamuje
wydzielanie komórek B i utrzymuje stałe stężenie glukozy we krwi.
Poza mechanizmem samoregulacji wydzielanie insuliny zwiększają:
impulsacja eferentna przewodzona przez nerw błędny, sekretyna, glukagon, enteroglukagon oraz adrenalina działająca poprzez beta-receptory
adrenergiczne i cykliczny AMP.
Pod wpływem zwiększającego się stężenia glukozy we krwi wydziela się
z komórek B wraz z insuliną drugi hormon — amylina. Ludzka amylina
(human amylin) jest polipeptydem o 37 aminokwasach, zbliżonym pod
względem budowy chemicznej do kalcytoniny i peptydu pochodnego
od genu kalcytoninowego (CGRP). Amylina działa antagonistycznie
w stosunku do insuliny, zwiększa uwalnianie glukozy z wątroby do krwi
i hamuje syntezę glikogenu w mięśniach poprzecznie prążkowanych
szkieletowych. W warunkach fizjologicznych wpływ insuliny na metabolizm węglowodanów znacznie przewyższa oddziaływanie antagonistyczne amyliny.
Glukagon, wytwarzany przez komórki A wysp trzustkowych, zwiększa
stężenie glukozy we krwi, aktywując fosforylazę w komórkach wątrobowych i przyspiesza glikogenolizę. Glukagon wydzielony przez wyspy
trzustkowe dostaje się do wątroby przez żyłę wrotną i tam prawie
całkowicie jest pochłaniany, tak że do krwi krążenia ogólnego przedostaje
się tylko w niewielkiej ilości. W czasie głodzenia wydzielanie glukagonu jest
znacznie zwiększone i prawidłowe stężenie glukozy we krwi jest zachowane. Działanie glukagonu jest antagonistyczne w stosunku do insuliny.
Insulina i glukagon są podstawowymi regulatorami przemiany węglowodanowej w organizmie, wpływają na aktywny transport przez błonę
komórkową i syntezę białek i tłuszczów w komórkach.
Ryc. 136. Regulacja zawartości glukozy we krwi.
Czynność gruczołów dokrewnych
263
Rola insuliny
Insulina działa na większość komórek w organizmie, ale szczególnie
silny jej wpływ zaznacza się na komórki wątrobowe, mięśniowe i tkankę
tłuszczową.
Pod wpływem insuliny wątroba zmniejsza uwalnianie i zwiększa
wchłanianie glukozy, zmniejsza wytwarzanie mocznika, zmniejsza stężenie cyklicznego AMP i zwiększa wychwytywanie fosforanów i potasu
z krwi. Działa za pośrednictwem kodu genetycznego kontrolującego
tworzenie w komórkach wątrobowych enzymów w ten sposób, że wytwarzanie enzymów związanych z syntezą glikogenu wątrobowego zwiększa się, enzymów glikogenolitycznych zaś maleje.
Komórki mięśni szkieletowych, mięśni gładkich i mięśnia sercowego
pod wpływem insuliny zwiększają aktywny transport przez błonę komórkową do wewnątrz: glukozy, aminokwasów, jonów potasowych i fosforanowych. Wewnątrz komórek mięśniowych zostaje przyspieszony metabolizm węglowodanów, synteza białek i synteza kwasów tłuszczowych.
Na tkankę tłuszczową insulina działa w ten sposób, że zwiększa syntezę kwasów tłuszczowych i triacylogliceroli oraz hamuje cyklazę adenylanową i tworzenie cyklicznego AMP. Zmniejszenie zawartości cyklicznego AMP hamuje uwalnianie z tkanki tłuszczowej wolnych kwasów
tłuszczowych.
I Szyszynka
Hormonem wydzielanym przez szyszynkę (epiphysis) jest melatonina.
Komórki gruczołowe szyszynki wychwytują z krwi 5-hydroksytryptaminę
i metabolizują ją do melatoniny, czyli N-acetylo-5-metoksytryptaminy,
oraz związków pochodnych.
Melatonina i jej pochodne metabolity wydzielane są do krwi i do płynu
mózgowo-rdzeniowego.
Wydzielanie szyszynki kontrolowane jest przez impulsy wysyłane przez
siatkówkę oka. Pod wpływem światła siatkówka wysyła impulsy nerwowe biegnące przez nerwy wzrokowe, jądro ponad skrzyżowaniem
wzrokowym podwzgórza (nucleus suprachiasmaticus hypothalami), pień
mózgowia i rogi boczne rdzenia kręgowego do zwoju szyjnego górnego
(ganglion cervicale superius). Synteza i wydzielanie melatoniny do krwi są
stale hamowane w ciągu dnia, wtedy kiedy światło pada na siatkówkę,
natomiast zwiększają się w czasie nocnych ciemności.
264
Kontrola środowiska wewnętrznego
Ryc. 137. Budowa schematyczna komórki szyszynki (pinealocytu). (Reprodukowano za
zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa
1997.)
Neurony zwojowe wspołczulne ze zwoju szyjnego górnego wysyłają
włókna zazwojowe do szyszynki. Uwalniana na zakończeniach tych
włókien noradrenalina, działając poprzez receptor beta-adrenergiczny
w błonie komórkowej pinealocytów i cykliczny AMP, pobudza te komórki do wydzielania melatoniny. Wydzielanie melatoniny jest jednym
z elementów tworzących dobowy zegar biologiczny.
Melatonina gromadzi się w tkance mózgowej, wpływając na ośrodki
związane ze snem i czuwaniem oraz z czynnością przysadki. Przede
wszystkim działa na ośrodki kontrolujące wydzielanie gonadotropin (FSH
i LH), opóźniając dojrzewanie płciowe (onset of puberty).
I Grasica
Z części korowej i rdzennej grasicy wyizolowano ponad dwadzieścia
związków o budowie peptydowej. Sześć z nich — to jest: tymopoetyna,
tymozyna alpha-1, alpha-7, beta-4, czynnik humoralny grasicy (thymic
humoral factor) i grasiczy czynnik surowicy (serum thymic factor), czyli
tymulina (thymulin) — ma uczestniczyć w tworzeniu się odporności
komórkowej w organizmie. Jako hormony grasicy mają brać udział
w procesie dojrzewania limfocytów T w samej grasicy stanowiącej
tkankę limfoidalną centralną, jak również po ich wyjściu z grasicy
Hormony tkankowe
265
w tkankach limfoidalnych obwodowych. Takie przypuszczenie oparte
jest na fakcie występowania hormonów grasicy w niej samej i we krwi
obwodowej (patrz str. 290).
Wrodzony brak grasicy prowadzi do zmniejszenia liczby limfocytów
we krwi obwodowej, zaniku układu limfoidalnego obwodowego, głównie
obszarów grasiczozależnych, oraz do braku odporności immunologicznej
typu komórkowego.
Ryc. 138. Udział podwzgórza, części gruczołowej przysadki i kory gruczołu nadnerczowego
w powstawaniu limfokin i tymozyny zwrotnie działającej na wydzielanie kortykoliberyny
(CRH) przez neurony peptydergiczne podwzgórza.
I Hormony tkankowe
W organizmie wytwarzane są liczne substancje, których działanie
ogranicza się do miejsca ich powstawania, lub które po dostaniu się do
krwi krążą w organizmie i działają na inne tkanki lub narządy.
W błonie śluzowej przewodu pokarmowego powstają hormony peptydowe żołądkowo-jelitowe kontrolujące wydzielanie soków trawiennych
i motorykę przewodu pokarmowego. Są to: gastryna, sekretyna, cholecystokinina, peptyd hamujący czynność żołądka (GIP), wazoaktywny
peptyd jelitowy (VIP), motylina, peptyd uwalniający gastrynę (GRP)
i somatostatyna (patrz Hormony żołądkowo-jelitowe, str. 450).
Nerki wydzielają do krwi reninę, która jest enzymem proteolitycznym,
i erytropoetynę działającą na erytropoezę w szpiku kostnym (patrz
str. 492).
266
Kontrola środowiska wewnętrznego
Komórki wątrobowe wydzielają polipeptyd — angiotensynogen —
i kininogeny, które są zamieniane w związki aktywne pod wpływem
enzymów krążących we krwi (patrz str. 492).
W tkankach wytwarzane są aminy biogenne jak histamina i serotonina,
które działają zarówno miejscowo, jak i ogólnie po przedostaniu się do
krwi.
Pochodne kwasu arachidonowego
Prostaglandyny (PG), prostacykliny (PGI), tromboksan (TxA) i leukotrieny (LT) są to eikozanoidy pochodne dwudziestowęglowych wielonienasyconych kwasów tłuszczowych, głównie kwasu arachidonowego.
Powstające PG w komórkach całego organizmu po przejściu do krwi
są szybko metabolizowane na związki fizjologicznie nieaktywne. Najszybciej PG są metabolizowane w płucach, wolniej w wątrobie, w nerkach i w łożysku. Nieaktywne metabolity usuwane są z organizmu
z moczem. Mała stabilność prostaglandyn w układzie krążenia kwalifikuje je do grupy hormonów tkankowych, o działaniu ograniczonym
do komórek, tkanek i narządów, w których są wytwarzane.
Pod wpływem prostaglandyn należących do grupy E, zwłaszcza PGE2,
w komórkach zwiększa się zawartość cyklicznego adenozynomonofosforanu (cAMP), zmniejsza zaś zawartość cyklicznego guanozynomonofosforanu (cGMP). Prostaglandyny należące do grupy F, zwłaszcza PGF 2,
mają działanie przeciwne. Pod ich wpływem w komórkach zwiększa się
zawartość cGMP, a zmniejsza cAMP.
Egzogenne PGEj wprowadzone dożylnie zmniejszają całkowity obwodowy opór naczyniowy (TPR), obniżają także ciśnienie krwi w zbiorniku tętniczym dużym i płucnym. PGF 2 wprowadzone tą samą drogą
działają kurcząco na błonę mięśniową naczyń krwionośnych, głównie
żylnych, i podwyższają ciśnienie w zbiorniku tętniczym płucnym.
W komórkach, zwłaszcza w komórkach tłuszczowych, PGE 2 są regulatorem procesów katabolicznych. Noradrenalina, działając za pośrednictwem beta-receptorów adrenergicznych, aktywuje cyklazę adenylanową
i przyspiesza tworzenie się cAMP. W wyniku tego zmagazynowane
w komórkach triacyloglicerole podlegają lipolizie. Uwalniane wolne
kwasy tłuszczowe, w tym również kwas arachidonowy, są substratem,
z którego biosyntetyzowane są prostaglandyny. Zwiększająca się w komórkach zawartość PGE 2 zwrotnie hamuje aktywność cyklazy adenylanowej i tworzenie się cAMP. Dzięki temu zostaje zwrotnie zahamowana
lipoliza w komórkach.
Hormony tkankowe
2Q7
W komórkach śródbłonka naczyń krwionośnych są syntetyzowane
prostacykliny PGI2 z prekursorów prostaglandyn. Nie ulegają one szybko
zmetabolizowaniu w płucach, tak jak PGE 2 i PGF2, są więc hormonem
krążącym we krwi. Pod wpływem PGI2 następuje rozszerzenie naczyń
krwionośnych w sercu, w nerkach, w przewodzie pokarmowym i w mięśniach poprzecznie prążkowanych oraz zwiększa się zawartość cAMP
w trombocytach.
Tabela 14
Najważniejsze eikozanoidy — miejsce ich powstawania i działania
Miejsce
Eikozanoid
Symbol
Działanie
powstawania
Prostaglandyna E
PGE2
komórki w całym
organizmie
rozkurcza błonę mięśniową
naczyń krwionośnych
Prostaglandyna F
PGF2
komórki w całym
organizmie
kurczy błonę mięśniową naczyń
krwionośnych
Tromboksan A2
TxA2
trombocyty
kurczy błonę mięśniową naczyń
krwionośnych
Prostacyklina I2
PGI2
hamuje agregację trombocytów
Leukotrien B4
Leukotrien C4
Leukotrien D4
Leukotrien E4
LTB4
LTC4
LTD4
LTE4
śródbłonek naczyń
krwionośnych
granulocyty
komórki tuczne
w płucach
chemotaksja dodatnia
kurczy mięśnie gładkie w ścianach oskrzeli i w ścianach naczyń
krwionośnych w płucach
Z powierzchni błony komórkowej granulocytów uwalniany jest leukotrien B 4 (LTB4) działający chemotaktycznie dodatnio i wyzwalający
odczyn zapalny w tkankach. Komórki tuczne w płucach uwalniają leukotrieny C 4, D4 i E4.
Leukotrieny są to aminolipidy, a więc wielonienasycone kwasy tłuszczowe związane poprzez atom siarki cysteiny z krótkimi peptydami:
leukotrien C 4 (LTC4) — z tripeptydem glutationowym, leukotrien D4
(LTD4) — z dipeptydem cysteinyloglicynowym, leukotrien E 4 (LTE4) — z
samą cysteiną, leukotrien F4 (LTF4) — z dipeptydem cysteinyloglutaminowym. Leukotrieny uwolnione w płucach wywołują skurcz mięśni
gładkich w ścianach oskrzeli i w ścianach naczyń krwionośnych krążenia
płucnego. Pod ich wpływem zwiększa się całkowity płucny opór naczyniowy.
268
Kontrola środowiska wewnętrznego
I Czynniki wzrostowe
Szczególne znaczenie w organizmie wśród hormonów tkankowych
mają czynniki wzrostowe (growth factors). Wytwarzane są przez komórki
różnych tkanek i oddziałują zarówno na komórki znajdujące się w sąsiedztwie (działanie parakrynne), jak i na komórki w odległych tkankach
(działanie endokrynne). Stale zwiększa się liczba nowo odkrytych czynników wzrostowych o specyficznym działaniu.
Obecnie czynniki wzrostowe można podzielić na cztery grupy:
•
•
•
•
czynniki wzrostowe tkankowe;
cytokiny i limfokiny;
czynniki hematopoetyczne;
czynniki angiogenetyczne.
Czynniki wzrostowe tkankowe
Do tkankowych czynników wzrostowych zalicza się: czynnik I i II
wzrostowy insulinopodobny, relaksynę, czynnik wzrostu nerwów, czynnik
wzrostu naskórka, czynnik wzrostu jajników i czynnik wzrostu fibroblastów.
Czynniki wzrostowe insulinopodobne I i II oraz relaksyna są to hormony
białkowe zbliżone pod względem budowy chemicznej do insuliny wytwarzanej przez komórki B wysp trzustkowych (patrz str. 261). Czynnik I
wzrostowy insulinopodobny — IGF-I (insulin-like growth factor-I), nazywany wcześniej somatomedyną C, wytwarzany jest głównie przez
komórki wątrobowe pod wpływem krążącego we krwi hormonu wzrostu
(hGH). Czynnik I wzrostowy insulinopodobny — pochodzący zarówno
z komórek wątrobowych, jak i wytwarzany miejscowo w tkance chrzestnej — powoduje rozrost tkanki chrzestnej, szczególnie chrząstek przynasadowych kości długich i ich wydłużanie się.
Czynnik wzrostowy insulinopodobny II (IGF-II) powstaje poza wątrobą
w komórkach różnych tkanek. Ma podobne do czynnika I działanie
na tkankę kostną.
Relaksyna wytwarzana jest u kobiet w ciałku żółtym miesiączkowym
i ciążowym oraz w łożysku w czasie ciąży. Pod jej wpływem następuje
rozluźnienie połączeń kości miednicy, szczególnie spojenia łonowego
i rozluźnienie szyjki macicy.
Hormony tkankowe
269
Tabela 15
Czynniki wzrostowe
Skrót
Nazwa
lub
symbol
IGF-I
IGF-II
Liczba
aminokwasów
czynnik wzrostowy
insulinopodobny I
czynnik wzrostowy
insulinopodobny II
130
180
118
Narządy i tkanki
wydzielające
wątroba i inne
tkanki
wzrost kości i
chrząstek
wątroba i inne
tkanki
wzrost narządów u
płodów
wzmaga wzrost
neuronów i ich
wypustek, hamuje
apoptozę neuronów
tkanki unerwione
przez nerwy czuciowe lub autonomiczne
NGF
czynnik wzrostu
nerwów
EGF
czynnik wzrostu
naskórka
53
czynnik wzrostu
fibroblastów
155
relaksyna
—
wątroba i inne
tkanki
czynniki wzrostu
transformujący
alpha
czynnik wzrostu
transformujący
beta
112
większość tkanek
w organizmie
FGF-1 —
FGF-9
—
TGF-a
TGF-p"
Działanie
większość tkanek
w organizmie
wzmaga podziały
mitotyczne, hamuje
podziały komórek
nowotworowych
wzmagają podziały
mitotyczne
rozluźnia więzadła
przed porodem
wzrost i różnicowanie się komórek
wielu tkanek w czasie embriogenezy
Pominięte zostały czynniki wzrostowe hematopoetyczne.
Cytokiny i limfokiny
Cytokiny i limfokiny pobudzające wzrost są to przekaźniki chemiczne
wytwarzane przez limfocyty, makrofagi, komórki śródbłonka naczyniowego, komórki szpiku kostnego czerwonego, komórki glejowe, fibroblasty
i komórki innych tkanek. Pod wpływem cytokin i limfokin dochodzi
do licznych reakcji komórkowych, w tym również do wzrostu komórek
biorących udział w odpowiedzi immunologicznej organizmu na działanie
270
Kontrola środowiska wewnętrznego
antygenu. Do tej grupy czynników wzrostowych zalicza się interleukiny
(interleukins): 1-alpha (IL-1 a), 1-beta (IL-lp), 2 (IL-2), 4 (IL-4), 5 (IL-5),
6 (IL-6) i 7 (IL-7) oraz cytokiny: czynnik martwicy nowotworów (TNF),
interferony alpha (IFN-a), beta (IFN-(3) i gamma (IFN-y) (patrz str. 268).
Czynniki wzrostowe hematopoetyczne
U ludzi dorosłych komórki układu hematopoetycznego intensywnie
wzrastają, dzielą się i różnicują. Procesy te regulowane są przez czynniki
wzrostowe hematopoetyczne działające bezpośrednio na (1) komórki
szpiku pluripotencjalne hematopoetyczne pnia i macierzyste nieukierunkowane oraz na (2) komórki prekursorowe linii erytrocytów, granulocytów i monocytów.
(1) Interleukina 3 (IL-3) pobudza wzrost w szpiku kostnym czerwo
nym komórek pluripotencjalnych hematopoetycznych pnia i komórek
ukierunkowanych linii erytrocytów (BFU-E). Wyzwala również różnico
wanie się limfocytów B i proliferację limfocytów T oraz ekspresję genów
dla czynnika wzrostowego makrofagów (CSF-M).
Czynnik wzrostowy granulocytów i makrofagów (CSF-GM) wywołuje
proliferację komórek pluripotencjalnych hematopoetycznych pnia, komórek ukierunkowanych linii erytrocytów (BFU-E), neutrofilów (CFU-G),
eozynofilów (CFU-EoS) i makrofagów (CFU-M). Czynnik ten wyzwala
również ekspresję genu dla interleukiny 1 (IL-1) w neutrofilach i w innych
leukocytach (patrz tab. 21, str. 309).
(2) Do czynników wzrostowych hematopoetycznych Unii erytrocytów,
neutrofilów, monocytów i limfocytów zalicza się pięć czynników wzro
stowych. Erytropoetyna (EPO) pobudza wzrost klonalny komórek
linii erytrocytów (BFU-E, CFU-E). Czynnik wzrostowy granulocytów
(CSF-G) pobudza proliferację komórek linii neutrofilów, jak również
aktywuje ich właściwości fagocytarne. Wzrost monocytów i makrofagów
zachodzi pod wpływem czynnika wzrostowego makrofagów (CSF-M),
który również aktywuje właściwości fagocytarne i wydzielnicze makro
fagów. Interleukina 7 (IL-7) pobudza wzrost prolimfocytów B i T, inter
leukina 12 (IL-12) zaś wyzwala różnicowanie się limfocytów T h pomagają
cych (patrz tab. 21).
Hormony tkankowe
TT\
Czynniki wzrostowe angiogenetyczne
Czynniki wzrostowe pobudzające angiogenezę wytwarzane są przez
fibroblasty, komórki śródbłonka naczyniowego i inne komórki, jak
również uwalniane są z trombocytów. Czynnik naczyniowy wzrostu
śródbłonka — VEGF (vascular endothelial growth factor) — zbudowany
jest z dwóch homogennych podjednostek, każda o masie cząsteczkowej
23 kDa. Początkowo został wykryty w komórkach przysadki, ale występuje również w innych komórkach. Poza silną aktywnością angiogenetyczną w stosunku do komórek śródbłonka naczyniowego, zwiększa
również przepuszczalność ścian naczyń krwionośnych.
Czynniki wzrostowe fibroblastów — FGFs (fibroblast growth fac-tors)
— stanowią grupę przekaźników chemicznych pobudzających do
wzrostu komórki pochodzące z mezodermy. Czynnik wzrostowy transformujący alpha — TGF-a (transforming growth factor — alpha) — jest
polipeptydem o masie cząsteczkowej 55 kDa, wytwarzanym przez transformujące fibroblasty, jak również przez makrofagi. Działa poprzez
receptor dla czynnika wzrostu naskórka (EGF). Czynnik wzrostowy
transformujący beta (TGF-p) ma masę cząsteczkową 25 kDa i składa się
z dwóch homologicznych dimerów. Działa podobnie do TGF-a.
Czynnik wzrostu śródbłonka pochodzenia płytkowego — PDECGF
(platelet-derived endothelial celi growth factor) — jest to pojedynczy
łańcuch polipeptydowy o masie cząsteczkowej 45 kDa. Występuje w trombocytach oraz w komórkach wielu tkanek. Działa wyłącznie na wzrost
komórek śródbłonka naczyniowego.
Czynnik martwicy nowotworów alpha — TNF-a (tumor necrosis factor-alpha) — wytwarzany jest przez aktywowane makrofagi. Prawdopodobnie pobudza angiogenezę i tworzenie się naczyń krwionośnych włosowatych w tkankach objętych procesem zapalnym lub nowotworowym.
Angiogenina (angiogenin) jest polipeptydem o silnym działaniu angiogenetycznym, o masie cząsteczkowej 14,4 kDa, wytwarzanym przez fibroblasty i komórki wątrobowe.
Interleukina 8 (IL-8) wytwarzana przez makrofagi jest również czynnikiem angiogenetycznym. Przypuszcza się, że takie właściwości mają
również białka układu krzepnięcia krwi — trombina (czynnik II) i fibryna
(czynnik I).
272
Kontrola środowiska wewnętrznego
I Termoregulacja
Ssaki, a więc i człowiek, zostały wyposażone w rozwoju filogenetycznym w fizjologiczny mechanizm utrzymujący stałą temperaturę środowiska wewnętrznego organizmu. Dzięki temu mechanizmowi temperatura krwi przepływającej przez serce waha się jedynie nieznacznie
i wynosi około 37°C. Z punktu widzenia praktyki lekarskiej można
temperaturę ciała uważać za stałą, mając na uwadze cykliczne dzienne
jej zmiany w granicach 0,5—0,7°C. Temperatura ciała jest najniższa
w czasie snu we wczesnych godzinach rannych, a najwyższa we wczesnych
godzinach wieczornych.
U kobiet temperatura ciała zmienia się w zależności od okresu cyklu
miesiączkowego — podwyższa się w dniu owulacji i utrzymuje na tym
poziomie w fazie lutealnej cyklu miesiączkowego.
Temperatura środowiska wewnętrznego jest w znacznym stopniu
niezależna zarówno od szybkości utraty ciepła, jak i wielkości wytwarzania ciepła w organizmie.
Odprowadzanie ciepła z
organizmu
Organizm traci ciepło (body heat loss) w stosunku do otoczenia o niższej temperaturze przez:
• skórę (promieniowanie, przewodnictwo i pocenie się);
• układ oddechowy (z wydychanym powietrzem);
• przewód pokarmowy i układ moczowy (z kałem i moczem).
Podwyższenie temperatury krwi dopływającej do podwzgórza, do
ośrodka termoregulacji, powoduje pobudzenie termodetektorów i przyspiesza utratę ciepła, co zachodzi w wyniku:
• rozszerzenia naczyń skórnych i wydzielania potu przez gruczoły po
towe;
• przyspieszenia pracy serca i pogłębienia oddechów. Przez płuca prze
pływa więcej krwi i z powietrzem wydychanym odprowadzane jest
więcej ciepła;
• pobudzenia ośrodka hamującego drżenie mięśniowe w śródmózgowiu.
Pod wpływem tego ośrodka nie występuje drżenie mięśniowe, czyli
skurcze komórek mięśniowych.
Termoregulacja
273
Wytwarzanie ciepła w organizmie
Wytwarzanie ciepła w organizmie (body heat production) zależy od:
• podstawowej przemiany materii (BMR — basal metabolic ratę) zwią
zanej ze spoczynkową czynnością wszystkich komórek i narządów
niezbędną do utrzymania organizmu przy życiu;
• pracy mięśni szkieletowych w czasie poruszania się;
• czynności przewodu pokarmowego związanej z trawieniem i wchła
nianiem pokarmów.
Nadmiar ciepła wytworzonego w organizmie jest odprowadzany do
otoczenia, jak również niedobór ciepła w organizmie uruchamia fizjologiczne mechanizmy zwiększające jego wytwarzanie.
Ośrodek termoregulacji
Temperaturę ciała utrzymuje na stałym poziomie ośrodek termoregulacji znajdujący się w części przedniej podwzgórza. Ośrodek ten działa
na zasadzie termostatu, kontrolując inne ośrodki.
Obniżenie temperatury krwi przepływającej przez podwzgórze hamuje
termodetektory, co powoduje zwiększone wytwarzanie ciepła lub zmniejszoną jego utratę za pośrednictwem:
• ośrodka wyzwalającego drżenie mięśniowe — przestaje on być hamo
wany przez ośrodek termoregulacji. Pojawiają się dreszcze, czyli skur
cze komórek mięśniowych i wytwarza się ciepło w mięśniach szkie
letowych — termogeneza drżeniowa (shivering thermogenesis);
• ośrodków kontrolujących układ współczulny — pobudzenie układu
współczulnego powoduje wydzielanie się z zakończeń neuronów zwojo
wych noradrenaliny. Pod wpływem noradrenaliny przyspiesza się
metabolizm komórek mięśni szkieletowych i komórek tkanki tłusz
czowej;
• ośrodków kontrolujących wydzielanie gruczołów dokrewnych —
z rdzenia nadnerczy wydziela się adrenalina przyspieszająca metabolizm
glukozy w wątrobie i w mięśniach szkieletowych oraz metabolizm
tkanki tłuszczowej;
• hormonów gruczołu tarczowego (trijodotyroniny i tyroksyny) — dłu
żej trwające obniżenie średniej temperatury otoczenia powoduje stałe
zwiększenie metabolizmu wewnątrzkomórkowego w organizmie. Biorą
w tym udział: podwzgórzowy hormon uwalniający hormon tyreotro-
274
Kontrola środowiska wewnętrznego
powy (TRH), hormon tyreotropowy z części gruczołowej przysadki
(TSH) i hormony gruczołu tarczowego — trijodotyronina i tyroksy-na
— działające bezpośrednio na komórki organizmu; • ośrodka
naczyniowego — naczynia skórne zwężają się i zmniejsza się utrata
ciepła przez skórę.
Przysadka
: -------część
----------------------------------------------TSH
gruczołowa
Tarczyca ------------------------------------------------------------------- "■ T3 + T
\
W
y
i
w
a
r
z
a
n
i
e
c
i
e
p
ł
a
w
tkankach
Ryc. 139. Udział hormonu gruczołu tarczowego (T 3 i T4) w termoregulacji w warunkach
długotrwałej niskiej średniej temperatury otoczenia.
Stosunek jonów Ca 2+ do jonów Na + w podwzgórzu warunkuje
wrażliwość termodetektorów na zmiany temperatury krwi dopływającej
do mózgowia. Przesunięcie tego stosunku na korzyść jonów Ca 2+ powoduje przestawienie się termostatu biologicznego na niższą temperaturę
i temperatura ciała obniża się o ułamek stopnia. Zwiększenie koncentracji jonów Na+ w podwzgórzu wywołuje reakcję przeciwną —
przestawienie termostatu biologicznego na wyższą temperaturę i temperatura ciała podwyższa się.
Gorączka
Termodetektory w podwzgórzu pod wpływem wytwarzanych miejscowo prostaglandyn stają się mniej wrażliwe na podwyższającą się
temperaturę krwi. Termostat biologiczny przestawia się na wyższą tem-
Termoregulacja
27'5
peraturę krwi, co powoduje podwyższenie się temperatury ciała, czyli
wystąpienie gorączki.
Toksyny bakteryjne i inne czynniki powodują wytwarzanie przez
prawie wszystkie komórki w organizmie interleukiny 1 (IL-1), wcześniej
nazywanej endogennym pirogenem (endogenous pyrogen), czynnikiem
aktywującym leukocyty (leukocyte-activating factor) i jeszcze inaczej
nazywanej. Interleukina 1 jest polipeptydem o masie cząsteczkowej 17 kDa
występującym w dwóch postaciach, jako interleukina 1 alpha (IL -1 a)
i interleukina 1 beta (IL-1J3). Pod wpływem toksyn bakteryjnych komórki
w organizmie, szczególnie makrofagi i leukocyty, wytwarzają interleukinę 1, która krążąc we krwi, przenika do podwzgórza w miejscach
pozbawionych bariery krew-mózg, a więc w narządzie naczyniowym
blaszki krańcowej ponad skrzyżowaniem wzrokowym i w narządzie
podsklepieniowym. Pod wpływem interleukiny 1 tworzą się w podwzgorzu
prostaglandyny — termostat biologiczny przestawia się na wyższą temperaturę i występuje gorączka.
Krążenie
Krążenie płynów w organizmie wyrównuje różnice występujące w środowisku wewnętrznym oraz stanowi drogę przenoszenia informacji zakodowanej w postaci substancji chemicznych.
Krążenie obejmuje:
• płyny ustrojowe — krew (blood), chłonkę (lymph) i płyn tkankowy
(interstitial fluid);
• łożysko krwionośne (vascular bed), które wypełnione jest przepływa
jącą krwią dzięki:
o sercu zapewniającemu różnicę ciśnień pomiędzy zbiornikami krwi
(blood reservoirs), o układowi naczyń krwionośnych, czyli
zbiornikom krwi i połączeniom
pomiędzy zbiornikami w postaci sieci naczyń włosowatych.
I Krew
Krew wypełniająca łożysko krwionośne stanowi tkankę płynną, odgraniczoną od innych tkanek organizmu co najmniej jedną warstwą
komórek, którą tworzą komórki śródbłonka naczyniowego.
Całkowita objętość krwi (TBV — total blood volume) wypełniająca
łożysko krwionośne stanowi od 1/20 do 1/13 masy ciała.
Krew zawiera:
• elementy upostaciowane, czyli erytrocyty (krwinki czerwone), leukocyty
(krwinki białe) i trombocyty (płytki krwi);
• nieupostaciowane składniki osocza krwi.
Czynność tkanek krwiotwórczych
277
Elementy morfotyczne stanowią mniej niż 50% objętości krwi. Stosunek objętości erytrocytów do objętości pełnej krwi nosi nazwę wskaźnika
hematokrytu — Het (hematocrit).
W składzie krwi u mężczyzn i u kobiet występują różnice. Z tego
względu wartości odnoszące się do krwi muszą uwzględniać różnice płci.
I Rola krwi w organizmie
Zasadniczą rolą krwi w organizmie jest utrzymanie stałego środowiska
wewnętrznego. Krew spełnia swe funkcje dzięki temu, że krążąc w organizmie:
• transportuje tlen z płuc do tkanek;
• transportuje dwutlenek węgla z tkanek do płuc;
• transportuje do wszystkich tkanek produkty energetyczne i budulcowe
wchłonięte z przewodu pokarmowego;
• transportuje wchłonięte z tkanek produkty przemiany materii do nerek,
skąd zostają one wydalane z organizmu wraz z moczem;
• transportuje hormony syntetyzowane w organizmie i witaminy wchło
nięte w przewodzie pokarmowym;
• magazynuje hormony gruczołu tarczowego i hormony steroidowe po
ich związaniu z białkami osocza;
• wyrównuje ciśnienie osmotyczne we wszystkich tkankach;
• wyrównuje stężenie jonów wodorowych (pH) we wszystkich tkankach;
• wyrównuje różnice temperatur występujące pomiędzy poszczególnymi
narządami i tkankami;
• tworzy zaporę przed inwazją drobnoustrojów, które po dostaniu się do
środowiska wewnętrznego są stale pożerane przez leukocyty;
• eliminuje za pomocą przeciwciał i układu dopełniacza substancje obce,
szczególnie o charakterze białkowym, np. produkty przemiany drobno
ustrojów — toksyny.
I Czynność tkanek krwiotwórczych
Tkanki krwiotwórcze, wytwarzające elementy morfotyczne krwi, dzielą
się na pierwotne — czyli centralne tkanki hematopoetyczne (central
hematopoetic tissues), do których zalicza się szpik kostny czerwony i
grasicę, oraz na wtórne — czyli obwodowe tkanki hematopoetyczne
278
Krążenie
(peripheral hematopoetic tissues) obejmujące węzły chłonne, grudki chłonne w błonach śluzowych i śledzionę.
Komórki hematopoetyczne pluripotencjalne pnia, jak i komórki hematopoetyczne ukierunkowane, mogą przemieszczać się pomiędzy tkankami
hematopoetycznymi wraz z krwią krążącą w układzie krążenia.
I Szpik kostny
Szpik kostny czerwony {medulla ossium rubra) jest zasadniczym narządem krwiotwórczym w życiu pozapłodowym. Szpik kostny stanowi około
5% masy ciała, w tym około połowa, czyli 2,5% masy ciała, przypada na
szpik czerwony, w którym powstają elementy morfotyczne krwi. Wypełnia
istotę gąbczastą {substantia spongiosa) kości płaskich, mostek (sternum),
żebra (costae), kości biodrowe (ossa ilia), trzon kręgów (corpus vertebrae)
oraz jamy szpikowe {cava medullaria) w sąsiedztwie nasad (epiphyses)
kości długich. W okresie wzmożonej czynności szpik kostny czerwony
zwiększa swą masę, zajmując miejsce szpiku kostnego żółtego.
W szpiku kostnym czerwonym, poza komórkami hematopoetycznymi,
o różnym stopniu zróżnicowania, występują w niewielkiej liczbie adipocyty, komórki zrębu, komórki tworzące ściany zatok szpikowych, czyli
komórki śródbłonka i przydanki, oraz komórki bariery. Komórki stano-
Komórki hematopetyczne
Fibroblast zrębu
Ryc. 140. Budowa schematyczna szpiku kostnego czerwonego. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa
1997.)
Czynność tkanek krwiotwórczych
279
wiące zrąb szpiku kostnego (stroma medullae) są to komórki siateczki
(cellulae reticulares) tworzące sieć, na której zawieszone jest utkanie tkanki
szpikowej. Poza funkcją tkanki podporowej komórki siateczki spełniają
istotną rolę w hematopoezie. Mają właściwości żerne, pochłaniając jądra
komórkowe wydalane przez dojrzewające erytroblasty. Wydzielają czynniki wzrostowe, leukotrieny, cytokiny, prostaglandyny i inne przekaźniki
chemiczne. Zwiększają również stężenie czynników wzrostowych w szpiku
w wyniku ich adsorpcji na błonie komórkowej.
Komórki bariery (barrier cells) mają zdolność do kurczenia się, łącząc
się zaś ze sobą, otaczają kolonie komórek hematopoetycznych, chroniąc
KOMÓRKA
CFU-E
KOMÓRKA
C FU -G
KOMÓRKA
CFU-M
KOMÓRKA
CFU-Eos
KOMÓRKA KOMÓRKA
CFU-Baso CFU-Mk
LIMFOCYT
Ryc. 141. Hematopoeza w szpiku kostnym czerwonym. Z komórki pluripotencjalnej pnia
tworzą się komórki macierzyste nieukierunkowane pnia (komórki CFU-GEMM) i komórki
macierzyste limfoidalne pnia — komórki prekursorowe limfocytów szpikowych. Komórki
CFU-GEMM różnicują się i przekształcają w komórki ukierunkowane: linii erytrocytów
(komórki BFU-E i komórki CFU-E), linii granulocytów i makrofagów (komórki CFU-GM),
linii eozynofilów (komórki CFU-Eos), linii bazofilów (komórki CFU-Baso), linii megakariocytów (komórki BFU-Mk i komórki CFU-Mk). Komórki ukierunkowane CFU-GM
przekształcają się w komórki ukierunkowane linii neutrofilów (komórki CFU-G) i linii
makrofagów (CFU-M). Czynniki hematopoetyczne: CSF-G, IL-1, IL-6, IL-7, IL-10, IL-11,
IL-12.
280
Krążenie
Tabela 16
Czynniki wzrostowe hematopoetyczne
Skrót lub
symbol
Nazwa
Komórki wydzielające
Główne działanie
CSF-GM
czynnik wzrostowy
komórek prekursorowych granulocytów i
monocytów
komórki zrębu szpiku
kostnego czerwonego
limfocyty T komórki
tuczne
hematopoeza w szpiku
kostnym czerwonym komórek niezróżnicowanych
CSF-G
czynnik wzrostowy
komórek prekursorowych neutrofilów
komórki zrębu szpiku
kostnego czerwonego
makrofagi, monocyty
hematopoeza w szpiku
kostnym czerwonym komórek niezróżnicowanych
i prekursorów neutrofilów
CSF-M
czynnik wzrostowy
monocytów
komórki zrębu szpiku
kostnego czerwonego
makrofagi, monocyty
powstawanie monocytów i
makrofagów
erytropoetyna
nerki
wątroba
różnicowanie i dojrzewanie komórek prekursorowych erytrocytów
EPO
Interleukiny o działaniu wzrostowym hematopoetycznym przedstawione są w tabeli 21.
je przed działaniem czynników niepożądanych, jak również wpływają na
przepływ krwi przez różne miejsca w jamach szpikowych.
Zgodnie z teorią unitarystyczną wszystkie elementy morfotyczne krwi
pochodzą od komórki szpiku pluripotencjalnej hematopoetycznej pnia
(pluripotent hematopoietic stem celi). Pod wpływem czynnika wzrostowego granulocytów (CSF-G) sześciu interleukin (IL-1, IL-6, IL-7, IL-10,
IL-11, IL-12) i innych czynników z komórki pluripotencjalnej powstają
komórki macierzyste nieukierunkowane (CFU-GEMM) i komórki macierzyste limfoidalne (lymphoid progenitor).
Komórki macierzyste nieukierunkowane (komórki CFU-GEMM) przekształcają się w:
• komórki ukierunkowane linii erytrocytów, czyli komórki tworzące
kolonie rozsadzające erytroidalne — BFU-E (burst-forming unit erythroid), i następne komórki macierzyste linii erytrocytów — CFU-E
(colony-forming unit erythroid);
• komórki ukierunkowane granulocytów i makrofagów — CFU-GM (co
lony-forming unit — granulocyte/macrophage);
• komórki macierzyste linii eozynofilów — CFU-Eos (colony-forming
unit — eosinophils);
Czynność tkanek krwiotwórczych
281
• komórki macierzyste linii bazofilów — CFU-Baso (colony-forming
unit — basophils);
• komórki ukierunkowane linii megakariocytów, czyli komórki tworzące
kolonie rozsadzające megakariocytarne — BFU-Mk (burst-forming
unit — megakaryocytes) i następne komórki macierzyste megakariocy
tów ukierunkowane — CFU-Mk (colony-forming unit — megakaryo
cytes).
Komórki ukierunkowane granulocytów i makrofagów (komórki CFUGM) przekształcają się dalej w komórki macierzyste linii neutrofilów
(komórki CFU-G) i komórki macierzyste linii makrofagów (komórki
CFU-M).
Tabela 17
Elementy morfotyczne w szpiku kostnym czerwonym i we krwi obwodowej
Układ
Szpik kostny
Krew obwodowa
Zakres prawiOkres
Średnia
Średnia
dłowych
połowicznego
(%)
(%)
wartości
rozpadu
(%)
Granulocytamy
56,2
50,4—70,5
0,088
7 godzin
Erytrocytarny
25,6
18,4—33,8
99,857
Limfocytarny
16,2
11,1—23,2
0,047
28 dni
od kilku dni do
kilkunastu
tygodni
Monocytarny
Megakariocytarny
0,3
0,1
0—0,8
0—0,4
0,005
_*
kilka dni
trombocyty
4—5 dni
Komórki
plazmatyczne
Komórki siateczki
1,3
0,4—3,9
0,3
0—0,9
—
—
-
—
* Z jednego megakariocytu powstają liczne trombocyty.
We wzroście i różnicowaniu się komórek progenitorowych nieukierunkowanych w komórki ukierunkowane biorą udział liczne czynniki wzrostowe, w tym pięć interleukin (patrz ryc. 141, str. 279).
Progenitorowe komórki limfoidalne przekształcają się w szpiku kostnym
w limfocyty pre-B i limfocyty pre-T oraz limfocyty naturalni niszczyciele
— limfocyty NK (natural killer cells).
282
Krążenie
Wszystkie komórki hematopoetyczne szpiku kostnego należą do jednej z trzech grup:
• puli komórek pnia (stem-cells pool);
• puli komórek dzielących się (proliferative pool);
• puli komórek dojrzewających i rezerwy szpikowej (maturation and
reserve pool).
Erytrocytopoeza
W szpiku kostnym czerwonym z komórek macierzystych linii erytrocytów, komórek CFU-E, powstają kolejno: proerytroblasty, erytroblasty
zasadochłonne I i II oraz erytroblasty polichromatofilne. Wszystkie te
komórki należą do puli komórek dzielących się. Na etapie erytroblastów
zasadochłonnych zachodzi w tych komórkach ekspresja genów związanych z syntezą łańcuchów polipeptydowych globiny. Cytoplazma erytroblastów zasadochłonnych i polichromatofilnych zaczyna wypełniać się hemoglobiną. W miarę wypełniania się cytoplazmy hemoglobiną w jądrach
komórkowych zmniejsza się synteza matrycowego kwasu rybonukleinowego (mRNA). Erytroblasty ortochromatyczne przeciskają się pomiędzy
komórkami tworzącymi ścianę szpikowych zatok żylnych, pozostawiając
piknotyczne jądra komórkowe w miąższu szpiku, gdzie są fagocytowane
przez komórki siateczki. Tak powstające retikulocyty znajdują się w szpikowych zatokach żylnych, tworząc pulę rezerwy szpikowej.
Cykl rozwojowy erytrocytów w szpiku kostnym, czyli łączny czas
przypadający na dzielenie, różnicowanie i dojrzewanie, począwszy od
komórek pnia aż do erytrocytu, trwa około 5 dni.
We krwi obwodowej retikulocyty stanowią około 0,5% puli eryt rocytów krążących we krwi. Po utracie znacznych ilości krwi zwiększa się
procent retikulocytów we krwi w związku z ich przyspieszonym przechodzeniem z puli rezerwy szpikowej do puli komórek krążących.
Zasadniczym czynnikiem wzrostowym pobudzającym erytrocytopoezę
jest erytropoetyna — EPO (erythropoietin). Jest to białko o masie
cząsteczkowej 18 kDa (34—39 kDa kiedy jest całkowicie glikozylowane)
wytwarzane u ludzi dorosłych w 85% w nerkach i w 15% w wątrobie.
Zmniejszenie prężności tlenu w nerkach jest zasadniczym czynnikiem
zwiększającym wydzielanie erytropoetyny do krwi (patrz Czynność wewnątrzwydzielnicza nerek, str. 492).
Jony żelaza są niezbędne w biosyntezie hemoglobiny. Erytroblasty
pozyskują jony żelaza za pośrednictwem białka osocza — transferryny
Czynność tkanek krwiotwórczych
283
ERYTROCYT
^yC- 1^* Erytrocytopoeza w szpiku kostnym
czerwonym. Z komórki ukierunkowanej linii erytrocytów CFU-E powstają kolejne proerytroblasty i erytroblasty. Erytroblasty ortochromatyczne tracą jądro komórkowe i przechodzą z miąższu szpiku do zatok szpikowych,
przekształcając się w retikulocyty. Czynniki hematopoetyczne: CSF-GM, IL-3, IL-9, IL-11,
EPO.
^Mft
284
Krążenie
(transferrin). U dorosłych mężczyzn stężenie żelaza w osoczu wynosi
23 umol/L (130 ug/dL), natomiast u dorosłych miesiączkujących ko - I
biet — 19 umol/L (110 ug/dL). Utrzymanie takiego stężenia wymaga
stałego wprowadzania żelaza wraz z pokarmami, u mężczyzn około 10 mg
na dobę i u kobiet około 18 mg na dobę. Tylko część jonów żelaza I
wchłaniana jest w przewodzie pokarmowym. Wchłanianie jonów żelaza
zależy od wielu czynników, w jakich pokarmach występują i czy są to jo- I
ny Fe2+, czy też jony Fe3+.
Erytroblasty wychwytują żelazo z osocza dzięki receptorom transferrynowym. Najwięcej receptorów transferrynowych mają na błonie komórkowej erytroblasty zasadochłonne. W tych komórkach zachodzi
najintensywniejsza biosynteza hemoglobiny. Niedobór jonów żelaza
w osoczu krwi — i tym samym zmniejszony ich wychwyt przez erytroblasty — hamuje biosyntezę hemoglobiny na etapie translacji mRNA dla
hemoglobiny. Zapobiega to gromadzeniu się metabolitów hemu w erytroblastach i następnie w erytrocytach.
Prawidłowa erytrocytopoeza jest również uzależniona od dwóch witamin, a mianowicie od witaminy B 12 i od kwasu foliowego.
Zapotrzebowanie dobowe na witaminę B12 (cyanocobalamin) wynosi
od 1 do 3 ng, w wątrobie zaś jest zmagazynowane 3—4 mg witaminy B12.
Ilość zmagazynowana pokrywa zapotrzebowanie na 3 lata.
Witamina B12 zawarta w pokarmach łączy się w żołądku z czynnikiem
wewnętrznym (intrinsic factor) wydzielanym przez błonę śluzową żołądka
i w tej postaci zostaje w jelicie krętym wchłonięta do krwi. W przypadkach
zmniejszonego wydzielania przez błonę śluzową żołądka czynnika wewnętrznego witamina B12 nie jest wchłaniana w jelicie.
Kwas foliowy (folie acid) spożywany z pokarmami jest wchłaniany
w dwunastnicy i jelicie cienkim. Zapotrzebowanie dobowe na kwas foliowy wynosi 50 ug, a ilość zmagazynowana w wątrobie pokrywa
zapotrzebowanie na około 4 miesiące.
W regulacji erytrocytopoezy i — pośrednio — liczby erytrocytów we
krwi obwodowej uczestniczą:
• erytropoetyna;
• czynniki wzrostowe takie, jak interleukiny (IL-3, IL-9, IL-11) i czynnik
wzrostowy neutrofilów i makrofagów (CSF-GM);
• hormony gruczołu tarczowego (T 3 i T4) zwiększające erytrocytopoezę;
• hormony płciowe, androgeny pobudzają, metabolity estrogenów zaś
hamują erytrocytopoezę.
Czynność tkanek krwiotwórczych
285
Granulocytopoeza
Z trzech różnych komórek macierzystych szpiku kostnego czerwonego
wywodzą się granulocyty, czyli: (1) leukocyty obojętnochłonne — neutrofile, (2) leukocyty eozynochłonne — eozynofile i (3) leukocyty zasadochłonne — bazofile.
Neutrofile pochodzą od komórki macierzystej linii neutrofilów, czyli
komórki CFU-G. Z tych komórek powstają kolejno: mieloblasty, promielocyty i mielocyty tworzące pulę komórek dzielących się (proliferative
pool).
Proliferacja i dojrzewanie komórek szpikowych linii neutrofilów zachodzi pod wpływem czynników wzrostowych, takich jak: czynnik wzrostowy granulocytów — CSF-G (colony-stimulating factor-granulocyte),
czynnik wzrostowy pierwszy — CSF-1 (colony-stimulating factor 1), i
czynnik wzrostowy granulocytów i makrofagów — CSF-GM (colony-stimulating factor granulocyte/macrophage).
Dalszy etap rozwoju komórek szpiku linii neutrofilów, to ich dojrzewanie bez mitotycznego podziału. Do puli komórek dojrzewających
i rezerwy szpikowej (maturation and reserve pool) należą metamielocyty i neutrofile o jądrach komórkowych pałeczkowatych i segmentowanych.
W szpiku kostnym czerwonym komórki linii neutrofilów są najliczniejsze. Łączny czas potrzebny na przejście od pluripotencjalnej komórki pnia
przez wszystkie etapy podziałów, różnicowania i dojrzewania do postaci
neutrofilów segmentowanych wynosi około 6—7 dni.
Komórki szpiku linii eozynofilów pochodzą od komórki macierzystej
linii eozynofilów — CFU-Eos (colony forming unit eosinophils) i przechodzą kolejno etapy podziałów, różnicowania i dojrzewania, tak jak
komórki linii neutrofilów. Procesy te są regulowane przez czynnik komórek pnia — SCF (stem celi factor), interleukinę 3 (IL-3), czynnik
wzrostowy granulocytów — CSF-G (colony-stimulating factor-granulocyte) i są wspomagane przez interleukinę 5 (IL-5) oraz czynnik wzrostowy
granulocytów i makrofagów — CSF-GM (colony-stimulating factor
granulocyte/macrophage).
Komórki szpiku kostnego pochodzące od komórki macierzystej linii
bazofilów — CFU-Baso (colony-forming unit basophil) — przechodzą
kolejne etapy podziałów, różnicowania i dojrzewania, tak jak komórki
linii neutrofilów. Czynnikami regulującymi te procesy są następujące
cytokiny: czynnik komórek pnia (SCF), interleukiny (IL-3, IL-4, IL-10)
i czynnik wzrostu nerwów (NGF).
286
Krążenie
Ryc. 143
Czynność tkanek krwiotwórczych
287
Ryc. 143. Granulocytopoeza i monocytopoeza w szpiku kostnym czerwonym. Z komórek
ukierunkowanych linii neutrofilów (komórki CFU-G) powstają: mieloblasty, mielocyty
i metamielocyty, które przekształcają się w młode neutrofile, o pałeczkowatym jądrze
komórkowym, przechodzące z miąższu szpiku do zatok szpikowych. Z komórek linii
makrofagów (komórki CFU-M) powstają monoblasty, promonocyty i monocyty przechodzące do krwi krążącej w łożysku krwionośnym. Następnie wychodzą z krwi w łożysku
krwionośnym do tkanek, zamieniając się w makrofagi tkankowe. Czynniki hematopoetyczne:
CSF-GM, CSF-1, CSF-G, IL-6.
Monocytopoeza
Do makrofagów zaliczane są duże komórki żerne występujące w tkankach, w tym również w szpiku kostnym czerwonym. Te same komórki
krążące we krwi zaliczane są do monocytów. Komórki, od których
wywodzą się makrofagi/monocyty są to komórki macierzyste makrofagów
ukierunkowane — komórki CFU-M (colony-forming unit macrophage).
W czasie proliferacji, różnicowania i dojrzewania makrofagów/monocytów dochodzi do ekspresji cząsteczki różnicującej CD 14. Dzięki tej
cząsteczce różnicującej w błonie komórkowej (patrz str. 293) można
dokładnie odróżnić monocyty od innych komórek znajdujących się we
krwi obwodowej, szczególnie od dużych limfocytów, które pod względem
morfologicznym mogą się nie różnić od monocytów.
Komórki CFU-M proliferują, różnicują się i dojrzewają przechodząc
kolejno etapy: monoblastów, promonocytów i monocytów. Procesy te trwają
łącznie od 1 do 3 dni i kończą się przejściem monocytów ze szpiku do krwi
krążącej. Monocyty nie tworzą puli rezerwy szpikowej, lecz po zakończeniu dojrzewania przechodzą do krwi krążącej. Czynnikiem pobudzającym komórki macierzyste CFU-M jest czynnik wzrostowy granulocytów
i makrofagów (CSF-GM) działający wspólnie z czynnikiem wzrostowym
pierwszym (CSF-1), lub też interleukina 6 (IL-6) wraz z czynnikiem
wzrostowym pierwszym (CSF-1).
Trombocytopoeza
Trombocyty, czyli płytki krwi, są to krążące we krwi fragmenty
cytoplazmy megakariocytów. Komórka macierzysta megakariocytów —
CFU-Mk (colony-forming unit megakariocyte) kończy etap proliferacji
w szpiku kostnym czerwonym. Komórka ta dzieli się od dwóch do pięciu
288
Krążenie
Ryc. 144
Czynność tkanek krwiotwórczych
289
Ryc. 144. Granulocytopoeza w szpiku kostnym czerwonym. Z komórek ukierunkowanych
linii eozynofilów (komórki CFU-Eos) i linii bazofilów (komórki CFU-Baso) poprzez
mieloblasty, promielocyty, mielocyty i metamielocyty odpowiednich linii komórkowych
powstają eozynofile i bazofile. Czynniki hematopoetyczne: SCF, IL-3, IL-4, IL-5, IL-10,
CSF-G, CSF-GM, NGF.
Ryc. 145. Trombocytopoeza w szpiku kostnym czerwonym. Fragmenty cytoplazmy megakariocytów krążą we krwi w łożysku krwionośnym jako trombocyty. Czynniki hematopoetyczne: IL-3, IL-6, CSF-GM.
10 — Fizjologia człowieka
290
Krążenie
razy pod wpływem interleukiny 3 (IL-3) i czynnika wzrostowego granulocytów i makrofagów (CSF-GM). Powstające z komórek CFU-Mk promegakarioblasty nie dzielą się mitotycznie, lecz w ich jądrach komórkowych zachodzi replikacja łańcuchów DNA i ich jądra komórkowe stają
się poliploidalne. Proces ten jest wyzwalany przez liczne czynniki wzrostowe hematopoetyczne, przede wszystkim przez interleukinę 3 i 6 (IL-3,
IL-6).
W megakarioblastach i następnie w megakariocytach, które kończą linię tych komórek, dochodzi do 32—64-krotnego nagromadzenia się łańcuchów DNA w stosunku do innych komórek, na skutek powtarzającego
się procesu endomitozy, to jest powielania łańcuchów DNA bez podziału
komórkowego. W tych komórkach wzrasta również stopniowo objętość
cytoplazmy w stosunku do objętości poliploidalnego jądra komórkowego.
Na tym etapie fragmenty cytoplazmy megakariocytów odczepiają się
i krążą we krwi jako trombocyty.
Nieliczne megakariocyty opuszczają szpik kostny, krążą we krwi
i osiadają w śledzionie oraz w płucach. Również wytwarzają trombocyty.
Ich udział w trombocytopoezie nie przekracza 5—7%.
Regulowana jest całkowita objętość trombocytów krążących we krwi,
nie zaś ich liczba w jednostce objętości krwi. Zmniejszenie całkowitej
objętości trombocytów we krwi ma powodować uwalnianie trombopoetyny — TPO (thrombopoietin) i zwiększenie uwalniania do krwi trombocytów. Trombopoetyna nie została zidentyfikowana pod względem
budowy chemicznej i prawdopodobnie jest to mieszanina kilku cytokin.
Zwiększenie całkowitej objętości trombocytów krążących we krwi
powoduje hamowanie ich uwalniania przez megakariocyty do krwi.
Hamowanie trombocytopoezy związane jest z działaniem kilku cytokin,
przede wszystkim czynnika wzrostowego transformującego beta-TGF
(transforming growth factor-beta).
Limfocytopoeza
Limfocytopoeza (lymphocytopoiesis) zachodzi w tkankach limfoidalnych centralnych (central lymphoid tissues), do których zalicza się szpik
kostny i grasicę, oraz w tkankach limfoidalnych obwodowych (peripheral
lymphoid tissues), do których należą węzły chłonne, grudki chłonne
w błonach śluzowych i śledziona.
W szpiku kostnym czerwonym z komórek macierzystych limfoidalnych
powstają komórki, z których część podlega dalszemu dojrzewaniu w szpiku kostnym — są to limfocyty pre-B, a część wychodzi ze szpiku, krąży we
Czynność tkanek krwiotwórczych
Plazmpcyt
w WĘŹLE CHŁONNYM
291
Ryc. 146. Limfocytopoeza w centralnych tkankach limfoidal-
KOMÓRKA PNIA
LIMFOIDALNA
nych: w szpiku kostnym czerwonym i grasicy oraz w obwodowej
tkance limfoidalnej — w węzłach chłonnych. Limfocyty pre-B,
dojrzewając w szpiku kostnym czerwonym, przekształcają się w limfocyty B szpikozależne.
Limfocyty pre-T po przejściu przez grasicę krążą we krwi jako limfocyty T grasiczozależne.
W grasicy w czasie dojrzewania limfocytów pre-T dochodzi do ekspresji wielu cząsteczek
różnicujących (CD). Limfocyty T CD4, T CD8 i T CD3 różnią się funkcją. Limfocyty
NK — naturalni niszczyciele — nie dojrzewają ani w szpiku, ani w grasicy.
292
Krążenie
krwi i zatrzymuje się w grasicy — są to limfocyty pre-T. Część komórek nie dojrzewa ani w szpiku, ani w grasicy i krąży we krwi jako
limfocyty naturalni niszczyciele — limfocyty NK (natural killer cells).
Limfocyty pre-B, dojrzewając w szpiku kostnym, przekształcają się
w limfocyty B szpikozależne wyspecjalizowane w wytwarzaniu immunoglobulin. Limfocyty pre-T, które wyszły w grasicy poza naczynia krwionośne, przekształcają się w tymocyty dzieląc się i dojrzewając początkowo
w części korowej grasicy, a następnie wędrują do jej części rdzennej.
W czasie dojrzewania w korze grasicy komórki nabłonkowe zrębu grasicy
otaczają tymocyty, izolując je od oddziaływań ze strony krwi.
Limfocyty B szpikozależne i limfocyty T grasiczozależne krążą we
krwi, osiadają w tkankach limfoidalnych obwodowych, gdzie się dzielą
i następnie powracają do krwi. Limfocyty wielokrotnie przechodzą z krwi
do chłonki, osiadają w tkankach limfoidalnych obwodowych i powracają
do krwi, tworząc w ten sposób pule limfocytów recyrkulujących (recirculating lymphocytic pool).
Cząsteczki różnicujące
Czynność wszystkich komórek w organizmie, w tym również limfocytów, jest uwarunkowana występowaniem i budową chemiczną receptorów błonowych. W celu rozpoznawania komórek na podstawie ich właściwości czynnościowych, a nie tylko ich cech morfologicznych, wpro-
Czynność tkanek krwiotwórczych
293
wadzono w 1982 roku system rozróżniania leukocytów na podstawie
występowania lub niewystępowania na ich błonie komórkowej tak zwanych markerów lub cząsteczek różnicujących — CD (cluster designations).
Do 1996 roku wykazano występowanie na błonach komórkowych
elementów morfotycznych krwi 155 różnych cząsteczek różnicujących.
Wiele z nich — jako cząsteczki pośredniczące, receptory cytokin lub
cząsteczki adhezyjne — jednocześnie bierze udział w regulacji aktywacji,
proliferacji i różnicowaniu się limfocytów.
W czasie dojrzewania i różnicowania leukocytów, w tym szczególnie
limfocytów, zmienia się ekspresja genów kodujących cząsteczki różnicujące. Na pewnych stadiach dojrzewania może przejściowo występować jednocześnie ponad dwadzieścia różnych cząsteczek różnicujących
(CD) na błonie komórkowej limfocytów. Limfocyty B szpikozależne
opuszczając szpik kostny, mają w błonie komórkowej antygen głównego układu zgodności tkankowej (MHC — major histocompatibility complex), tak jak wszystkie komórki w całym organizmie, oraz cząsteczki
różnicujące CD 10 i CD34. Niektóre z tych limfocytów mają również
cząsteczki różnicujące CD 19, CD22, CD38 i CD40. Zakończenie ekspresji
wszystkich tych cząsteczek różnicujących kończy proces dojrzewania
limfocytów pre-B w limfocyty B, które krążąc we krwi, osiadają w tkankach limfoidalnych obwodowych. W limfocytach B dochodzi również do
ekspresji genów dla immunoglobulin.
Limfocyty pre-T, tak jak limfocyty pre-B, wywodzą się z komórek
macierzystych limfoidalnych o cząsteczkach różnicujących CD34. W
opuszczających szpik kostny limfocytach pre-T zachodzi ekspresja
cząsteczek różnicujących CD7, CD34 i CD45.
Limfocyty pre-T po opuszczeniu szpiku kostnego krążą we krwi
w układzie krążenia i zatrzymują się w korze grasicy — przekształcając
się w tymocyty. W tych komórkach w korze grasicy dochodzi do ekspresji
cząsteczek różnicujących: CD2, CD3, CD4, CD7 i CD8. Większość
tymocytów ginie przed przemieszczeniem się do rdzenia grasicy. Giną
wszystkie tymocyty, w których doszło do jednoczesnej ekspresji CD4
i CD8. Przeżywają te, w których wystąpiła ekspresja jednej z dwóch
cząsteczek różnicujących CD4 lub CD8. Grasicę opuszczają limfocyty
T grasiczozależne jako nie w pełni jeszcze dojrzałe komórki — w około
60% limfocyty T CD4 i w około 20—30% limfocyty T CD8.
Cząsteczka różnicująca CD4 jest to glikoproteina należąca do immunoglobulin o czterech zewnątrzkomórkowych domenach podobnych
do immunoglobulin i o masie cząsteczkowej 55 kDa. Cząsteczka różnicująca CD8 jest to również glikoproteina, heterodimer o dwóch łań-
294
Krążenie
cuchach alpha i beta i o jednej domenie podobnej do immunoglobuliny
oraz o masie cząsteczkowej 32—34 kDa.
Trzeci rodzaj limfocytów stanowią limfocyty naturalni niszczyciele —
limfocyty NK. We wszystkich tych limfocytach dochodzi do ekspresji
cząsteczek różnicujących CD 16 i CD56 oraz kilku innych cząsteczek
różnicujących. Limfocyty NK nie podlegają dojrzewaniu w obwodowych
tkankach limfoidalnych.
I Grasica
Limfocyty pre-T po przejściu do krwi ze szpiku kostnego krążą wraz
z krwią w całym organizmie. Z krwi przepływającej przez grasicę limfocyty
pre-T wychodzą i osiadają w korze grasicy sąsiadującej z torebką grasicy.
W korze grasicy pod torebką limfocyty tworzą warstwę dużych komórek
dzielących się, czyli tymocytów. Następnie tymocyty stopniowo przenikają w głąb kory grasicy do pogranicza pomiędzy korą i rdzeniem grasicy.
W tym czasie zachodzi proces dojrzewania przemieszczających się tymocytów. Przemieszczając się stykają się z komórkami zrębu grasicy, w tym
z komórkami nabłonkowo-siateczkowymi (epithelial-reticular cells), z komórkami siateczki, komórkami bariery i z makrofagami.
Komórki zrębu grasicy oddziałują na dojrzewanie tymocytów w wyniku:
• wydzielania tymozyny (thymosyne) i innych czynników grasiczych,
które wzmagają lub osłabiają dojrzewanie, dzielenie się i przemiesz
czanie tych komórek;
• tworzenia woreczków (sacs, enclosure), które izolują dojrzewające
tymocyty i utrzymują duże stężenie czynników regulujących proces
dojrzewania;
• kontroli występowania w błonie komórkowej ludzkiego leukocytarnego
antygenu — HLA (human leukocyte antygen) — dzięki ekspresji ge
nów głównego układu zgodności tkankowej — MHC (major histocompatibility complex).
Pojawienie się w błonie komórkowej limfocytów własnego antygenu
HLA pozwala im odróżniać komórki własnego organizmu od komórek
organizmów obcych. Wszystkie komórki organizmu człowieka na błonie
komórkowej mają cząsteczki glikoproteiny zmienne osobniczo i kodowane przez geny głównego układu zgodności tkankowej. Własne komórki
Czynność tkanek krwiotwórczych
295
Ryc. 148. Budowa schematyczna grasicy. (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
nie są niszczone przez limfocyty T. Obce komórki mające na błonie
komórkowej obce cząsteczki glikoproteiny są rozpoznawane i niszczone
przez limfocyty T.
W części rdzennej grasicy — podobnie jak w korze grasicy — występują
komórki nabłonkowate i tymocyty. Pomiędzy tymocytami w korze
i w rdzeniu grasicy występują jednak istotne różnice. Tymocyty w korze
grasicy nie są jeszcze całkowicie dojrzałe i giną pod wpływem steroidów
kory nadnerczy, szczególnie pod wpływem kortyzolu. Tymocyty w rdzeniu
grasicy są całkowicie dojrzałe i wykazują oporność na działanie steroidów
kory nadnerczy.
W procesie dojrzewania w grasicy około 95% tymocytów ginie, a tylko
około 5% tych komórek jest uwalniane z grasicy do krwi jako limfocyty T
grasiczozależne, immunologicznie kompetentne, lecz jeszcze niecałkowicie
dojrzałe komórki. Krążąc we krwi, komórki te zatrzymują się w śledzionie,
296
Krążenie
gdzie podlegają końcowemu ukszałtowaniu tak, aby następnie powrócić
do krwi krążącej i utworzyć pulę limfocytów T grasiczozależnych recyrkulujących i długo żyjących.
I Węzły chłonne i grudki chłonne
Węzły chłonne i grudki chłonne w błonach śluzowych tworzą tkankę
limfoidalną obwodową (peripheral lymphoid tissue). Do węzłów chłonnych napływają limfocyty wraz z chłonką. Są to zarówno limfocyty B, jak
i limfocyty T stanowiące pulę limfocytów recyrkulujących — RLP
(recirculating lymphocyte pool). Limfocyty T wychodzą z chłonki i zatrzymują się w miąższu węzłów chłonnych w strefie przykorowej —
grasiczozależnej (T-cell region). Dzielą się tam, tworząc ośrodki rozmnażania. Następnie opuszczają węzły chłonne, odpływając wraz z chłonką, z którą trafiają do krwi.
W węzłach i grudkach chłonnych błon śluzowych limfocyty B zatrzymują się w strefie podtorebkowej — szpikozależnej (B-cell region). W
ośrodkach rozmnażania limfocytów B zostaje zakończony proces
dojrzewania tych komórek. Limfocyty B, podobnie jak limfocyty T,
dostają się z chłonką do krwi, recyrkulując wielokrotnie pomiędzy chłonką i krwią, ale większość swojego życia spędzają w węzłach chłonnych.
Ryc. 149. Budowa schematyczna węzła chłonnego. (Reprodukowano za zezwoleniem z W.
Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Czynność tkanek krwiotwórczych
W kontroli limfocytopoezy biorą udział:
czynniki wytwarzane przez grasicę (thymic factors), powodując dojrzewanie limfocytów T;
hormony gruczołu tarczowego, trijodotyronina (T 3) i tyroksyna (T 4),
wzmagające limfocytopoezę;
hormon wzrostu (GH) i czynniki wzrostowe, powodujące wzrost tkanki
limfoidalnej w organizmie;
hormony kory nadnerczy hamujące mitozę komórek zarówno w centralnych, jak i w obwodowych narządach limfoidalnych.
I Śledziona
Przez śledzionę (Hen), która jest małym narządem, przepływa krew
w ilości około 5% pojemności minutowej serca w spoczynku. Krew
tętnicza przepływa przez miazgę białą śledziony (pulpa lienis alba), krew
żylna zaś odpływa od miazgi czerwonej śledziony (pulpa lienis rubra).
Miazgę białą tworzą grudki chłonne należące do obwodowego układu
limfoidalnego. Pomiędzy tętniczkami i żyłkami śledziony krew przepływa
przez zatoki śledziony, które nie mają zachowanej ciągłości śródbłonka
naczyniowego.
Śledziona tworzy filtry dla elementów morfotycznych krwi powstających w innych tkankach hematopoetycznych. Pierwsze filtry, które
zatrzymują wybiórczo limfocyty T i limfocyty B, znajdują się w miazdze
białej. Zatrzymane limfocyty tworzą oddzielne ośrodki rozmnażania
limfocytów T grasiczozależnych i limfocytów B szpikozależnych.
Strefa przejściowa pomiędzy miazgą białą i miazgą czerwoną stanowi
następny filtr, który zatrzymuje uszkodzone erytrocyty, granulocyty i trombocyty, jak również mikroorganizmy, które przeniknęły do krwi. Monocyty zatrzymane w strefie brzeżnej, pomiędzy miazgą białą i czerwoną
przekształcają się w śledzionowe makrofagi.
Miazga czerwona stanowi ostatni filtr zatrzymujący wszystkie elementy
morfotyczne krwi: erytrocyty, granulocyty, monocyty, trombocyty i limfocyty.
Po jednokrotnym przepłynięciu krwi przez śledzionę zatrzymywana jest
w niej tylko nieznaczna liczba elementów morfotycznych. Ponieważ krew
stale przepływa przez śledzionę, narząd ten ma istotny wpływ na liczbę
krążących we krwi elementów morfotycznych.
298
Krążenie
Ryc. 150. Budowa schematyczna śledziony. (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
W śledzionie zachodzą następujące procesy:
• wytwarzanie limfocytów;
• niszczenie trombocytów;
• rozpad starych erytrocytów krążących we krwi. Komórki należące do
układu siateczkowo-śródbłonkowego śledziony (RES) fagocytują roz
padające się erytrocyty, niszczą ich otoczkę i uwalniają z ich wnętrza
hemoglobinę, która podlega dalszemu rozpadowi.
Przepływ krwi przez śledzionę zatrzymuje się w czasie ostrego niedotlenienia organizmu. Tętnice śledzionowe obkurczają się pod wpływem
wzmożonej impulsacji wspołczulnej, jak również dopływającej adrenaliny
z krwią tętniczą.
I Elementy morfotyczne krwi
I Erytrocyty
Erytrocyty, czyli krwinki czerwone, w życiu pozapłodowym wytwarzane są przez szpik kostny. Krążą we krwi obwodowej około 120 dni, okres
ich połowicznego rozpadu (T 1^) — oznaczany metodą znakowania
erytrocytów izotopem chromu 51Cr — wynosi 28 dni.
Elementy morfotyczne krwi
299
Erytrocyty spełniają jedną z podstawowych funkcji w organizmie —
transportują cząsteczki tlenu z płuc do tkanek. Charakteryzuje je wiele
oznaczanych wielkości. Są to: liczba w 1 L krwi obwodowej — Erys
(erythrocytes), wskaźnik hematokrytu — Het, zawartość hemoglobiny — Hb, średnia objętość — MCV (mean corpuscular volume), średni
ciężar hemoglobiny — MCH (mean corpuscular hemoglobin), średnie
procentowe stężenie hemoglobiny — MCHC (mean corpuscular hemoglobin concentration) i średnia średnica erytrocytu — MCD (mean celi
diameter).
Średnica erytrocytu wynosi przeciętnie około 8 um i jest większa od
średnic naczyń włosowatych w różnych tkankach. Erytrocyt, przepływając
przez naczynia włosowate o mniejszej średnicy niż 8 um, odkształca
się i następnie powraca do kształtu dwuwklęsłego dysku. Wielokrotny
powrót
Tabela 18
Wartości średnie charakteryzujące erytrocyty
Mężczyźni
zakres
średnia
Kobiety
średnia
zakres
prawidłowych
wartości
prawidłowych
wartości
Liczba erytrocytów w 1 L
krwi obwodowej x 1012
Wskaźnik hematokrytu
(Het)
5,21
4,52—5,90
4,60
4,10—5,10
0,46
0,42—0,50
0,40
0,36—0,45
Zawartość hemoglobiny
(HB) w mmol w 1 L krwi
(w g w dL)
10,0
15,7
Średnia objętość erytrocytu (MCV) w fL
88,0
Średnia masa hemoglobiny w erytrocycie
(MCH) w fmol (pg)
Średnie stężenie hemoglobiny w erytrocycie
(MCHC) w mmol w 1 L
(w g w dL)
8,7—11,2
14,0—17,5
8,7
13,8
7,5—10,0
12,3—15,3
80,0—96,1
88,0
80,0—96,1
1,89
(30,4)
1,71—2,06
(27,5—33,2)
1,89
(30,4)
1,71—2,06
(27,5—33,2)
21,35
(34,4)
20,73—22,3
(33,4—35,5)
21,35
(34,4)
20,73—22,03
(33,4—35,5)
Het
— hematocrit, ratio
MCV — mean celi volume
MCH — mean corpuscular hemoglobin
MCHC — mean corpuscular hemoglobin concentration
300
Krążenie
do właściwego kształtu krążących we krwi erytrocytów zapewnia cytoszkielet erytrocytu. Tworzy go sieć cząsteczek spektryny (spektrin), białka
zbudowanego z dwóch łańcuchów polipeptydowych: alpha-podjednostki,
o masie cząsteczkowej 260 kDa, i beta-podjednostki, o masie cząsteczkowej 246 kDa. Cząsteczki spektryny połączone są ze sobą przy udziale
cząsteczek aktyny i białka 4.1. Do wewnętrznej powierzchni otoczki
przymocowane są zaś za pośrednictwem cząsteczek białka ankiryny
(ankyrin).
Otoczka erytrocytów jest błoną półprzepuszczalną, przez którą cząsteczki wody z łatwością dyfundują. W fizjologicznym roztworze 0,9%
chlorku sodowego ta sama ilość cząsteczek wody dyfunduje przez otoczkę
erytrocytu w obu kierunkach, to jest do jego wnętrza i z erytrocytu na
zewnątrz. Ciśnienie osmotyczne panujące w erytrocycie i wywierane przez
0,9% roztwór fizjologiczny NaCl jest jednakowe. Równa się 310 mOsm,
czyli jest izotoniczne.
Przy zwiększonym stężeniu NaCl płyn staje się hipertoniczny, cząsteczki wody dyfundują na zewnątrz i erytrocyty obkurcząją się. W wyniku
zmniejszenia stężenia NaCl płyn staje się hipotoniczny, cząsteczki wody
dyfundują do wnętrza i powiększają swoje wymiary aż do momentu,
w którym otoczka pęka. Pękanie otoczek zaczyna się w 0,48% roztworze
NaCl, a w 0,33% roztworze hemoliza jest całkowita.
Hemoglobina
Średnie procentowe stężenie hemoglobiny (Hb) w poszczególnych
erytrocytach — MCHC (mean corpuscular hemoglobin concentration) —
wynosi 34%. Hemoglobina o masie cząsteczkowej 64,5 kDa jest zbudowana z białka — globiny, składającego się z 4 łańcuchów polipeptydowych
0 masie cząsteczkowej około 15,5 kDa i z 4 cząsteczek hemu. Każda
z czterech cząsteczek hemu jest połączona z jednym łańcuchem polipeptydowym. U ludzi dorosłych 97% stanowi hemoglobina A x , 2,5%
hemoglobina A 2 i około 0,5% hemoglobina płodowa, czyli F (fetal
hemoglobin).
W skład hemoglobiny A1 wchodzą 2 łańcuchy polipeptydowe typu
alpha, zawierające po 141 aminokwasów, i 2 łańcuchy typu beta, z których
każdy ma 146 aminokwasów. Hemoglobina A 2 ma 2 łańcuchy alpha
1 2 łańcuchy delta, a Hb F ma 2 łańcuchy alpha i 2 łańcuchy gamma.
Hemoglobina płodowa w życiu płodowym występuje w największym
procencie i zaraz po urodzeniu jej ilość szybko się zmniejsza. Zawartość
Hb F około 0,5% ustala się w 6 miesiącu życia.
Elementy morfotyczne krwi
301
Ryc. 151. Schematyczna budowa cząsteczki hemoglobiny A,. HbA, zbudowana jest z dwóch
łańcuchów polipeptydowych alpha i dwóch łańcuchów polipeptydowych beta oraz czterech
cząsteczek hemu. Łańcuchy alpha i beta łączą liczne wiązania. Łańcuchy polipeptydowe są
odsunięte od siebie, tak aby lepiej była widoczna ich struktura.
Oksyhemoglobina
Cząsteczka prostetyczna hemoglobiny — hem, zawierająca atom żelaza
dwuwartościowego Fe2+, wiąże się z jedną cząsteczką tlenu, tworząc
hemoglobinę związaną z tlenem, czyli oksyhemoglobinę (Hb4O8).
Stopień wysycenia hemoglobiny tlenem, czyli równowaga pomiędzy
wiązaniem hemoglobiny z tlenem i uwalnianiem O 2 z oksyhemoglobiny,
zależy od:
•
•
•
•
prężności tlenu — POl;
temperatury krwi;
prężności dwutlenku węgla — Pco2'
stężenia jonów wodorowych, czyli pH.
302
Krążenie
Wraz ze zwiększeniem prężności tlenu we krwi zwiększa się wysycenie Hb tlenem. Przy prężności tlenu we krwi (Po 2 ) odpowiadającej
12,7 kPa już 97,5% Hb jest całkowicie wysycone tlenem. Ciśnienie
parcjalne O2 w powietrzu wdychanym wynosi 21,1 kPa.
Wraz ze spadkiem temperatury krwi i przy zachowanej tej samej
prężności tlenu (P Q2) zwiększa się wiązanie Hb z tlenem.
Ryc. 152. Krzywa dysocjacji hemoglobiny
przedstawia równowagę pomiędzy wiązaniem Hb i O2 i uwalnianiem O2 z Hb4O8.
Na osi rzędnych wysycenie hemoglobiny
tlenem w procentach. Na osi odciętych
ciśnienie parcjalne tlenu w mm Hg i kPa.
Ryc. 153. Wpływ temperatury na dysocjację hemoglobiny. Na osi rzędnych wysycenie Hb tlenem w procentach. Na osi odciętych ciśnienie parcjalne O2 w mm Hg
ikPa.
Elementy morfotyczne krwi
303
Zwiększenie prężności dwutlenku węgla (CO 2) we krwi zmniejsza
stopień wysycenia Hb tlenem. Zwiększenie stężenia wolnych jonów
wodorowych we krwi, czyli zmniejszenie pH krwi, również zmniejsza
stopień wysycenia Hb tlenem.
We krwi w naczyniach włosowatych pęcherzyków płucnych w warunkach dużej Po2, niższej temperatury niż w innych tkankach (35°C), małej
Pco2 i wyższego pH krwi prawie 100% Hb zostaje wysycone tlenem.
Zasadniczą rolą hemoglobiny jest jej udział w transporcie tlenu z płuc
do tkanek, jako Hb 4O8. Transportuje ona w pewnym stopniu również
dwutlenek węgla z tkanek do płuc. Hemoglobina i mioglobina związane
z tlenem stanowią magazyn tlenu w organizmie. Magazyn ten zaspokaja
zapotrzebowanie na tlen tylko na kilka sekund.
Ryc. 154. Wpływ dwutlenku węgla o różnym ciśnieniu cząstkowym (P co = 2,0;
5,3; 9,3 kPa) na dysocjację hemoglobiny.
Na osi rzędnych wysycenie Hb tlenem
w procentach. Na osi odciętych ciśnienie
parcjalne O2 w mm Hg i kPa.
Pod wpływem związków utleniających dwuwartościowy atom żelaza
(Fe2+) w cząsteczce hemu zostaje zamieniony na atom trójwartościowy
(Fe3+), tworząc methemoglobinę. Nie ma ona zdolności do transportu O2,
czyli jego wiązania i uwalniania.
Tlenek węgla (CO) jest związkiem współzawodniczącym z tlenem przy
wiązaniu się z hemoglobiną. Tlenek węgla ma około 200 razy większe
powinowactwo do Hb i wypiera O 2 z oksyhemoglobiny, zajmując jego
miejsce i tworząc karboksy hemoglobinę.
304
Krążenie
Hemoglobina jest syntetyzowana w szpiku kostnym w komórkach
potomnych proerytroblastu, w erytroblastach zasadochłonnych i polichromatofilnych.
Dojrzałe erytrocyty, czyli normocyty, krążą we krwi około 120 dni i
następnie rozpadają się w układzie siateczkowo-śródbłonkowym (RES —
reticuloendothelial system), w śledzionie i wątrobie, uwalniając ze swojego
wnętrza hemoglobinę. Cząsteczka globiny jest rozkładana na aminokwasy,
hem zaś jest zamieniany na biliwerdynę, a odczepione atomy żelaza
powracają do osocza krwi. Żelazo zostaje ponownie wykorzystane do
syntezy hemoglobiny. Biliwerdyna metabolizowana jest do bilirubiny,
która w komórkach wątrobowych wiąże się z kwasem glukuronowym
i w tej postaci jest wydalana z żółcią do dwunastnicy.
Grupy krwi
W błonach komórkowych elementów morfotycznych krwi występują
antygeny, w osoczu krwi u innych ludzi zaś obecne są naturalne przeciwciała przeciwko tym antygenom. Spośród wielu odkrytych grup krwi
praktyczne znaczenie mają grupy krwi układu ABO, Rh i ludzkie antygeny
leukocytarne.
Antygeny grupowe ABO występują w otoczce erytrocytów. Są to cząsteczki polisacharydów stanowiące substancje grupowe A, B i H. Na
podstawie ich występowania został dokonany podział na cztery główne grupy krwi: A, B, AB i 0. U ludzi, u których antygen A znajduje
się w otoczce erytrocytów, w osoczu występuje naturalne przeciwciało
anty-B, czyli beta. W grupie B z antygenem B w otoczce erytrocytów
w osoczu są przeciwciała naturalne anty-A, czyli alpha. W grupie AB oba
antygeny A i B znajdują się w otoczce erytrocytów, w osoczu zaś nie
występują naturalne przeciwciała. U osób z grupą 0 substancja grupowa H ma bardzo słabe właściwości antygenowe, czyli praktycznie nie mają
one antygenu, natomiast w ich osoczu krwi występują naturalne
przeciwciała anty-B, czyli beta, i anty-A, czyli alpha.
Antygen A nie jest jednorodny, dzieli się na Aj i A2. Dlatego w praktyce wyróżnia się sześć grup krwi układu ABO: At, A2, B, AXB, A2B i 0.
Niezależnie od podziału krwi na sześć grup na podstawie występowania antygenów A, B i H istnieje podział na dwie grupy układu
Rh: Rh-dodatnią i Rh-ujemną. W otoczkach erytrocytów ludzi z grupą
Rh-dodatnią występuje antygen D, a u ludzi z grupą Rh-ujemną brak jest
tego antygenu. Przetoczenie krwi Rh-dodatniej biorcy Rh-ujemnemu
powoduje pojawienie się w jego osoczu izoprzeciwciał anty-D. Ponowne
Elementy morfotyczne krwi
305
przetoczenie krwi Rh-dodatniej temu biorcy prowadzi do niszczenia
erytrocytów dawcy przez przeciwciała powstałe po pierwszej niezgodnej
grupowo transfuzji.
Antygen D odgrywa również dużą rolę w konflikcie serologicznym
pomiędzy Rh-ujemną matką i Rh-dodatnim płodem. W czasie pierwszej
ciąży może dojść do izoimmunizacji i powstania przeciwciał anty -D
u matki. W czasie następnej ciąży u Rh-dodatniego płodu może dochodzić
do niszczenia krwinek przez izoprzeciwciała anty-D matki.
Pozostałe antygeny układu Rh, a jest ich ponad czterdzieści, mają małe
praktycznie znaczenie, ale oznacza się również antygeny C, c, E i e.
Klasyfikacji grup krwi dokonuje się jednocześnie na podstawie występowania antygenów układu ABO i układu Rh. Wśród wielu innych
układów grupowych znaczenie praktyczne w transfuzjologii mają antygeny: S — z układu MNSs, kell — z układu Kell oraz Jk — z układu Kidd,
które mogą wywołać izoimmunizację i odczyny poprzetoczeniowe. W celu uniknięcia ewentualnych powikłań, przed przetoczeniem krwi przeprowadza się próbę krzyżową, czyli inkubację erytrocytów dawcy z osoczem biorcy i erytrocytów biorcy z osoczem dawcy. Po inkubacji obu
próbek sprawdza się, czy nie nastąpiła w nich aglutynacja. Występowanie
aglutynacji dyskwalifikuje krew do przetoczenia, mimo zgodności w zakresie grup głównych.
Na powierzchni leukocytów i trombocytów występują również antygeny grupowe. Ludzkie antygeny leukocytarne — HLA (human leukocyte
antigens), czyli główne antygeny zgodności tkankowej — MHC (major
Tabela 19
Podziat grup krwi na podstawie występowania antygenów grupowych A, B i 0
Grupa krwi
Antygen grupowy w
otoczce
erytrocytów
Przeciwciała w
osoczu
Częstość występowania
grup w Polsce*
A,
A2
A,
A2
anty-B
anty-B
B
anty-Aj
Aj+B
brak
31,5
9,5
19,0
6,4
1,6
32,5
B
AXB
A2B
0
A2 + B
brak
—
+ anty-Aj
anty-B
* Według Seyfriedowej i Walewskiej (1979 r.) w %.
306
Krążenie
histocompatibility complex), są rozpoznawane przez receptory na błonie
komórkowej limfocytów T. Receptory te różnicują własne antygeny
układu HLA od antygenów obcych. Układ HLA ma zasadnicze znaczenie
w transplantologii. Od zgodności grupowej zależy przyjęcie, lub też
odrzucenie przeszczepionego narządu.
I Leukocyty
Leukocyty, czyli krwinki białe, znajdują się we krwi obwodowej w liczbie około 7,5 x 109 w 1 L krwi. Jako granice fizjologiczne przyjęto wartości
od 4x 109 do 10 x 109 krwinek białych w 1 L krwi obwodowej.
Do leukocytów zalicza się:
• granulocyty zawierające w cytoplazmie ziarnistości wytwarzane w szpi
ku kostnym czerwonym (medulla ossium rubra);
• limfocyty wytwarzane w szpiku kostnym czerwonym, w grasicy (thymus), w węzłach chłonnych {nodi lymphatici), w śledzionie {Heń)
i w grudkach chłonnych przewodu pokarmowego (Jolliculi lymphatici
canalis alimentarii);
• monocyty pochodzące ze szpiku kostnego czerwonego, które po przej
ściu do tkanek i narządów stanowią część komórek układu siatecz kowo-śródbłonkowego — RES.
krwi obwodowej
Tabela 20
Leukocyty
we
Bezwzględna liczba
Rodzaje leukocytów
w 1 L krwi x 109
Względna liczba
odsetkowa
średnia
zakres
prawidłowych
wartości
7,4
4,5—11,0
4,4
0,20
1,8—7,7
0—0,45
59
3
40—70
0—4
bazofile
Limfocyty
0,04
2,5
0—0,2
1,0—4,8
0,5
33,5
0—1,8
22—44
Monocyty
0,30
0,1—0,8
4
2—7
Całkowita liczba
leukocytów
Granulocyty
neutrofile
eozynofile
średnia
zakres
prawidłowych
wartości
Elementy morfotyczne krwi
307
Granulocyty
Granulocyty dojrzałe, czyli segmentowane — PMN (polymorphonuclear), wywodzą się z komórek szpiku kostnego i dzielą się na:
• granulocyty obojętnochłonne o ziarnistosciacłi cytoplazmatycznych
obojętnochłonnych — neutrofile. Stanowią od 35 do 71% wszystkich
krążących we krwi leukocytów;
• granulocyty kwasochłonne o ziarnistościach kwasochłonnych — eozynofile. Stanowią od 0 do 8% wszystkich leukocytów;
• granulocyty zasadochłonne o ziarnistościach zasadochłonnych — bazofile. Stanowią od 0 do 2% wszystkich leukocytów.
Okres połowicznego krążenia neutrofilow we krwi obwodowej wynosi
około 7 godzin. Po przejściu przez ścianę naczyń włosowatych do tkanek,
dzięki zdolności do diapedezy, nie powracają już więcej do krwi krążącej.
Całkowita pula neutrofilow krwi — TBNP (total blood neutrophil
pool) — wynosi około 70 x 109 komórek. Średnio nieco mniej niż połowa
TBNP przypada na pulę neutrofilow swobodnie krążących — CNP
Ryc. 155. Naczynie włosowate wypełnione granulocytami należącymi do
puli granulocytów swobodnie krążących (CGP) i puli granulocytów przyściennych (MGP). Po stronie prawej
granulocyty opuszczające krew krążącą
wykazują diapedeze, chemotaksję i fagocytozc.
308
Krążenie
(circulating neutrophil pool) — i nieco więcej niż połowa przypada n
neutrofile pozostające blisko ścian naczyń krwionośnych, czyli tzw. pu
neutrofilow przyściennych — MNP (marginal neutrophil pool).
Chwilowe i bardzo szybkie zwiększenie liczby neutrofilow krążącyc
może nastąpić w wyniku przesunięcia ich z puli przyściennej (MNP) d
puli swobodnie krążących (CNP). Obserwuje się to w czasie intensywne
pracy mięśni szkieletowych i pod wpływem wydzielonych hormoncn
rdzenia nadnerczy. Całkowita pula neutrofilow krwi wówczas nie zmieni:
się.
Całkowita pula neutrofilow krwi może ulec zwiększeniu pod wpływen
toksyn bakteryjnych lub czynników uwalnianych przez same neutrofile
Czynniki te powodują przejście neutrofilow z rezerwy szpikowej (reserv(
pool) do krwi krążącej. W ciągu kilku godzin całkowita liczba neutrofilów może wówczas zwiększyć się nawet do dziesięciu razy. Liczba
segmentów jąder neutrofilow wynosi od 1 do 5. Najwięcej komórek ma
trzy segmenty.
Neutrofile utrzymują równowagę pomiędzy makroorganizmem człowieka i drobnoustrojami. Inwazja drobnoustrojów do środowiska wewnętrznego organizmu jest powstrzymywana w ciągu całego życia człowieka przez neutrofile. Zmniejszenie bezwzględnej liczby neutrofilow
Ryc. 156. Wzór Arnetha — procent występujących we krwi neutrofilow o jądrze: pałe-
czkowatym, dwu-, trzy-, cztero- i pięciosegmentowym. Przesunięcie w lewo — przewaga
młodych komórek. Przesunięcie w prawo — przewaga starszych komórek. (Reprodukowano
za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa
1997.)
Elementy morfotyczne krwi
309
Tabela 21
Interleukiny
Komórki
wydzielające
Główne działanie
Skrót
Masa
cząsteczkowa kDa
Liczba
aminokwasów
IL-la
IL-ip
17,7
159
makrofagi różnorodne pobudza wydzielanie czynników wzrostowych
komórki somatyczne hematopoetycznych
IL-2
23
133
limfocyty T CD4
pobudza proliferacje i aktywację limfocytów
T, B i NK
IL-3
28
134
limfocyty T
pobudza czynni k wzrostowy hematopoetyczny
IL-4
19
153
limfocyty T CD4
aktywuje limfocyty B, pobudza wydzielanie
immunoglobulin
IL-S
40
134
limfocyty T
aktywuje limfocyty T cytotoksyczne i wytwarzanie eozynofilów
IL-6
28
187
fibroblasty
makrofagi
pobudza wydzielanie czynników wzrostowych
hematopoetycznych i trombocytopoezę
IL-7
25
152
komórki zrębu szpiku
kostnego czerwonego
pobudza wzrost limfocytów pre-B i pre-T w
szpiku kostnym czerwonym
IL-8
8
72
makrofagi fibroblasty
komórki śródbtonka
naczyń krwionośnych
aktywuje neutrofile i wydzielanie leukotrienów, wzmaga chemotaksję limfocytów T i
bazofilów
IL-9
40
300
limfocyty T CD4
wzmaga erytrocytopoezę, działając wraz z erytropoetyną na komórki szpikowe BFU-E
IL-10
18
178
limfocyty T CD4
hamuje wydzielanie i nterleukin i cytokin przez
monocyty i makrofagi
ILrll
23
199
fibroblasty komórki
pobudza proliferację limfocytów T i B oraz
zrębu szpiku kostnego erytropoezę, działając na komórki macierzyste
czerwonego
erytroidalne, pobudza trombocytopoezę
IL-12
70
makrofagi
pobudza proliferację limfocytów T i B oraz
progenitorowych komórek w szpiku kostnym
czerwonym
IL-13
10
limfocyty T
aktywuje monocyty i limfocyty B, pobudza
wydzielanie immunoglobulin
IL-14
limfocyty T
wzrost i różnicowanie limfocytów B
IL-15
różnorodne komórki
somatyczne
przyspiesza różnicowanie komórek krwiotwórczych i powstawanie komórek NK
IL-16
limfocyty
wytwarzanie czynnika chemotaktycznego dla
limfocytów i eozynofilów
IH7
limfocyty T
pobudza wytwarzanie cytokin przez komórki:
nabłonka, śródbłonka i fibroblasty
310
Krążenie
poniżej 1,5 xl O9 w 1 L krwi obwodowej jest niebezpieczne, ponieważ
liczba ta jest niedostateczna do obrony ustroju przed inwazją drobnoustrojów.
Czynność neutrofilów związana jest z ich właściwościami przemieszczania się, fagocytozy, degranulacji, oddychania wybuchowego oraz
odbioru i wysyłania humoralnych sygnałów w postaci cytokin.
Krążące we krwi neutrofile przyczepiają się do komórek śródbłonka
w naczyniach włosowatych i wychodzą z łożyska krwionośnego (diapedeza), kierując się do ognisk zapalnych, ognisk rozmnażania się bakterii, do
martwych tkanek i innych obcych ciał. Neutrofile przemieszczają się do
miejsc, w których tworzą się chemokiny — substancje wytwarzane przez
uszkodzone lub zaktywizowane komórki. Neutrofile pożerają, czyli fagocytują, bakterie i uszkodzone komórki, i trawią je w lizosomach za pomocą
enzymów hydrolitycznych.
Po dotarciu do ognisk zapalnych w neutrofilach zachodzi reakcja
degranulacji. Ziarnistości cytoplazmatyczne zawierające enzymy i liczne
aktywne substancje w procesie egzocytozy uwalniane są do środowiska
otaczającego neutrofile.
Neutrofile niszczą mikroorganizmy, wytwarzając wolne tlenowe rodniki przy udziale fosforanu dinukleotydu dihydronikotynoamido-adeninowego — NADPH (dihydronicotinamide adeninę dinucleotide phosphate). W czasie tzw. oddychania wybuchowego NADPH odbiera jeden
elektron od cząsteczki tlenu i powstaje anion o wolnym rodniku:
NADPH + 2O 2 -» NADP++20- + H+
Anion ten ma słabe działanie bakteriobójcze, ale pod wpływem enzymów powstaje silnie działający nadtlenek wodoru i tlen cząsteczkowy:
Nadtlenek wodoru nie tylko sam wykazuje działanie bakteriobójcze, ale
w reakcji katalizowanej przez jony żelazowe wytwarzany jest wodorotlenek
(HO) — najsilniej działający utleniacz:
Oddychanie wybuchowe nasila się pod wpływem cytokin takich, jak
czynniki wzrostowe hematopoetyczne CSF-G i CSF-GM. Neutrofile są
aktywizowane przez interferon gamma (IFN-^), czynnik martwicy nowotworów alpha (TNF-a) i interleukiny 1,4 i 8 (IL-1, IL-4, IL-8). Neutrofile
Elementy morfotyczne krwi
311
same również wydzielają cytokiny takie, jak: interleukinę 1, 6 i 8 (IL-1,
IL-6, IL-8), czynnik martwicy nowotworów alpha (TNF-a) i czynniki
wzrostowe hematopoetyczne CSF-G i CSF-M.
Eozynofile (eosinophils) wykazują te same właściwości diapedezy,
chemotaksji i fagocytozy co neutrofile. W warunkach fizjologicznych
eozynofile inaktywują substancje wywołujące odczyn zapalny, natomiast
w warunkach rozwiniętego procesu patologicznego nasilają odczyn zapalny. W stosunku do różnych pasożytów wykazują podobne reakcje do
reakcji neutrofilów w stosunku do bakterii.
Liczne czynniki chemotaktyczne aktywują eozynofile. Do tych czynników zalicza się: leukotrieny B4 (LTB4), czynnik aktywujący trombocyty — PAF (platelet-ativating factor), histaminę, interleukinę 5 (IL-5)
oraz substancje uwalniane przez pasożyty. Same zaś eozynofile wydzielają
leukotrieny C4 (LTC4) i B4 (LTB4) i czynniki wzrostowe transformujące
alpha (TGF-a) i beta (TGF-p).
Tabela 22
Cytokiny
Skrót
lub
symbol
Nazwa
Masa
cząsteczkowa
Komórki
wydzielające
Główne działanie
leukocyty
fibroblasty
leukocyty
fibroblasty
limfocyty T
limfocyty NK
makrofagi
komórki somatyczne limfocyty
TiB
przeciwwirusowe, hamuje
proliferację
przeciwwirusowe, hamuje
proliferację
przeciwwirusowe, hamuje
proliferację
zapalne, wzmaga wytwarzanie immunoglobulin
limfocyty T
zapalne, wzmaga wytwarzanie immunoglobulin
wzmaga podziały mitotyczne
IFN-a
interferon alpha
18
IFN-P
interferon beta
20
IFN-y
interferon gamma
25
TNF-a czynnik martwicy
nowotworów alpha
17
TNF-P czynnik martwicy
nowotworów beta
PDGF czynnik wzrostu z
trombocytów
18
TGF-P czynnik wzrostowy
transformujący beta
25
Pominięte zostały interleukiny.
trombocyty
trombocyty
limfocyty T
komórki somatyczne
przeciwzapalne
312
Krążenie
Bazofile (basophils) uczestniczą w reakcjach związanych z bezpośredn:
nadwrażliwością i w reakcjach anafilaktycznych. Immunoglobuliny E (Igl
wyzwalają degranulację bazofilów. Uwalniają się wtedy zmagazynować w
ziarnistościach heparyna i histamina. Znajdująca się w osoczu krw
histamina pochodzi z ziarnistości bazofilów. W czasie degranulacji uwal
niane są z błony komórkowej bazofilów pochodne kwasu arachidonowe
go, przede wszystkim leukotrien C4 (LTC4). W błonie komórkowej bazo
filów wykazano występowanie receptorów dla pięciu interleukin (IL-2
IL-3, IL-4, IL-5, IL-8) oraz dla czynnika wzrostu nerwów (NGF).
Występują podobieństwa pomiędzy czynnością bazofilów i komórek
tucznych. Przypuszcza się nawet, że po przejściu bazofilów z krwi do
tkanek spełniają tam funkcje komórek tucznych.
Limfocyty
Limfocyty krążące we krwi obwodowej są niejednorodną populacją
komórek, zarówno pod względem miejsca ich powstawania, jak i pod
względem pełnionych funkcji. Pochodzą z różnych tkanek i narządów,
w których są wytwarzane: ze szpiku kostnego czerwonego, grasicy, węzłów chłonnych, grudek chłonnych błon śluzowych i ze śledziony. Wymienione narządy wraz z naczyniami chłonnymi i krwionośnymi tworzą
układ limfoidalny (lymphoid system). Limfocyty, krążąc pomiędzy tkankami układu limfoidalnego, dzielą się, dojrzewają i zmieniają swoje
właściwości.
Dzięki występowaniu cząsteczek różnicujących — CD (cluster designations) — można rozpoznawać limfocyty we krwi obwodowej. Limfocyty dzielą się na trzy zasadnicze grupy: limfocyty T — grasiczozależne
(thymus dependent), limfocyty B — szpikozależne (bonę marrow dependent), i limfocyty NK — naturalni niszczyciele (natural killer cells).
Limfocyty T stanowią około 70% wszystkich limfocytów krążących we
krwi, limfocyty B — około 15% i limfocyty NK — około 15%.
Limfocyty T dzielą się na dwie główne podgrupy. Jedną podgrupę
tworzą limfocyty T pomagające (helper T cells), o cząsteczkach różnicujących CD4, których jest około 40%. Drugą podgrupę stanowią
limfocyty T cytotoksyczne (cytotoxic T cells), o cząsteczkach różnicujących CD8, których jest około 30%.
Limfocyty T CD4 pod wpływem swoistych immunogenów lub nieswoistych mitogenów wydzielają polipeptydowe przekaźniki humoralne, czyli
cytokiny, które z kolei aktywują limfocyty B, inne limfocyty T, w tym
również limfocyty T CD8. Do cytokin wydzielanych przez limfocyty CD4
Elementy morfotyczne krwi
313
zalicza się: pięć interleukin (IL-2, IL-4, IL-5, IL-6, IL-10), interferon
gamma (IFN-y) i czynnik martwicy nowotworów (TNF).
Limfocyty T CD8 niszczą komórki zawierające obce antygeny, np.
komórki, w których rozwijają się wirusy, jak również hamują aktywację
limfocytów B wywołaną przez limfocyty T CD4. Prawidłowy stosunek
ilościowy limfocytów T CD4 do limfocytów T CD8 we krwi obwodowej
zapewnia prawidłową reakcję immunologiczną typu komórkowego na
antygeny pojawiające się w organizmie.
Nadmiar limfocytów T CD8 lub niedobór limfocytów T CD4 prowadzi do osłabienia lub niewystąpienia reakcji immunologicznej typu komórkowego na antygen.
Limfocyty B CD19 są odpowiedzialne za humoralny mechanizm
odpowiedzi immunologicznej. Recyrkulujące limfocyty B zatrzymują się
w strefie podtorebkowej węzłów chłonnych, dzielą się i przekształcają
w dojrzałe komórki plazmatyczne intensywnie syntetyzujące immunoglobuliny.
Limfocyty NK CD16 wykazują aktywność cytotoksyczną, niszcząc
komórki, w których rozwijają się wirusy, jak również komórki nowotworowe. Limfocyty NK i limfocyty cytotoksyczne niszczą obce komórki
za pomocą wytwarzanych przez siebie białek. Do tych białek należy
perforyna, która uszkadza błonę komórkową obcych komórek, niszczonych przez limfocyty, tworząc otworki w ich błonach komórkowych.
Wytwarzana przez fagocyty interleukina 12 (IL-12) wzmaga aktywność
limfocytów NK.
Biosynteza immunoglobulin
Limfocyty B powstające w szpiku kostnym mają informację genetyczną
potrzebną do wytwarzania różnorodnych immunoglobulin. W procesie
dojrzewania i różnicowania w narządach limfoidalnych centralnych i obwodowych limfocyt B specjalizuje się w syntezie jednorodnej pod względem chemicznym immunoglobuliny.
Powstawanie immunoglobulin wiążących antygeny przebiega odmiennie w zależności od tego, czy antygen po raz pierwszy występuje w organizmie, czy też pojawia się ponownie. Pierwsze pojawienie się antygenu
wyzwala pierwotną odpowiedź immunologiczną (primary immune response). Antygen po wniknięciu do środowiska wewnętrznego organizmu
zostaje sfagocytowany przez komórkę żerną i dezaktywowany. Po pewnym
czasie nieczynny antygen zostaje uwolniony z komórki żernej i wraz
z chłonką dostaje się do węzłów chłonnych.
314
Krążenie
Limfocyt B
Makrofag
Ryc.
157.
Wytwarzanie
przeciwciał
pod wpływem
antygenu.
Makrofagi
fagocytują
antygen,
degradują w
Limfocyt B
zakfywowany
Immunoglobuliny
IgG IgA IgM
IgD IgE
wakuolach i wydalają. Fragmenty antygenu przyczepione do komórek prezentujących
antygen (APC) pobudzają limfocyty T CD4 do wydzielania interleukin (IL-1, IL-4 i IL-5),
które aktywują limfocyty B. Zaktywowane limfocyty B pod wpływem fragmentów
antygenu przyczepionych do komórek APC transformują w plazmocyty wytwarzające
immunoglobuliny. Swoiste immunoglobuliny stanowiące przeciwciała w stosunku do
antygenu wiążą się z nim i w ten sposób unieczynniają go.
Zdezaktywowany antygen przyczepia się do błony komórkowej makrofagów w węzłach chłonnych. Makrofagi spełniają rolę komórek prezentujących antygen (antigen presenting cells) limfocytom T CD4 pomagającym (helper T cells). Pod wpływem cytokin wytwarzanych przez limfocyty
T CD4 i komórek prezentujących antygen dochodzi do aktywacji limfocytów B i ich przekształcania się w komórki plazmatyczne wytwarzające
w znacznych ilościach specyficzne immunoglobuliny. Krążące w płynach
ustrojowych specyficzne immunoglobuliny wiążą antygen i unieczynniają
go. Ubywanie antygenu krążącego w płynach ustrojowych wywołuje
przewagę działania limfocytów T CD8 — supresorowych (supressor T
cells) — nad limfocytami T CD4. Pod wpływem cytokin wydzielanych
przez limfocyty T CD8 komórki plazmatyczne zmniejszają lub przestają
wydzielać specyficzne immunoglobuliny.
Elementy morfotyczne krwi
315
PIERWOTNA ODPOWIEDŹ IMMUNOLOGICZNA
Nieczynny antygen ■+■ Makrofag
Makrofag
prezentujący antygen (APC)
Limfocyt T CDU ------------------ ». Limfocyt T CDU
nieaktywny
aktyw ny
I
Cytokiny
{
Limfocyt B ---------- +■ Komórka plazmatyczna ------- ►- Immunoglobuliny
IgG IgA IgM
IgD IgE
WTÓRNA ODPOWIEDŹ IMMUNOLOGICZNA
Antygen
Limfocyt B
V
ze swoistymi
immunoglobulinami na
powierzchni
Plazmocytoblast
z antygenem
na powierzchni
Cyłokiny
I
Komórka ________ Immunoglobuliny
*" plazmaIgG
IgA IgM
tyczna
IgD
IgE
Ryc. 158. Pierwotna i wtórna odpowiedź immunologiczna na pojawiający się antygen.
Limfocyty B, przekształcając się w komórki plazmatyczne, wytwarzają
specyficzną immunoglobulinę o właściwościach przeciwciała wiążącego
określony antygen. W różnicujących się limfocytach B zachodzą zmiany
w aparacie genetycznym. Różne fragmenty łańcuchów kwasu deoksyrybonukleinowego (DNA) łączą się ze sobą i w czasie transkrypcji
powstają odmienne łańcuchy matrycowego kwasu rybonukleinowego
(mRNA), na których w procesie translacji wytwarzane są specyficzne
immunoglobuliny.
Wtórna odpowiedź immunologiczna (secondary immune response) zachodzi pod wpływem tego samego antygenu, po jego ponownym pojawieniu się wewnątrz organizmu. Antygen dostaje się do chłonki i zostaje
związany ze swoistymi przeciwciałami znajdującymi się na powierzchni
błony komórkowej limfocytów B. Limfocyty B ze związanym na swojej
powierzchni antygenem dzielą się i przekształcają w duże zasadochłonne
plazmoblasty w strefie podtorebkowej węzłów chłonnych. Plazmoblasty,
316
Krążenie
a przede wszystkim pochodzące od nich dojrzałe komórki płazmatyczne
zaczynają intensywnie wytwarzać swoistą immunoglobułinę.
Immunoglobuliny są syntetyzowane wewnątrz komórki na polirybosomach ziarnistej siateczki śródplazmatycznej. Stąd są transportowane na
zewnątrz komórki.
Limfocyty B krążące we krwi syntetyzują niewielkie ilości immunoglobułin, które są zatrzymywane na zewnętrznej powierzchni ich błony
komórkowej. Natomiast duże, dojrzałe komórki płazmatyczne syntetyzują
znaczne ilości immunoglobulin, które są uwalniane do chłonki i z nią
dostają się do krwi.
Wszystkie immunoglobuliny mają zbliżoną budowę chemiczną, której
podstawową jednostką jest cząsteczka składająca się z czterech łańcuchów
polipeptydowych: o dwóch identycznych lekkich łańcuchach (masa cząsteczkowa od 22 kDa do 23,5 kDa) i również dwóch identycznych ciężkich
łańcuchach (masa cząsteczkowa od 52 kDa do 77 kDa).
Ryc. 159. Immunoglobuliny. Monomer jest
zbudowany z dwóch łańcuchów polipeptydo
wych lekkich i dwóch łańcuchów ciężkich.
Dimer składa się z dwóch podstawowych
jednostek (monomerów), zaś pentamer z pięMonomer O/mer
Pentamer
ciu podstawowych jednostek.
Łańcuchy te są ze sobą połączone dwusiarczkowymi wiązaniami.
Poszczególne immunoglobuliny różnią się między sobą odmienną sekwencją aminokwasów w łańcuchach polipeptydowych oraz występowaniem
w postaci oddzielnych, pojedynczych jednostek o czterech łańcuchach lub
w postaci połączonych ze sobą dwóch i więcej jednostek.
Monocyty
Monocyty (monocytes) krążące we krwi pochodzą u ludzi dorosłych
ze szpiku kostnego czerwonego. Po wyjściu ze szpiku pozostają we krwi
od 8 do 72 godzin. Ponad trzy razy więcej monocytów krwi osadzonych
jest na śródbłonku naczyń krwionośnych, tworząc pulę monocytów
przyściennych, w stosunku do monocytów swobodnie krążących we krwi.
Elementy morfotyczne krwi
31 7
Monocyty po przejściu z krwi do tkanek stają się makrofagami tkankowymi i wykazują zróżnicowaną funkcję w zależności od tkanki, do której przeszły. Będą to pęcherzykowe makrofagi w płucach, komórki
siateczkowo-śródbłonkowe gwiaździste w wątrobie, komórki kościogubne — czyli osteoklasty — w tkance kostnej, makrofagi w jamie
otrzewnej i w torebkach stawowych. Wszystkie te komórki należą do
układu siateczkowo-śródbłonkowego (reticuloendothelial system).
Czynniki, które aktywują makrofagi tkankowe można zaliczyć do
czynników fizjologicznych pochodzących z własnego organizmu i do
czynników patologicznych wytwarzanych przez bakterie, pasożyty,
wirusy i obce substancje chemiczne.
W komórkach śródbłonka naczyniowego, pod wpływem czynników
wytwarzanych w ognisku zapalnym, zachodzi ekspresja cząsteczek adhezyjnych komórek naczyniowych — VCAM-1 (vascular celi adhesion molecule — 1) i cząsteczek adhezyjnych śródbłonkowych leukocytarnych —
ELAM-1 (endothelial leukocyte adhesion molecule — 1). Monocyty
przyczepiają się do komórek śródbłonka, dzięki występowaniu cząsteczek
adhezyjnych, i przechodzą pomiędzy nimi do tkanek. Następnie przyciągane przez liczne czynniki chemotaktyczne wytwarzane w ognisku zapalnym
przemieszczają się w jego kierunku.
Monocyty i makrofagi biorą udział w następujących procesach:
regulacja biosynezy immunoglobulin;
reakcje przeciwbakteryjne przeciwpasożytnicze, przeciwgrzybicze
i przeciwwirusowe;
usuwanie uszkodzonych tkanek;
kierowanie czynnością fibroblastów i komórek tkanki łącznej;
angiogeneza;
wytwarzanie czynników wzrostowych.
Uwalniany przez inne komórki zdezaktywowany antygen wiąże się
z cząsteczką klasy II głównego układu zgodności tkankowej (MHC) na
błonie komórkowej makrofagów. W ten sposób makrofagi stają się
komórkami prezentującymi antygen (antigen-presenting cells) limfocytom T i wyzwalają reakcję komórkową na antygen lub limfocytom B,
inicjując reakcję humoralną na antygen.
Kierowanie czynnością fibroblastów i komórek tkanki łącznej oraz
angiogeneza odbywa się za pośrednictwem licznych przekaźników humoralnych wydzielanych przez makrofagi. Rozmieszczone w różnych tkankach makrofagi mogą łącznie wydzielać ponad sto różnych przekaźników
humoralnych.
318
Krążenie
Przekaźniki humoralne wydzielane przez makrofagi można podzielić
na kilka grup:
interleukiny (IL-1);
czynniki wzrostowe (czynnik wzrostowy pochodzący od trombocytów — PDGF, czynnik wzrostowy transformujący beta — TGF-0);
eikozanoidy (prostaglandyny E — PGE2);
enzymy i czynniki hamujące aktywność enzymów;
czynniki krzepnięcia krwi (czynnik II, V, VII, IX, X);
białka adhezyjne (fibronektyna);
tlenek azotu (NO).
1 Trombocyty
Trombocyty (platelets), czyli płytki krwi, są wytwarzane w szpiku
kostnym czerwonym z megakariocytów. Są one fragmentami cytoplazmy
megakariocytów, która oderwała się od nich w wyniku ich dojrzewania.
W 1 L krwi obwodowej znajduje się średnio 250 xl O 9 płytek krwi
(140 — 440 x 109). Krążą we krwi od 8 do 10 dni, a czas ich połowicznego
krążenia wynosi od 4 do 5 dni. Zatrzymywane są w śledzionie, która jest
zasadniczym miejscem ich rozpadu. Uczestniczą w hemostazie.
W miejscu uszkodzenia śródbłonka naczyniowego i odsłonięcia warstwy podśródbłonkowej naczynia krwionośnego trombocyty przylepiają się
i tworzy się czop. W warstwie podśródbłonkowej występują białka
rozpoznawane przez receptory trombocytów. Do tych białek należą:
kolagen, fibronektyna, trombospondyna, witronektyna, czynnik von Willebranda i 1 amin i na.
Trombocyty, przyczepiając się do białek warstwy podśródbłonkowej,
jednocześnie agregują ze sobą i uwalniają ze swoich ziarnistości cytoplazmatycznych liczne czynniki, które sprzyjają dalszej agregacji.
Do czynników agregujących należą:
•
•
•
•
•
adenozynodifosforan (ADP) i adenozynotrifosforan (ATP);
adrenalina i serotonina;
jony wapniowe;
czynniki krzepnięcia krwi (czynnik I, II, V, XI);
czynniki wzrostowe (czynnik wzrostowy pochodzący z trombocytów —
PDGF, czynnik transformujący beta — TGF-P);
• białka adhezyjne (fibronektyna, trombospondyna).
Osocze
319
Z błony cytoplazmatycznej agregujących trombocytów uwalniane są pochodne kwasu arachidonowego w postaci tromboksanów (TxA 2, TxB2)
i leukotrienów (LTB4, LTC 4, LTD4, LTE4).
I Osocze
Osocze krwi (blood plasma) należy do płynu zewnątrzkomórkowego
i zawiera składniki nieorganiczne i organiczne.
Składniki nieorganiczne osocza dzielą się na:
• kationy — najwięcej jest jonów sodowych i potasowych;
• aniony — najliczniejsze są jony chlorkowe i węglanowe.
Składniki organiczne osocza dzielą się na:
• białka;
• składniki pozabiałkowe z resztą azotową i bez reszty azotowej;
• lipidy.
Zawartość składników nieorganicznych w płynie tkankowym jest
zbliżona do zawartości ich w osoczu. Większe różnice w składzie szybko
wyrównują się. Od prawidłowego składu płynu tkankowego, pośrednio od
składu osocza, zależy pobudliwość komórek, a więc właściwości błony
Tabela 23
Osocze krwi
Składniki
nieorganiczne
Stężenie
Na+
K+
142 mmol/L
4 mmol/L
Ca2 +
2,5 mmol/L
Składniki
organiczne
Białka
Stężenie
Pozabiałkowe:
70—75
4—6
g/L
mmol/L
Glukoza
Kwas mlekowy
Aminokwasy
0,4—1,7 mmol/L
30—55 mg/L
Mg2 +
Cl
1
103
mmol/L
mmol/L
HCO"
26
mmol/L
Amoniak
24—41
1 mmol/L
0,5 mmol/L
Mocznik
Bilirubina
1,3—3,3 mmol/L
1,7—6,8 umol/L
Kwas moczowy
180—380 umol/L
POJ-
so2.-
Kreatynina
umol/L
62—133 umol/L
320
Krążenie
komórkowej i metabolizm wewnątrzkomórkowy. Szczególne znaczeni
ma stosunek jonów sodowych do potasowych.
Zwiększenie stężenia jonów potasowych zaburza prawidłowe stosunk
pomiędzy ich rozmieszczeniem wewnątrz i zewnątrz komórek i moż<
spowodować utratę pobudliwości komórek nerwowych i mięśniowych
a szczególnie komórek mięśnia sercowego.
Równowaga kwasowo-zasadowa
W tkankach organizmu stale wytwarzane są kwasy, takie jak: węglowy, mlekowy, moczowy i inne. W czasie pracy mięśni ilość tworzących
się w organizmie kwasów znacznie zwiększa się.
Trawienie pokarmów związane jest z wydzielaniem soków trawiennych
zarówno silnie kwaśnych, np. sok żołądkowy, jak i zasadowych, np. sok
trzustkowy. Powoduje to ubywanie anionów lub kationów z osocza krwi.
Spożywane pokarmy zawierają również składniki kwaśne i zasadowe,
które wchłaniane są do krwi. Stale usuwany jest z organizmu z wydychanym powietrzem kwas węglowy. Kwasy i zasady nielotne są usuwane
z moczem.
Utrzymanie równowagi kwasowo-zasadowej (acid-base balance) zależy przede wszystkim od właściwości buforowych krwi. Właściwości te są
związane z występowaniem we krwi:
• buforu wodorowęglanowego:
[HCO7]
[H2CO3]
buforu fosforanowego:
[H2PO;J
• białek osocza, których
kwasy, jak i zasady;
• krwinek czerwonych.
cząsteczki wiążą zarówno
W płucach, we krwi tętniczej, aniony Cl" przechodzą przez otoczkę
erytrocytów do osocza, a w tkankach, we krwi żylnej, aniony Cl" powracają do krwinek.
Osocze
321
Wędrówka anionów Cl jest spowodowana właściwościami oksyhemoglobiny, która będąc silniejszym kwasem od hemoglobiny, wypiera aniony
Cl z erytrocytów. Powrót anionów Cl" we krwi żylnej do erytrocytów
sprzyja utrzymaniu stałej wartości pH osocza krwi żylnej.
I Białka osocza
Białka osocza (plasma pr oteins) występują w ilości 70—75 g
w 1 L osocza. Białka osocza dzielą się na trzy zasadnicze frakcje:
• albuminy;
• globuliny;
• fibrynogen.
Frakcje albumin i fibronogenu są stosunkowo jednorodne, natomiast
globuliny dzielą się na liczne grupy. W zależności od zastosowanego
sposobu frakcjonowania uzyskuje się większą lub mniejszą liczbę grup
białek we frakcji globulin.
Rozdzielając białka osocza za pomocą najczęściej stosowanej metody
elektroforetycznej, uzyskuje się:
• albuminy —55,1%
• globuliny — 38,4%
o alphaj — 5,3%
o alpha 2 — 8,7%
o beta
— 13,4%
o gamma — 11,0%
• fibrynogen — 6,5%
(SI: 0,551 kg/kg)
(SI: 0,384 kg/kg)
(SI: 0,053 kg/kg)
(SI: 0,087 kg/kg)
(SI: 0,134 kg/kg)
(SI: 0,110 kg/kg)
(SI: 0,065 kg/kg)
Za pomocą metody ultrawirowania otrzymuje się trzy zasadnicze
frakcje zależne od masy cząsteczkowej białek:
• A — albuminy i małe globuliny o współczynniku sedymentacji (sedimentation coefficient) od 4 do 5 S i masie cząsteczkowej 50 — 90 kDa;
• G — globuliny o współczynniku sedymentacji 7 S i masie cząsteczko
wej około 150 kDa;
• M — makroglobuliny o współczynniku sedymentacji 19 S i masie
cząsteczkowej około 900 kD.
Frakcję A wyprzedzają globuliny o mniejszej cząsteczce od albumin
i mniejszym współczynniku sedymentacji 3,5 S.
11 — Fizjologia człowieka
322
Krążenie
Tabela 24
Białka
osocza
krwi
Frakcja białek
Masa
Zawartość
cząsteczkowa (g/D
(kDa)
Prealbuminy
54,98
0,25—0,45
Albuminy
66,46
37—53
54,0
0,01—0,03
Globuliny
«i
«2
«2 - Pi
P,
r
wiążąca tyroksynę (TBG)
wiążąca kortykosteron (CBG)
białko wiążące
retinol
fibronektyna
20,96
440,00
globulina wiążąca steroidy
transferryna
65,00
hemopeksin
60,00
79,55
składnik HDL
hamuje aktywność
enzymów osocza
wiązanie i transport T4
wiązanie Hb
w osoczu
0,03—0,06 wiązanie retinolu
(wit. A)
zlepia ze sobą
0,25—0,4
komórki
0,003—0,015 wiązanie steroidów
płciowych
transport
jonów
2,3—4,3
Fe3+
0,5—1,15 wiązanie hemu
w osoczu
apolipoproteiny 250,0—514,0
B
immunoglobu150,0
8,0—17,0
liny: IgG
160,0
971,0
175,0
190,0
340,0
transport hormonów tarczycy
ciśnienie onkotyczne
wiązanie i transport T4
0,015—0,02 wiązanie i transport hormonów
kory nadnerczy
1,15—2,1
składnik HDL
apolipoproteiny
28,02
A-l
apolipoproteiny
17,44 0,26—0,66
A-2
alphaj-antytry54,00
1,4—3,2
psyna
wiążąca tyro54,00 0,01—0,03
ksyne
haptoglobiny
86,0—200,0 2,9—11,9
IgA
IgM
IGD
IgE
Fibrynogen
55,7
Czynność,
właściwości
1,0—17,0
0,6—3,7
0,15
0,0003
2,0—1,0
składnik LDL
przeciwciała
przeciwciała
przeciwciała
przeciwciała
przeciwciała
czynnik I krzepnięcia krwi
Osocze
323
Albuminy
Ryc. 160. Rozdzielenie białek surowicy krwi za pomocą elektroforezy bibułowej. Rozdzielone
frakcje surowicy po odpowiednim zabarwieniu dają prążek odpowiadający albuminom
i cztery prążki globulin (a,, a2, P i y). U góry wielkość poszczególnych frakcji oznaczona na
pasku bibuły metodą densytometryczną.
Albuminy
Albuminy wytwarzane są w wątrobie, a ich zasadniczą funkcją jest
wiązanie wody. Dzięki wiązaniu dużej liczby cząsteczek wody wywierają
na ścianki naczyń włosowatych ciśnienie koloidoosmotyczne, czyli onkotyczne (oncotic pressure), wynoszące około 3,3 kPa. Dzięki temu
ciśnieniu woda przefiltrowana przez ścianę naczyń włosowatych do
przestrzeni międzykomórkowych powraca do łożyska krwionośnego.
Na cząsteczkach albumin osadzają się związki drobnocząsteczkowe,
między innymi hormony. Albuminy pełnią funkcję nośnika we krwi dla
tych związków.
Globuliny
We frakcji globulin występują:
• mukoproteiny i glikoproteiny, które są połączeniem białka z węglowodanami;
324
Krążenie
• Iipoproteiny, które są połączeniem białka z lipidami;
• globuliny, które wiążą jony metali (metal binding proteins): transfer
rytyna połączona z żelazem i ceruloplazmina — z miedzią;
• gamma-globuliny, które dzielą się na:
o immunoglobuliny G — IgG,
o immunoglobuliny A — IgA,
o immunoglobuliny M— IgM,
o immunoglobuliny D — IgD,
o immunoglobuliny E — IgE.
Gamma-globuliny wytwarzane są w węzłach chłonnych, natomiast
pozostałe globuliny powstają w wątrobie.
Zasadniczą rolą gamma-globulin jest inaktywacja antygenów, ponieważ w tej frakcji białek zawarte są przeciwciała.
Fibrynogen
Fibrynogen jest białkiem wytwarzanym przez wątrobę, o masie cząsteczkowej 340 kDa i znacznie wydłużonej cząsteczce, o stosunku osiowym
jak 20:1 i o okresie połowicznego rozpadu w osoczu (TV2) wynoszącym
4 dni. Cząsteczka fibrynogenu składa się z dwóch identycznych podjednostek, z których każda zbudowana jest z trzech łańcuchów polipeptydowych:
Aalpha, Bbeta i gamma. Obie podjednostki skierowane są do siebie Nkońcami swoich łańcuchów polipeptydowych. W części środkowej
fibrynogenu obie podjednostki połączone są mostkiem dwusiarczkowym
pomiędzy łańcuchami gamma, natomiast łańcuchy Aalpha i Bbeta mają
wolne końce utworzone przez fibrynopeptydy A i B.
W osoczu stale występują w formie nieaktywnej dwa enzymy: protrombina i plazminogen, które po aktywacji przecinają wiązania peptydowe
w cząsteczce fibrynogenu. Aktywny enzym — trombina — działa swoiście,
odcinając od łańcuchów Aalpha i Bbeta obu podjednostek dwa krótkie
łańcuchy, czyli fibrynopeptydy A i B. Powstające cząsteczki fibryny łączą się ze sobą i powstaje sieć włókien tworzącego się skrzepu krwi (blood
ciot).
Aktywny enzym — plazmina — przecina łańcuchy polipeptydowe
zarówno fibrynogenu, jak i fibryny we wszystkich miejscach występowania wiązania peptydowego pomiędzy aminokwasami argininą i lizyną.
Powstające fragmenty fibrynogenu hamują proces krzepnięcia krwi.
Osocze
325
Składniki organiczne pozabiałkowe
osocza
I
Do składników organicznych pozabiałkowych osocza należą:
•
•
•
•
węglowodany i produkty ich przemiany;
produkty przemiany białkowej;
produkty przemiany hemu;
inne produkty organiczne przemiany wewnątrzkomórkowej.
We krwi występuje stale glukoza w stężeniu od 3,9 do 6,2 mmol/L
osocza. Po spożyciu pokarmów bogatych w węglowodany zawartość
glukozy we krwi zwiększa się. Kwas mlekowy, który jest końcowym
produktem glikolizy w fazie beztlenowej, znajduje się stale we krwi. Stężenie jego mieści się, w zależności od pracy mięśni szkieletowych,
w granicach od 0,4 do 1,7 mmol/L osocza.
Aminokwasy (amino acids) są najważniejszym produktem przemiany
białkowej. Występują w osoczu w ilości od 30 do 55 mg/L. Wchłaniane
są z przewodu pokarmowego oraz uwalniane do krwi w tkankach
w rezultacie preteolizy. We krwi występuje amoniak w stężeniu od 23,6 do
41,3 mmol/L osocza, który powstaje w tkankach jako produkt deaminacji
aminokwasów.
Komórki wątrobowe wychwytują krążący we krwi amoniak, syntetyzują z niego mocznik i wydzielają go do krwi. Prawidłowa zawartość
mocznika wynosi od 1,3 do 3,3 mmol/L osocza.
W osoczu stale znajdują się produkty katabolizmu hemu — bilirubina,
w ilości od 0,7 do 6,8 umol/L osocza, oraz urobilinogen.
Produktami katabolizmu wewnątrzkomórkowego występującymi stale
we krwi są kwas moczowy i kreatynina.
Kwas moczowy (178—386 umol/L osocza) jest jednym z końcowych
produktów katabolizmu kwasu deoksyrybonukleinowego i kwasu rybonukleinowego.
Kreatynina (62—133 umol/L osocza) powstaje w komórkach mięśniowych z kreatyny.
I Lipidy osocza
Zawartość całkowita lipidów w osoczu (plasma lipids) wynosi od 5 do
8 g w 1 L osocza.
326
Krążenie
W skład lipidów osocza wchodzą:
cholesterol (3,9 mmol/L osocza);
fosfolipidy (3 g/L osocza);
triacyloglicerole (1,5 g/L osocza);
witaminy rozpuszczalne w tłuszczach — A, D, E i K (fat-soluble
vitamins);
hormony stereoidowe wydzielane przez korę nadnerczy, jądro i jajnik;
wolne kwasy tłuszczowe — FFA (free fatty acids).
Lipidy we krwi, poza FFA, są związane z białkiem osocza, tworząc
lipoproteiny. W czasie trawienia i wchłaniania tłuszczów z przewodu
pokarmowego cząsteczki triacylogliceroli kondensują się w enterocytach
jelita, trafiają do naczyń chłonnych jelit, tworząc tzw. chylomikrony,
które następnie dostają się do krwi.
Lipoproteiny osocza dzielą się na:
• chylomikrony;
• lipoproteiny o bardzo małej gęstości — VLDL (very Iow density
lipoproteins);
• lipoproteiny o pośredniej gęstości — IDL (intermediate density lipo
proteins);
• lipoproteiny o małej gęstości — LDL (Iow density lipoproteins);
• lipoproteiny o dużej gęstości — HDL (high density lipoproteins) (patrz
tab. 39, str. 460).
Zasadniczą funkcją lipoprotein osocza jest transport cholesterolu,
fosfolipidów, triacylogliceroli, witamin i hormonów steroidowych. Cząsteczki białka są nośnikiem dla tych substancji lipidowych. Enzym — lipaza lipoproteinowa — znajdujący się w ścianach naczyń krwionośnych,
przede wszystkich w obrębie tkanki tłuszczowej i mięśni, hydrolizuje
triacyloglicerole występujące w lipoproteinach osocza. Uwolnione z triacylogliceroli kwasy tłuszczowe wnikają do komórek śródbłonka naczyń
i stąd do innych komórek.
I Hemostaza
Zatrzymanie krwi w łożysku krwionośnym, czyli hemostaza, zależy od:
• właściwości samych naczyń krwionośnych. Błona mięśniowa uszkodzonych naczyń krwionośnych kurczy się i światło naczyń ulega zamknięciu;
Osocze
327
• obecności trombocytów i tworzenia się z nich czopu trombocytarnego;
• czynników osoczowych powodujących zamianę fibrynogenu na fibrynę,
która wraz z czopem trombocytarnym, erytrocytami i leukocytami
tworzy skrzep krwi.
W czasie tworzenia się czopu z trombocytów, w miejscu uszkodzonego
śródbłonka naczyniowego, uwalniają one tromboksan (TxA), który przyspiesza ich agregację i jednocześnie wywołuje silny skurcz naczyń krwionośnych. Uwalniane ze śródbłonka naczyniowego prostacykliny (PGI)
działają przeciwnie — hamują zlepianie się trombocytów i rozkurczają
naczynia krwionośne.
Śródbtonek
Erytrocyty
Miocyty
Agregujące
gładkie
Uszkodzenie ściany
naczynia
trombocyty
Ryc. 161. Tworzenie się czopu płytkowego
w miejscu uszkodzonej ściany naczynia
krwionośnego.
(Reprodukowano
za
zezwoleniem z W. Sawicki Histologia, II
wyd. Wydawnictwo Lekarskie PZWL,
Warszawa 1997.)
Czop
utworzony
z
trombocytów ściśle przylega do
miejsca, w którym uszkodzony jest śródbłonek naczynia krwionośnego i
są odsłonięte białka adhezyjne, ponieważ na powierzchni trombocytów
występują receptory dla tych białek. Trombocyty wiążą się za
pośrednictwem swoich receptorów z białkami adhezyjnymi.
328
Krążenie
Do białek adhezyjnych należą:
kolagen;
czynnik von Willebranda — vWF (von Willebrand factor);
fibrynogen;
fibronektyna;
trom bospondyna;
laminina;
witronektyna.
Białka adhezyjne takie, jak czynnik von Willebranda (vWF) i fibrynoger
występują również w osoczu krwi. Czynnik von Willebranda występując)
w osoczu jest dimerem o masie cząsteczkowej około 600 kDa. Z dimerów
vWF tworzą się multimery o masie cząsteczkowej do około 20 000 kDa.
Czynnik von Willebranda nie tylko zlepia trombocyty, lecz również łączy
ze sobą łańcuchy lekkie z łańcuchami ciężkimi białka stanowiącego
czynnik przeciwhemofilowy (czynnik VIII krzepnięcia krwi). Na skutek
niedoboru czynnika von Willebranda występuje wtórny niedobór aktywnego czynnika przeciwhemofilowego w osoczu krwi.
Krzepnięcie krwi
Istotą procesu krzepnięcia krwi (blood clotting) i powstania skrzepu
jest zamiana hydrofilnego fibrynogenu — czynnika I — na hydrofobną
fibrynę — czynnik la. W procesie tym bierze udział enzym trombina —
czynnik Ha, która we krwi występuje w postaci nieaktywnej, czyli
protrombiny — czynnika II.
Istnieją dwa mechanizmy aktywacji protrombiny:
• wewnątrzpochodny;
• zewnątrzpochodny.
Wewnątrzpochodny mechanizm krzepnięcia krwi (intrinsic clotting mechanism) ujawnia się wtedy, kiedy krew krzepnie na skutek kontaktu
z materiałami lub związkami o ładunku ujemnym. Wewnątrz łożyska
naczyniowego krew zazwyczaj krzepnie w miejscu uszkodzenia śródbłonka naczyniowego. Trombocyty przyczepiają się bezpośrednio do odsłoniętych białek adhezyjnych, głównie kolagenu, po uszkodzeniu śródbłonka naczyniowego. Kolagen, kalikreiny i kininogeny osoczowe aktywują czynnik XII krzepnięcia krwi. Czynnik XII krzepnięcia krwi może
być również zaktywowany przez ściany probówek lub butelek o ładunku
ujemnym, w których znajduje się krew pobrana z żyły.
Osocze
Tabela 25
Czynniki krzepnięcia krwi
Czynnik Masa
cząsteczkowa
kDa
I
340
II
72
Synonim
Synonim angielski
fibrynogen
fibrinogen
protrombina
prothrombin
III
tromboplastyna tkankowa
thromboplastin
IV
wapń (Ca2+)
calcium
proakceleryna Acglobulina czynnik
chwiejny
proaccelerin
Ac-globulin
labile factor
akceleryna (aktywny czynnik V)
accelerin
prokonwertyna
proconvertin
V
330
VI
VII
50
VIII
330
globulina przeciwhemofilowa
AHG czynnik
przeciwhemofilowy AHF
czynnik przeciwhemofilowy A
antihemophilic globulin
antihemophilic factor
antihemophilic factor A
IX
56
czynnik Christmas PTC
czynnik przeciwhemofilowy B
Christmas factor plasma
tromboplastic
component
antihemophilic factor B
X
56
czynnik Stuarta
Stuart-Prower factor
XI
160
czynnik Rosenthala PTA
czynnik przeciwhemofilowy C
Rosenthal factor plasma
thromboplastin
antecedent
antihemophilic factor C
XII
80
czynnik Hagemana
czynnik kontaktowy
Hageman factor
glass factor
XIII
320
czynnik stabilizujący skrzep
— FSF czynnik LL fibrynaza
fibrin stabilizing factor
Laki-Lorand factor
329
330
Krążenie
Czynniki stanowiące kolejne ogniwa w łańcuchu reakcji enzymatycznych doprowadzających do zamiany fibrynogenu w fibrynę występują
w coraz większym stężeniu. Z tego względu proces krzepnięcia krwi
przyjęto przedstawiać według schematu „kaskadowego".
Wewnątrzpochodny mechanizm krzepnięcia krwi rozpoczyna się od
aktywacji czynnika XII. W kolejnych „kaskadach" dochodzi do zamiany
nieaktywnych czynników: XI, IX, X i II na aktywne aż do ostatniej
„kaskady", w której fibrynogen — czynnik I, przekształca się w fibrynę
labilną — czynnik la.
Pod wpływem czynnika XHIa i jonów Ca 2+ fibryna labilna (la)
zamienia się w fibrynę stabilną (Ib).
Zewnątrzpochodny mechanizm krzepnięcia krwi (extrinsic clotting mechanism) występuje po zetknięciu się krwi wypływającej z naczyń krwionośnych z uszkodzonymi tkankami. Tromboplastyna tkankowa wraz
z aktywnym czynnikiem VII, występującym również w tkankach, aktywują
czynnik IX lub czynnik X krzepnięcia krwi. Aktywny czynnik X zwrotnie
przyspiesza aktywację czynnika VII. Poczynając od aktywacji czynnika X,
wewnątrzpochodny i zewnątrzpochodny mechanizm aktywacji protrombiny i krzepnięcia krwi przebiegają jednakowo.
Proces krzepnięcia krwi dzieli się na fazy:
• I faza — obejmuje aktywację wszystkich czynników niezbędnych do
zamiany czynnika II na aktywny czynnik Ha;
• II faza — następuje zamiana protrombiny — czynnika II — na trombinę — czynnik Ila;
• III faza — powstaje fibryna stabilna — czynnik Ib — z fibryno
genu — czynnika I.
Po wytworzeniu się skrzepu dochodzi do jego retrakcji (ciot retraction). Włókienka fibryny skracają się i wyciskają ze skrzepu część płynną
osocza — surowicę krwi (blood serum).
Fibrynogen stabilny podlega proteolitycznemu rozpadowi, czyli fibrynolizie. Enzymem, który wywołuje fibrynolizę, jest plazmina stale
występująca we krwi w postaci nieczynnego proenzymu — plazminogenu.
Przejście proenzymu w aktywny enzym jest zarówno aktywowane, jak
i hamowane przez inne enzymy. Aktywatorami plazminogenu są:
• tkankowy aktywator plazminogenu — t-PA (tissue plasminogen activator);
• urokinazowy aktywator plazminogenu — u-PA (urokinase plasmino
gen activator).
Osocze
331
Ryc. 162. Schemat kaskadowy wewnątrzpochodnego mechanizmu aktywacji protrombiny
i krzepnięcia krwi.
Ryc. 163. Schemat kaskadowy zewnątrzpochodnego mechanizmu aktywacji protrombiny
i krzepnięcia krwi.
332
Krążenie
Aktywację wywołuje przecięcie tylko jednego wiązania peptydowego
w proenzymie pomiędzy aminokwasami argininą i waliną w pozycji
561—562 łańcucha polipeptydowego plazminogenu. Enzymami hamującymi aktywację plazminogenu są dwa enzymy inhibitory aktywacji plazminogenu 1 i 2 — PAI-1, PAI-2 (plazminogen activator inhibitor 1, 2).
Aktywny enzym — plazmina — degraduje fibrynogen do jego fragmentów rozpuszczalnych w wodzie. Proces ten może być również hamowany
przez inny enzym — alpha, antyplazminę.
Wrodzony brak lub niedobór jednego z czynników krzepnięcia krwi
powoduje zwolnienie lub upośledzenie procesu tworzenia się skrzepu.
Wrodzony brak czynnika VIII lub czynnika von Willebranda jest przyczyną hemofilii typu A, brak czynnika IX — hemofilii typu B, i brak
czynnika XI — hemofilii typu C.
Niedobór witaminy K w pokarmach, jej wypieranie przez pochodne
dikumarolu lub jej upośledzone wchłanianie z jelita, hamuje przede
wszystkim syntezę w wątrobie protrombiny — czynnika II, jak również
hamuje syntezę czynników: VII, IX i X oraz białka C i białka S. Krew nie
krzepnie lub krzepnie znacznie wolniej. Stan nabytego niedoboru czynników krzepnięcia nosi nazwę hemofilii rzekomej lub pseudohemofilii.
Krzepnięcie krwi wypełniającej łożysko krwionośne stale jest hamowane przez wiele czynników. Do tych czynników hamujących krzepnięcie
krwi zalicza się takie białka, jak: trombomodulinę, białko C i białko S.
oC2 antyplazmina Ryc. 164.
Schemat fibrynolizy i rozpuszczania się skrzepu.
CMonka
333
Trombomodulina, występująca na powierzchni komórek śródbłonka
naczyniowego, wiąże czynnik Ha, czyli trombinę. Białko C niszczy czynnik
Va i czynnik VIIIa, działając wraz z białkiem S.
We krwi występuje stale heparyna w ilości około 100 ng/L osocza. Jest
ona wytwarzana przez granulocyty zasadochłonne oraz komórki tuczne
występujące w tkance łącznej. Heparyna blokuje aktywność wielu czynników, przede wszystkim działanie trombiny (czynnika Ha) na fibrynogen
(czynnik I) i w ten sposób hamuje krzepnięcie krwi.
Można również nie dopuścić do wystąpienia procesu krzepnięcia krwi
pobranej z żyły, np. konserwując krew przez dodanie słabych kwasów
wiążących jony Ca2+.
I Chłonka
Część osocza przefiltrowana do tkanek przez ściany naczyń włosowatych dostaje się do naczyń chłonnych i powraca w postaci chłonki do krwi
żylnej przez przewód piersiowy {ductus thoracicus) i przewód chłonny
prawy {ductus lymphaticus dexter). Z osocza do tkanek zostaje przefiltrowana woda oraz rozpuszczone w niej składniki drobnocząsteczkowe.
Białka krwi filtrują się do płynu tkankowego w ilości około ułamka
procentu.
Chłonka przepływająca przez naczynia chłonne (vasa lymphatica) nie
ma jednolitego składu. Szczególnie zmienia się zawartość białek. Skład
chłonki zależy od stanu czynnościowego organizmu i narządu, od którego
ona odpływa. W czasie trawienia pokarmów w przewodzie pokarmowym
chłonka odpływająca od narządów jamy brzusznej jest bogata w tłuszcze
wchłonięte w jelicie oraz białka zsyntetyzowane w narządach jamy
brzusznej, przede wszystkim w wątrobie.
W naczyniach chłonnych wątroby występuje największa zawartość
białek. W wątrobie syntetyzowana jest większość białek znajdujących się
we krwi i część z nich — zwłaszcza te, które tworzą duże cząsteczki —
odpływa przez naczynia chłonne.
Odpływ białek wytwarzanych przez komórki lub przefiltrowanych
z osocza krwi zapobiega gromadzeniu się białek w płynie tkankowym
i sprzyja utrzymaniu jego stałego ciśnienia onkotycznego.
Przez naczynia chłonne powracają do krwi nie tylko składniki bezpostaciowe, lecz również recyrkulujące limfocyty. Część recyrkulujących
limfocytów po przejściu z naczyń krwionośnych do tkanek trafia do naczyń chłonnych i tą drogą wraca do krwi.
334
Krążenie
I Układ sercowo-naczyniowy
Układ sercowo-naczyniowy ze względu na jego czynność dzieli się na:
• serce składające się z czterech jam: dwóch przedsionków prawego
i lewego {atrium dextrum et atrium sinistrum) i dwóch komór prawej
i lewej (yentriculus dexter et ventriculus sinister). Przedsionki kurcząc się
pracują tak, jak pompy objętościowe, komory zaś pracują jak pompy
ciśnieniowe, zwłaszcza komora lewa;
• tętnice (arteries) i żyły {yenae) krążenia dużego, które tworzą dwa
zbiorniki: zbiornik tętniczy duży (Ra s — systemie arterial reservoir)
i zbiornik żylny duży (Rvs — systemie venous reservoir);
• tętnice i żyły krążenia małego, czyli płucnego, które tworzą dwa
zbiorniki: zbiornik tętniczy płucny (Rap — pulmonary arterial reservoir)
i zbiornik żylny płucny (Rvp — pulmonary venous reservoir);
• dwie sieci naczyń włosowatych: pierwszą łączącą zbiornik tętniczy duży
(Ra s) ze zbiornikiem żylnym dużym (Rvs) i drugą łączącą w płucach
zbiornik tętniczy płucny (Rap) ze zbiornikiem żylnym płucnym (Rvp).
Posługując się najdalej idącym uproszczeniem, można przedstawić
układ sercowo-naczyniowy jako układ składający się z:
• czterech pomp pracujących szeregowo po dwie, czyli z prawego
przedsionka i prawej komory oraz z lewego przedsionka i lewej ko
mory;
• czterech zbiorników krwi (blood reservoirs): dwóch zbiorników dużych
i dwóch zbiorników płucnych;
• dwóch sieci naczyń włosowatych (capillary nets) łączących zbiorniki
tętnicze ze zbiornikami żylnymi.
Każdy z czterech zbiorników zawiera inną objętość krwi (blood volume) oraz w każdym z nich panuje inne średnie ciśnienie. Mimo tych
różnic pojemność krwi (blood capacity) tłoczona w jednostce czasu, np.
w czasie 1 minuty lub 1 godziny, do zbiornika tętniczego dużego musi być
prawie równa (tylko z niewielkim odchyleniem) pojemności krwi odpływającej ze zbiornika żylnego dużego.
W warunkach fizjologicznych, kiedy oddychanie jest swobodne, pojemność krwi tłoczona do zbiornika tętniczego dużego równa się pojemności
krwi tłoczonej do zbiornika tętniczego płucnego.
W spoczynku pojemność krwi odpływającej ze zbiornika żylnego
dużego równa się pojemności krwi odpływającej ze zbiornika żylnego
płucnego.
Układ sercowo-naczyniowy
335
S.n.d.
Ryc. 165. Schemat układu sercowo-naczyniowego. Vd — komora prawa, Vs — komora
lewa, ad — prawy przedsionek, as — lewy przedsionek, Ras — zbiornik tętniczy duży, Rvs—
zbiornik żylny duży, Rap — zbiornik tętniczy płucny, Rvp — zbiornik żylny płucny, s.n.d. —
sieć naczyń włosowatych krążenia dużego, s.n.p. — sieć naczyń włosowatych krążenia
płucnego.
336
Krążenie
Pojemność krwi krążącej pomiędzy zbiornikami zmienia się w określonych stanach fizjologicznych organizmu. W czasie pracy mięśni i rozszerzania naczyń krwionośnych w mięśniach szkieletowych lub w czasie
trawienia i rozszerzania naczyń krwionośnych w narządach jamy brzusznej
początkowo więcej krwi odpływa z Ra s niż dopływa do Rvs. Po zakończeniu pracy lub po strawieniu pokarmów w pewnym okresie więcej krwi
dopływa do Rvs niż odpływa z Ra s.
Postawa ciała ma również wpływ na objętość krwi wypełniającej
zbiorniki żylne. Po zmianie postawy ze stojącej na leżącą więcej krwi
dopływa do zbiornika żylnego płucnego, niż z niego odpływa i zwiększa się
objętość krwi wypełniającej ten zbiornik. Zmiana postawy z leżącej na
stojącą powoduje zmniejszenie objętości krwi wypełniającej zbiornik żylny
płucny.
Zasadniczym zadaniem serca jest pompowanie krwi ze zbiorników
żylnych do zbiorników tętniczych i utrzymywanie w zbiornikach tętniczych odpowiedniego ciśnienia. Zbiorniki tętnicze gromadzą krew tłoczoną przez serce i dzięki panującemu w nich ciśnieniu zapewniają stały
jej przepływ przez sieć naczyń włosowatych również w czasie rozkurczu
serca.
Zbiorniki żylne, zwłaszcza zbiornik żylny duży, w którym stale znajduje się około 54% krwi krążącej, gromadzą krew niezbędną do wypełnienia jam serca i tworzą jej rezerwę. W chwili zwiększonego zapotrzebowania na tlen transportowany przez krew serce przyspiesza częstotliwość swych skurczów. Zwiększa się pojemność minutowa krwi tłoczonej
przez komory (cardiac output) do zbiorników tętniczych. W tym samym
czasie napływa do serca więcej krwi ze zbiorników żylnych.
I Serce
Z czynnością serca ściśle związane są zjawiska:
elektryczne;
mechaniczne;
akustyczne.
I Czynność bioelektryczna serca — EKG
Rozrusznikiem (pacemaker) dla potencjałów czynnościowych mięśnia
sercowego, które wyprzedzają skurcz, jest tkanka układu przewodzącego
Serce
337
(conduction system). Błona komórkowa komórek tkanki układu przewodzącego odznacza się zdolnością do rytmicznej spontanicznej depolaryzacji (pacemaker potential). Komórki tworzące węzeł zatokowo-przedsionkowy depolaryzują się najszybciej w stosunku do pozostałych komórek
tkanki układu przewodzącego. W związku z tym węzeł zatokowo-przedsionkowy stanowi ośrodek pierwszorzędowy, narzucając swój rytm całemu
sercu (patrz str. 113).
Depolaryzacja z węzła zatokowo-przedsionkowego przenosi się do
węzła przedsionkowo-komorowego (Hisa) za pośrednictwem trzech pęczków międzywęzłowych: przedniego (Bachmana), środkowego (Wenckebacha) i tylnego (Thorela) oraz rozchodzi się na mięsień przedsionków,
obejmując prawy i lewy przedsionek. W strefie granicznej i w samym węźle
przedsionkowo-komorowym zaznacza się największe zwolnienie prędkości przenoszenia się depolaryzacji. Z tego węzła depolaryzacja przewodzona jest do mięśnia komór za pośrednictwem pęczka przedsionkowokomorowego .
Węzeł zatokowo -przedsionkowy
Lewa odnoga pęczka
przedsionkowo ■komorowego
Drogi międzywęzłowe
przednia
środkowa tylna
Węzeł przedsionkowo
- komorowy
Pęczek przedsionkowo - komorowy
Prawa odnoga pęczka
przedsionkowo -komorowego
Ryc. 166. Układ przewodzący serca.
Wiązka przednia
lewej odnogi
Wiązka tylna
lewej odnogi
338
Krążenie
Pęczek przedsionkowo-komorowy stanowi jedyne połączenie pomię-1
dzy mięśniem przedsionków i mięśniem komór, przez które przewodzony I
jest stan czynny. W obrębie komór pęczek dzieli się na dwie odnogi: prawą I
biegnącą do prawej komory i lewą — do lewej komory. Odnoga lewa I
pęczka dzieli się w obrębie lewej komory na dwie wiązki: wiązkę przednią I
i wiązkę tylną. Odnoga prawa pęczka i wiązki odnogi lewej pęczka
przechodzą pod wsierdziem w komórki mięśniowe sercowe przewodzące
(włókna Purkinjego). Depolaryzacja przewodzona przez pęczek przedsion- |
kowo-komorowy zaczyna się szerzyć na komórki mięśnia komorowego
w następującej kolejności: warstwa podwsierdziowa w dolnej 1/3 przegrody
międzykomorowej po stronie lewej, od lewej do prawej warstwy
podwsierdziowej przegrody międzykomorowej, warstwa podwsierdziowa
przegrody międzykomorowej po stronie prawej, podwsierdziowe war stwy przykoniuszkowe prawej i lewej komory, ściana lewej komory, ściana prawej komory do nasierdzia, ściana lewej komory do nasierdzia, części przypodstawne przegrody międzykomorowej oraz prawej i lewej komory. Wędrówka dużej liczby jonów przez błonę komórkową związana
jest z postępującą depolaryzacją komórek mięśnia przedsionków i mięśnia komór.
Zmiany ładunku elektrycznego występujące na powierzchni mięśnia
sercowego mogą być odebrane za pomocą elektrod przystawionych
bezpośrednio do serca lub do powierzchni skóry. Elektrody przystawione
Mięsień
przedsionka
Węzeł zatokowo-przedsionktmy
Mięsień komorowy
Ryc. 167. Potencjały
czynnościowe odbierane
z wnętrza: a) komórki
węzła
zatoko-woprzedsionkowego;
b)
komórki
mięśnia
przedsionka; c) komórki
mięśnia komorowego (0,
1, 2, 3 — fazy
repolaryzacji).
Serce
339
Prędkość przenoszenia się depolaryzacji w układzie przewodzącym i w mięśniu sercowym
Komórki
Prędkość
przenoszenia
(m/s)
Węzła zatokowo-przedsionkowego
Mięśnia przedsionków
Strefy granicznej pomiędzy mięśniem
przedsionków i węzłem przedsionkowo-komorowym
0,05
1,0
0,05
Węzła przedsionkowo-komorowego
0,1
Pęczka przedsionkowo-komorowego
2,0
Przewodzące w mięśniu komór
Mięśnia komór pod wsierdziem
Mięśnia komór pod osierdziem
4,0
1,0
0,4
do powierzchni skóry odbierają potencjał czynnościowy mięśnia sercowego o wielokrotnie niższej amplitudzie niż elektrody przystawione bezpośrednio do serca. Ze względów praktycznych stosuje się w medycynie odbiór potencjałów czynnościowych mięśnia sercowego za pomocą elektrod przystawianych do powierzchni skóry.
W celu ujednolicenia metody pomiaru i oceny potencjałów czynnościowych mięśnia sercowego zostały ustalone miejsca na powierzchni ciała,
do których przystawia się elektrody.
Odprowadzenia EKG
Umieszczając elektrody na kończynach i rejestrując różnice potencjałów występujące pomiędzy elektrodami, odbiera się czynność bioelektryczną mięśnia sercowego, czyli elektrokardiogram — EKG, za pomocą tzw.
odprowadzeń kończynowych.
Trzy odprowadzenia kończynowe dwubiegunowe, klasyczne Einthovena, uzyskuje się rejestrując różnice potencjałów pomiędzy:
• prawym przedramieniem (R) i lewym przedramieniem (L) — I odprowadzenie kończynowe;
340
Krążenie
• prawym przedramieniem (R) i lewą golenią (F) — II odprowadzeni
kończynowe;
• lewym przedramieniem (L) i lewą golenią (F) — III odprowadzeni!
kończynowe.
Elektrody przystawione do kończyn zachowują pewne stałe przestrzenne położenie w stosunku do mięśnia sercowego. Elektrody przystawione
do kończyn, które spełniają tylko funkcję przewodników elektrycznych,
odbierają z różnych stron czynność bioelektryczną serca. W związku
Cechowanie
Ryc. 168. Odprowadzenie kończynowe dwubiegunowe standardowe stosowane w elektrokardiografii. G — galwanometr pisakowy. Załamki EKG: P, Q, R, S, T.
Serce
341
Ryc. 169. Trójkąt Einthovena, oś elektryczna serca (x — y) i pole elektryczne wokół serca
w czasie szerzenia się depolaryzacji. I — odprowadzenie pierwsze kończynowe, II — odprowadzenie drugie kończynowe, III — odprowadzenie trzecie kończynowe, R — elektroda
na prawym przedramieniu, L — elektroda na lewym przedramieniu, F — elektroda na lewej
goleni.
z tym każda z elektrod ustawiona jest pod innym kątem względem wektora
siły elektromotorycznej serca (cardiac vector of the electromotive force).
Szerzenie się w mięśniu przedsionków, a następnie w mięśniu komór depolaryzacji, za którą postępuje repolaryzacja, powoduje stałą zmianę
chwilowego wektora siły elektromotorycznej serca. Największy wektor
siły elektromotorycznej serca jest skierowany przestrzennie wewnątrz
klatki piersiowej od okolicy prawej pachy do lewego łuku żebrowego.
Zapis EKG zarejestrowany z każdego z trzech odprowadzeń kończynowych jest inny, ponieważ stanowi wypadkową potencjałów elektrycznych serca występujących w przestrzeni pomiędzy tymi elektrodami.
Rzutując na przednią ścianę klatki piersiowej wektor siły elektromotorycznej serca, można sprowadzić potencjały czynnościowe serca występujące
w przestrzeni do procesu przebiegającego na płaszczyźnie. Łącząc ze sobą
na płaszczyźnie punkty przystawiania elektrod, otrzymuje się trójkąt,
w którego środku znajduje się rzut serca na płaszczyznę czołową.
342
Krążenie
Wewnątrz rzutu serca znajduje się rzut wektora siły elektromotoryczne
serca. Każda z trzech elektrod kończynowych jest ustawiona pod innyn
kątem w stosunku do rzutu wektora siły elektromotorycznej serca, czyli dc
osi elektrycznej serca (cardiac axis). W związku z tym załamki EKG
zarejestrowane z poszczególnych odprowadzeń mają inną amplitudę,
a nawet mogą być inaczej skierowane w stosunku do linii izoelektrycznej.
Załamki EKG są dodatnie, jeśli są skierowane ponad linię izoelektryczną,
oraz ujemne, jeśli są skierowane w dół od tej linii.
Do oceny EKG dogodniejsze są odprowadzenia jednobiegunowe. Za ich
pomocą rejestruje się różnice potencjałów pomiędzy elektrodą aktywną
i elektrodą nieaktywną. Jednobiegunowe odprowadzenie przedsercowe
otrzymuje się po przystawieniu elektrody aktywnej do klatki piersiowej
i rejestracji różnic potencjałów pomiędzy tą elektrodą i elektrodą nieaktywną. Zapis EKG odebrany za pomocą odprowadzeń przedsercowych różni
się od EKG odebranego za pomocą odprowadzeń kończynowych. Elektrody przedsercowe są inaczej ustawione przestrzennie w stosunku do wektora siły elektromotorycznej serca.
Stosuje się sześć jednobiegunowych odprowadzeń przedsercowych
(unipolar chest leads), czyli odprowadzeń przedsercowych Wilsona, ozna-
Ryc. 170. Miejsca przystawienia elektrod
na klatce piersiowej przy jednobiegunowych odprowadzeniach przedsercowych
Wilsona (V,—V6).
Serce
343
czonych: V15 V2, V3, V4, V5 i V6. Elektroda I przedsercowego odprowadzenia V\ jest przystawiona do skóry w czwartym międzyżebrzu po stronie prawej mostka. Pozostałe elektrody V2— V6 są przystawiane do skóry
po stronie lewej klatki piersiowej.
Trzeci rodzaj odprowadzeń stanowią odprowadzenia kończynowe jednobiegunowe. Zwierając bezpośrednio dwie elektrody kończynowe, uzyskuje
się z trzeciej elektrody aktywnej EKG o załamkach o wyższej amplitudzie.
Otrzymuje się w ten sposób trzy odprowadzenia EKG kończynowe,
jednobiegunowe nasilone (augmented unipolar limb leads), czyli odprowadzenia Goldbergera:
• aVR — elektroda aktywna na prawym przedramieniu;
• aVL — elektroda aktywna na lewym przedramieniu;
• aVF — elektroda aktywna na lewej goleni.
Centralna
końcówka
Wilsona
Ryc. 171. Schemat połączeń pomiędzy elektrodami nieaktywnymi
przymocowanymi do kończyn za
pośrednictwem oporu 5000 omów
i elektrodą czynną stosowaną w odprowadzeniach przedsercowych Wilsona. G — galwanometr pisakowy
elektrokardiografu, R — prawe
przedramię, L — lewe przedramię,
F — lewa goleń.
344
Krążenie
Porównywalność zapisów EKG wymaga nie tylko stosowania typowych odprowadzeń, lecz również jednakowych warunków rejestracji
Aparaty — elektrokardiografy — rejestrują krzywą EKG przy podstawie
czasu 2 sekundy, prędkości przesuwu papieru 25 lub 50 mm/s i wzmocnieniu odpowiadającemu 1 mV= 10 mm.
Elektrokardiografii
Elektrokardiogram (EKG) zarejestrowany za pomocą II odprowadzenia kończynowego dwubiegunowego ma pięć załamków oznaczonych
kolejno literami: P, Q, R, S i T. Załamek P odpowiada początkowi
depolaryzacji mięśnia przedsionków. Załamki Q, R i S występują obok
siebie, tworząc zespół QRS odpowiadający początkowi depolaryzacji
mięśnia komór, czyli fazie 0 potencjału czynnościowego komórek mięśnia
komór (patrz Mięsień sercowy, str. 110). Załamki T wiąże się z szybką
repolaryzacją mięśnia komór, co odpowiada fazie 3 potencjału czynnościowego komórek mięśnia sercowego.
Analiza krzywej EKG obejmuje:
• załamki — kierunek ich wychylenia ku górze lub ku dołowi od linii
izoelektrycznej, ich amplitudę, czas trwania, częstotliwość występowa
nia i kształt;
• odcinki — czas trwania linii izoelektrycznej pomiędzy załamkami;
• odstępy — obejmujące łączny czas trwania załamków i odcinków.
Czas trwania poszczególnych załamków, odcinków i odstępów zależy
od czasu trwania potencjałów czynnościowych mięśnia przedsionków
i komór.
Amplituda załamków zależy od wielu czynników. U ludzi dorosłych
jest przede wszystkim wykładnikiem liczby synchronicznie depolaryzujących i repolaryzujących się komórek mięśniowych. Amplituda załamków
zależy od oporu elektrycznego tkanek znajdujących się pomiędzy elektrodą i sercem, od budowy ciała i położenia serca w klatce piersiowej.
Amplituda poszczególnych załamków w każdym odprowadzeniu jest
inna. Mierzona w II odprowadzeniu wynosi dla:
• załamka P 1—2,5 mm = 0,1—0,25 mV;
• zespołu QRS 7—18 mm = 0,7—1,8 mV;
• załamka T 2,5—6 mm = 0,25—0,6 mV.
Serce
345
Tabela 27
Średni czas trwania poszczególnych załamków, odcinków i odstępów w EKG
czas przewodzenia depolaryzacji w mięśniu przedZatamek
P
sionków
100 ms
Odcinek
PQ
czas przejścia depolaryzacji przez węzeł przedsionkowo-komorowy i pęczek przedsionkowo-komorowy
50 ms
Odstęp
PQ
czas przewodzenia depolaryzacji do węzła zatokowo-przedsionkowego do mięśnia komór
150 ms
Zespół
QRS
czas szerzenia się depolaryzacji w mięśniu komór
90 ms
Odcinek
ST
120 ms
Załamek
T
okres depolaryzacji mięśnia komór
czas szybkiej repolaryzacji mięśnia komór (faza 3
repolaryzacji)
Odstęp
ST
280 ms
Odstęp
QT
czas wolnej i szybkiej repolaryzacji mięśnia komór
(2 i 3 faza repolaryzacji)
potencjał czynnościowy mięśnia komór (depolaryzacja i repolaryzacja)
Odstęp
RR
czas trwania jednego cyklu pracy serca
800 ms
120 ms
370 ms
Ryc. 172. Typowa krzywa elektrokardiograficzna zarejestrowana w II odprowadzeniu
kończynowym.
346
Krążenie
Ryc. 173. Prawidłowe zapisy elektrokardiograficzne zarejestrowane w czasie stosowania:
I—III — odprowadzeń kończynowych dwubiegunowych, V,—V6 odprowadzeń przedsercowych jednobiegunowych, aVR, aVL i aVF — odprowadzeń kończynowych jednobiegunowych, nasilonych (aVR — elektroda aktywna na prawym przedramieniu, aVL —
elektroda aktywna na lewym przedramieniu, aVF — elektroda aktywna na lewej goleni). Oś
elektryczna serca u badanego jest nachylona pod kątem +45°.
Serce
347
Nieprawidłowy kształt i amplituda załamków oraz nieprawidłowe
wartości odnoszące się do odcinków i odstępów wskazują na zaburzenia
w czynności bioelektrycznej mięśnia sercowego. Wydłużenie odstępu PQ
powyżej 200 ms świadczy o zwolnieniu przewodzenia stanu czynnego
przez układ przewodzący, zwłaszcza przez węzeł przedsionkowo-komorowy. Brak załamka S, uniesienie odcinka ST ponad linię izoelektryczną
lub inne zniekształcenia zespołu QRST mogą wskazywać na zaburzenia
w krążeniu krwi w naczyniach wieńcowych serca na skutek np. zaczopowania odgałęzienia jednej z tętnic wieńcowych.
Oś elektryczna serca
W zależności od ustawienia serca w klatce piersiowej zmienia się wektor siły elektromotorycznej serca, jak również jego rzut na płaszczyznę
czołową, czyli oś elektryczną serca — AQRS (cardiac axis). Na podsta-
Odchylenie
osi w lewo
Odprowa
dzeni el
+9- 2=+7
+ 180
Ustawienie
osi poziome
Ustawienie osi
pośrednie
Odchylenie osi
w prawo
Ustawienie osi pionowe
Ryc. 174. Wyznaczanie osi elektrycznej serca (AQRS) na podstawie odprowadzeń koń czynowych dwubiegunowych I i III. Prawidłowe ustawienie osi serca: poziome (od +30
do -30°), pośrednie (od +30° do +60°) i pionowe (od +60° do +110°). I — pierwsze
odprowadzenie kończynowe, III — trzecie odprowadzenie kończynowe.
348
Krążenie
wie pomiarów załamków Q, R i S w I i II odprowadzeniu EKG można
wyznaczyć oś elektryczną serca.
W warunkach prawidłowych oś elektryczna serca jest skierowana
podobnie jak bok trójkąta Einthovena wyznaczonego przez II odprowadzenie. Prawidłowe nachylenie osi elektrycznej serca w stosunku do skali
w stopniach przedstawionej na rycinie 174 wynosi od —30° w lewo i do
+ 110° w prawo. Oś elektryczna serca jest zorientowana: poziomo
w zakresie od —30° do +30°, pośrednio w zakresie od +30° do +60°
i pionowo w zakresie od +60° do +110°. Odchylenie w lewo osi
elektrycznej serca, poza wartości fizjologiczne, występuje wtedy, kiedy
kąt jest powyżej —30°. Odchylenie w prawo zaś występuje przy kącie
większym niż +110°.
I Czynność mechaniczna serca
Za falą depolaryzacji szerzy się skurcz mięśnia przedsionków i mięśnia
komór, repolaryzacja zaś wyprzedza ich rozkurcz. Skurcz i rozkurcz
przedsionków i komór powtarzają się cyklicznie, w spoczynku w rytmie
1,2 Hz (72 na minutę). Jeden cykl trwa około 800 ms.
Skurczowi i rozkurczowi mięśnia sercowego towarzyszy napełnianie
się i opróżnianie z krwi jam serca. Jeden kierunek przepływu krwi —
od zbiorników żylnych do zbiorników tętniczych — wiąże się z samą
budową jam serca i istnieniem zastawek, które uniemożliwiają cofanie się
krwi.
Krew szybko napływa z obu zbiorników żylnych do przedsionków
i przez otwarte ujścia przedsionkowo-komorowe do komór w fazie
rozkurczu w okresie szybkiego wypełniania się komór krwią.
Pierwszy kurczy się mięsień przedsionków (atrial systole) i krew
wypełniająca jamy przedsionków zostaje wtłoczona do komór przez
otwarte ujście przedsionkowo-komorowe prawe i lewe {ostium atrioventriculare dextrum et sinistrum). Następnie skurczem izowolumetrycznym
(isovolumetric ventricular contraction) rozpoczyna się skurcz komór
(ventricular systole). Zastawki przedsionkowo-komorowe: prawa — trójdzielna i lewa — dwudzielna (valvae atrioventriculares: dextra — tricuspidalis et sinistra — mitralis) zamykają się, napięcie mięśnia komór szybko
zwiększa się czemu towarzyszy szybki wzrost ciśnienia w obu komorach
(tab. 28 i tab. 29, str. 351).
W momencie kiedy ciśnienie krwi w komorach przewyższy ciśnienie
w zbiornikach tętniczych, zastawka pnia płucnego (yalva trunci pulmona-
Serce
349
Tabela 28
Cykl pracy serca
zy cyklu pracy serca
Fa
Rozkurcz
komór
Skurcz
komór
Rozkurcz
komór
Czas
trwania
(ms)
okres protodiastoliczny
izowolumetryczny
okres szybkiego wypełniania
się komór
przerwa
skurcz przedsionków
izowolumetryczny
izotookres maksymalniczny nego wyrzutu
okres zredukowanego wyrzutu
okres protodiastoliczny
izowolumetryczny
okres szybkiego wypełniania
się komór
przerwa
skurcz przedsionków
Łączny czas trwania skurczu i rozkurczu
komór
Zastawki
przedsionkowo-komorowe
pnia płucnego i
aorty
zamknięte
zamknięte
otwarte
50
90
zamknięte
otwarte
130
zamknięte
40
80
110
otwarte
190
110
800
lis) i zastawka aorty (valva aortae) otwierają się i krew jest wtłaczana do
zbiorników tętniczych. Otwieranie się zastawek pnia płucnego i aorty
rozpoczyna skurcz izotoniczny komór (isotonic ventricular contraction),
w czasie którego krew jest wtłaczana do zbiorników tętniczych (venticular
ejection).
Faza rozkurczu komór (ventricular diastole) rozpoczyna się z chwilą
wystąpienia krótkiego okresu protodiastolicznego (protodiastole), w którym ciśnienie w komorach zaczyna opadać. Potem występuje rozkurcz
izowolumetryczny komór (isovolumetric ventricular relaxation), w czasie
którego zarówno zastawki pnia płucnego i aorty, jak i przedsionkowo-komorowe są zamknięte.
Okres szybkiego wypełniania się komór (rapid inflow) rozpoczyna się
od otworzenia się zastawek przedsionkowo-komorowych i przepływu do
350
Krążenie
Ryc. 175. Zjawiska elektryczne i mechaniczne zachodzące w sercu w czasie dwóch cykli. 1 —
elektrokardiogram, 2 — ciśnienie krwi w lewej komorze, 3 — ciśnienie krwi w aorcie, 4 —
ciśnienie krwi w lewym przedsionku, 5 — ciśnienie krwi w prawej komorze, 6 — ciśnienie krwi
w pniu płucnym, 7 — ciśnienie krwi w prawym przedsionku, 8 — fonokardiogram: I, II i III
ton serca.
Serce
351
Tabela 29
Ciśnienie w jamach serca i w zbiornikach tętniczych
Miejsce pomiaru
Przedsionek prawy
Komora prawa Zbiornik
tętniczy płucny
Przedsionek lewy
Komora lewa Zbiornik
tętniczy duży
Ciśnienie
rozkurczowe
skurczowe
SI: kPa
mm Hg
SI kPa
mm Hg
00
00
0,8
6
0,9
7
3,3
25
3,3
25
00
9,3
00
70
0,9
16,0
16,0
7
120
120
komór krwi nagromadzonej w przedsionkach. Następnie występuje okres
przerwy (diastasis), po którym kurczy się mięsień przedsionków i druga
porcja krwi wypełniającej jamy przedsionków zostaje wtłoczona pod niewielkim ciśnieniem do komór. Kolejny cykl pracy serca rozpoczyna się
od skurczu izowolumetrycznego komór.
Przyspieszenie częstości skurczów serca skraca czas trwania poszczególnych cykli. W większym stopniu ulega skróceniu czas rozkurczu,
w mniejszym zaś czas skurczu. Bez względu na częstotliwość skurczów
serca czas trwania skurczu izowolumetrycznego komór zmienia się tylko
nieznacznie.
Objętości krwi tłoczonej przez prawą i lewą komorę do zbiorników
tętniczych w czasie pojedynczego cyklu pracy serca są sobie prawie równe,
natomiast ciśnienie skurczowe w lewej komorze jest pięciokrotnie wyższe
od ciśnienia w prawej komorze.
Wyrzut serca
Objętość wyrzutowa serca — SV (stroke volume) — jest to ilość krwi
wtłaczanej przez jedną z komór serca do odpowiedniego zbiornika
tętniczego. U dorosłego mężczyzny o masie ciała 70 kg, w spoczynku,
w pozycji leżącej, każda z komór tłoczy około 75 mL krwi w czasie
jednego cyklu pracy serca. W końcu skurczu pozostaje w każdej komorze
około 50 mL krwi stanowiącej objętość krwi zalegającej (residual blood
volume), która warunkuje objętość późnoskurczową komór (end-systolic
ventricular volume).
Frakcja wyrzutowa lewej komory serca określa stosunkiem objętości
wyrzutowej (75 mL) do objętości poźnorozkurczowej (75 mL + 50 mL =
352
Krążenie
= 125 mL). Frakcja ta stanowi dobry wskaźnik stanu czynnościowego
mięśnia lewej komory serca.
Pojemność minutowa serca (cardiac output) jest to ilość krwi tłoczonej przez jedną z komór serca w czasie jednej minuty i wynosi w spoczynku około 90 mL/s (5,4 L/min) krwi. Pojemność minutowa serca
w spoczynku jest przeliczana na 1 m2 powierzchni ciała jako tzw. wskaźnik sercowy (cardiac index). Pojemność minutowa komory wynosi
53 mL/s/m2 (3,2 L/min/m2) powierzchni ciała.
W różnych stanach fizjologicznych pojemność minutowa serca zwiększa się dzięki:
• zwiększeniu objętość' wyrzutowej serca — SV;
• przyspieszeniu czystości skurczów serca.
Objętość wyrzutowa serca zależy od siły skurczu mięśnia komór, która
z kolei jest uwarunkowana: początkowym rozciągnięciem komórek mięśniowych (objętością krwi zalegającej), ciśnieniem panującym w zbiornikach tętniczych i transmitterami układu autonomicznego uwalnianymi
z zakończeń nerwowych w mięśniu sercowym. W czasie intensywnego
wysiłku fizycznego pojemność minutowa serca wzrasta kilkakrotnie
w stosunku do pojemności minutowej w czasie spoczynku.
Zgodnie z prawem Starlinga, które mówi, że energia skurczu jest
proporcjonalna do początkowej długości komórek mięśnia sercowego,
objętość wyrzutowa serca zależy od stopnia wypełniania komór krwią
w końcu rozkurczu (end-diastolic volume). Wypełnianie się jam serca
zależy od warunków przepływu krwi w zbiorniku żylnym dużym. Zwiększenie objętości krwi krążącej (TBV — total blood volume), obniżanie się
ujemnego ciśnienia wewnątrz klatki piersiowej (negative intrathoracic pressure) w czasie pogłębiania oddechów, praca mięśni szkieletowych uciskających żyły są to czynniki zwiększające dopływ krwi do prawego
przedsionka. Zmniejsza się zaś dopływ po zmniejszeniu się objętości krwi
krążącej, spłyceniu oddechów i w czasie bezruchu.
Wzrost ciśnienia w zbiorniku tętniczym dużym na skutek zwiększenia
całkowitego obwodowego oporu naczyniowego (TPR — total peripheral
vessels resistance) początkowo zmniejsza objętość wyrzutową i zwiększa
objętość krwi zalegającej w komorach. Zwiększająca się objętość krwi
zalegającej w komorach w końcu skurczu powoduje większe rozciąganie
komórek mięśniowych i w wyniku tego zwiększenie objętości wyrzutowej
i pojemności minutowej serca. Zmniejszenie obwodowego oporu naczyniowego chwilowo zwiększa objętość wyrzutową, a następnie zmniejsza
objętość wyrzutową i pojemność minutową serca.
Serce
353
Ryc. 176. Przepływ krwi przez poszczególne narządy w spoczynku wyrażony w procentach
w stosunku do pojemności minutowej prawej i lewej komory serca.
Impulsacja biegnąca przez nerwy współczulne i uwalniana na ich
zakończeniach noradrenalina zwiększają siłę skurczów komórek mięśniowych komór bez początkowej zmiany ich długości. Zwiększa się objętość
wyrzutowa serca kosztem zmniejszenia się objętości krwi zalegającej
w komorach w końcu skurczu i zwiększa się pojemność minutowa.
12 — Fizjologia człowieka
354
Krążenie
Czynniki nerwowe i humoralne wpływają na mięsień sercowy i zmie
niają:
•
•
•
•
siłę jego skurczów, czyli mają działanie inotropowe;
częstotliwość skurczów, czyli mają działanie chronotropowe;
przewodzenia stanu czynnego, czyli mają działanie dromotropowe;
pobudliwość, czyli mają działanie batmotropowe.
Noradrenałina uwalniana z zakończeń nerwów współczulnych ma
działanie dodatnie inotropowe, chronotropowe, dromotropowe i batmotropowe, zwiększając objętość wyrzutową i pojemność minutową serca.
Działa ona w ten sposób na mięsień sercowy za pośrednictwem beta-receptora adrenergicznego i cyklazy adenylanowej.
Działanie przeciwne w stosunku do noradrenaliny ma acetylocholina
uwalniana z zakończeń neuronów przywspółczulnych. Wynikiem jej wpływu na serce jest zmniejszenie objętości wyrzutowej i pojemności minutowej serca.
Ruchy serca
W czasie skurczów serce wykonuje niewielki ruch obrotowy wokół swej
osi podłużnej. Jednocześnie pierścienie włókniste ujść przedsionko-wokomorowych obniżają się w czasie skurczów komór w kierunku
koniuszka serca. Oba przedsionki zostają rozciągnięte, co sprzyja ich
wypełnianiu się krwią napływającą ze zbiorników żylnych.
Ruch obrotowy serca wokół osi podłużnej i zwiększenie napięcia
mięśnia komór w czasie skurczu są przyczyną występowania w piątym
międzyżebrzu po stronie lewej tzw. uderzenia koniuszkowego serca.
Echokardiografia
Do badania echokardiograficznego, czyli ultrasonograficznego serca
(USG), wykorzystane są właściwości fal ultradźwiękowych (od 1 do
10 MHz), które przenikając przez tkanki, odbijają się od powierzchni
odgraniczającej dwie tkanki o różnej gęstości akustycznej. Wiązka fal
ultradźwiękowych generowana jest przez kryształ piezoelektryczny znajdujący się w przetworniku przystawionym do skóry na klatce piersiowej
nad sercem. Odbite fale ultradźwiękowe powracają do przetwornika
z różnym opóźnieniem w zależności od głębokości powierzchni je odbijającej.
Serce
355
Rozkurcz Skurcz Rozkurcz Skurcz
Ryc. 177. Zasada uzyskania echokardiogramu. Po lewej schemat przekroju serca w rozkurczu w klatce piersiowej. Po prawej schemat echokardiogramu uzyskanego w czasie dwóch
cykli pracy serca. Wiązka fal ultradźwiękowych kolejno odbija się od: powierzchni
zewnętrznej (1) i wewnętrznej (2) ściany klatki piersiowej, powierzchni zewnętrznej (3)
i wewnętrznej (4) ściany przedniej komory prawej, przegrody międzykomorowej (5),
przedniego (6) i tylnego (7) płatka zastawki dwudzielnej, powierzchni wewnętrznej (8)
i zewnętrznej (9) ściany tylnej komory lewej. D — rozwieranie się płatków zastawki
dwudzielnej, E—Et — maksymalne rozwarcie płatków zastawki dwudzielnej, A — rozwarcie
płatków zastawki dwudzielnej w czasie skurczu przedsionka lewego, C — zamknięcie
zastawki dwudzielnej. EKG — odprowadzenie II kończynowe.
Fale ultradźwiękowe odbijają się od powierzchni zewnętrznej i wewnętrznej ściany klatki piersiowej, od ścian komór i przedsionków serca oraz
od powierzchni zastawek, ponieważ krew wypełniająca jamy serca słabo
odbija fale ultradźwiękowe. Dzięki temu można zobrazować na ekranie
oscyloskopu sygnał elektryczny przetworzonych fal ultradźwiękowych
odbitych od struktur serca położonych w różnej odległości od kryształu
piezoelektrycznego. Zmieniając ustawienie przetwornika i jego kąt nachylenia względem powierzchni klatki piersiowej, uzyskuje się na ekranie
oscyloskopu obraz odbicia fal ultradźwiękowych od różnych struktur
serca.
356
Krążenie
Stosuje się kilka standardowych sposobów badania echokardiograficznego czynności mechanicznej serca. Podczas zastosowania tzw. sposobu M uzyskuje się na ekranie oscyloskopu na osi pionowej obraz odbicia
fal ultradźwiękowych od powierzchni, których odległość zmienia się
w czasie skurczu i rozkurczu. Zmiany w odbitym obrazie, zachodzące
w czasie, przesuwają się na ekranie oscyloskopu wzdłuż osi poziomej.
Przetwornik musi być przystawiony do skóry klatki piersiowej w czwartym międzyżebrzu, przy mostku, po stronie lewej. W sposobie M na
ekranie oscyloskopu uzyskuje się przesuwające się odbicie fal ultradźwiękowych od mięśnia przedniej ściany prawej komory serca, przegrody i mięśnia tylnej ściany lewej komory serca, jak również od przedniego
i tylnego płatka zastawki dwudzielnej. W czasie skurczu serca poszerza się
odbicie fal od powierzchni kurczącego się mięśnia sercowego. Płatki
zastawki dwudzielnej są zamknięte, dając obraz pojedynczego odbicia fal
ultradźwiękowych. W czasie rozkurczu zwężają się odbicia fal ultradźwiękowych od ścian komór, od otwartej zastawki dwudzielnej uzyskuje
się zaś dwa odbicia od obu płatków — przedniego i tylnego.
Korelacja zachodzących w czasie procesów elektrycznych zarejestrowanych jako elektrokardiogram (EKG) z procesami mechanicznymi,
w postaci echokardiogramu uzyskanego sposobem M, pozwala na pomiar:
czasu skurczu i rozkurczu mięśnia komór, szerokości światła prawej i lewej komory serca, grubości ściany przedniej prawej komory, grubości
przegrody międzykomorowej i grubości tylnej ściany lewej komory.
Umożliwia również pomiar rozwarcia płatków zastawki dwudzielnej.
I Zjawiska akustyczne
Drgania towarzyszące pracy serca są niejednorodnej częstotliwości.
Z punktu widzenia praktyki lekarskiej dzieli się je na stale występujące
w warunkach fizjologicznych, czyli tony serca (heart sounds), oraz na
patologiczne szmery.
W każdym cyklu pracy serca występują tony:
• pierwszy ton serca — wywołany zamykaniem się zastawek przedsionkowo-komorowych i początkiem skurczu serca. Trwa około 150 ms
i obejmuje drgania o częstotliwości od 25 do 45 Hz;
• drugi ton serca — powstaje w czasie zamykania się zastawek aorty i pnia
płucnego. Trwa krócej od pierwszego tonu — 120 ms — i ma większą
częstotliwość — około 50 Hz;
Serce
357
Ryc. 178. Fonokardiogram I, II i III tonu serca oraz mechanizm powstawania poszczególnych drgań. PP — przedsionek prawy, KP — komora prawa, PL — przedsionek lewy,
KL — komora lewa.
• trzeci ton serca — występuje w rozkurczu, w okresie wypełniania się
komór krwią napływającą z przedsionków. Jest najsłabszy. Spowodowany jest przez wibrację krwi szybko wypełniającej jamy obu komór.
W warunkach patologicznych występują dodatkowo w czasie pracy
serca szmery (murmurs) różniące się od tonów. Są one zazwyczaj
spowodowane nieprawidłowymi warunkami przepływu krwi pomiędzy
przedsionkami, komorami i zbiornikami tętniczymi.
Zjawiska akustyczne występujące w sercu można zarejestrować, otrzymując zapis w postaci fonokardiogramu. Do skóry w okolicy serca
przystawia się specjalny przetwornik. Wzmacnia się prądy elektryczne
wzbudzane w przetworniku przez drgania towarzyszące pracy serca
i rejestruje się je zazwyczaj za pomocą aparatu do elektrokardiografii.
I Krążenie wieńcowe
Krew tętnicza dopływa do mięśnia sercowego przez dwie tętnice
wieńcowe, lewą i prawą (arteria coronaria sinistra et dextra). W warunkach prawidłowych pomiędzy tymi tętnicami nie ma połączeń za pośred-
358
Krążenie
nictwem naczyń o większej średnicy. Połączenia takie występują tylko
pomiędzy naczyniami o średnicy około 40 um. W przypadku zaczopowania jednej z tętnic wieńcowych druga tętnica nie może jej zastąpić.
Przepływ krwi przez naczynia wieńcowe (coronary blood flow), a szczególnie przez lewą tętnicę, jest ściśle uzależniony od okresu cyklu pracy serca i od ciśnienia w aorcie. W okresie rozkurczu krew przepływa swobodnie
przez obie tętnice wieńcowe, natomiast na początku skurczu komór,
w miarę wzrastania ciśnienia w komorach, przepływ przez naczynia
wieńcowe gwałtownie się zmniejsza.
Ryc. 179. Ciśnienie krwi w aorcie i przepływ krwi przez lewą tętnicę wieńcową w czasie cyklu
pracy serca w przeliczeniu na mL/min.
W końcu skurczu izowolumetrycznego i na początku podokresu
maksymalnego wyrzutu ciśnienie w lewej komorze oraz ciśnienie śródścienne w samym mięśniu lewej komory przewyższają ciśnienie w aorcie.
Odgałęzienia w lewej tętnicy wieńcowej zostają zamknięte i krew cofa się
do aorty — występuje wsteczny przepływ krwi (backflow). W połowie
podokresu maksymalnego wyrzutu krew zaczyna przepływać przez lewą
tętnicę wieńcową i naczynia włosowate, aby w okresie zredukowanego
wyrzutu ponownie się zmniejszyć. Na początku rozkurczu przepływ przez
lewą tętnicę wieńcową osiąga swoje maksimum.
Serce
359
W prawej tętnicy wieńcowej przepływ krwi podlega podobnym wahaniom, zależnym od okresu cyklu pracy serca, z tą różnicą, że nie występuje
wsteczny przepływ krwi.
Przyspieszenie częstości skurczów serca jest szczególnie niekorzystne
dla utrzymania prawidłowej prężności tlenu (Po2) w komórkach mięśnia
sercowego, zwłaszcza mięśnia lewej komory. W miarę przyspieszania
częstości skurczów wzrasta łączny czas, w którym przepływ krwi w lewej
tętnicy wieńcowej zostaje znacznie zmniejszony lub całkowicie zatrzymany. Niedostateczny przepływ wieńcowy jest przyczyną niedotlenienia
mięśnia sercowego.
Czynniki zmieniające przepływ
wieńcowy
Zarówno czynniki nerwowe, jak i humoralne zmieniają przepływ krwi
przez naczynia wieńcowe. Nagle występujące znaczne niedotlenienie mięśnia sercowego kilkakrotnie zwiększa przepływ wieńcowy. Z komórek
niedotlenionego mięśnia sercowego uwalnia się adenozyna i adenozyno-fosforany, również prostaglandyny PGE2, histamina i cholina. Związki te
działają silnie rozkurczająco na błonę mięśniową tętnic wieńcowych.
Zwiększenie prężności dwutlenku węgla we krwi (Pco2) również powoduje zwiększenie przepływu wieńcowego, ponieważ jednocześnie zmniejsza
się prężność tlenu (Po2) we krwi. Wazopresyna natomiast działa silnie
kurcząco na błonę mięśniową tętnic wieńcowych i zmniejsza przepływ
wieńcowy.
Impulsacja przywspółczulna i współczulna zmienia przepływ wieńcowy,
działając bezpośrednio na błonę mięśniową tętnic wieńcowych i pośrednio
wpływając na częstość i siłę skurczów serca. Acetylocholina — uwalniająca
się w sercu z zakończeń przywspółczulnych komórek zwojowych — zwiększa przepływ wieńcowy. Impulsacja współczulna, jak również adrenalina
i noradrenalina krążące we krwi, działając poprzez beta-receptor adrenergiczny, przyspieszają częstość i zwiększają siłę skurczów serca oraz
przepływ wieńcowy. Po zablokowaniu beta-receptorów i zniesieniu dodatnich efektów chrono- i inotropowych, zarówno impulsacja współczulna,
jak i krążące we krwi adrenalina i noradrenalina zmniejszają przepływ
wieńcowy krwi, działając poprzez alpha-receptor adrenergiczny.
Przepływ wieńcowy zwiększa się pod wpływem wytwarzanej prostacykliny PGI2 przez komórki śródbłonka i błony mięśniowej tętnic wieńcowych. Zmniejsza się zaś pod wpływem tromboksanu (TxA2) uwalnianego
z tr ombocytów i leukotrienów (LTC 4 i LT D 4 ) wytwar zanych
przez granulocyty obojętnochłonne.
360
Krążenie
Przemiana i odżywianie mięśnia sercowego
Przez naczynia wieńcowe przepływa około 5% krwi stanowiącej
pojemność minutową serca, czyli około 270 mL krwi na minutę w spoczynku. Mięsień sercowy jednocześnie zużywa około 10% całego zapotrzebowania organizmu na tlen. Czerpie energię niezbędną do skurczów ze
składników odżywczych dopływających z krwią, takich jak: glukoza,
mleczany, pirogroniany i wolne kwasy tłuszczowe — FFA.
I Krążenie duże
Krew tłoczona przez lewą komorę do aorty wypełnia zbiornik tętniczy
duży — Ras, z którego poprzez sieć naczyń włosowatych odpływa do
zbiornika żylnego dużego — Rvs.
I Zbiornik tętniczy duży
Zbiornik tętniczy jest pojęciem czynnościowym. Zbiornik tętniczy duży
(systemie arterial reservoir) zawiera krew wypełniającą wszystkie duże,
średnie i małe tętnice krążenia dużego. Zbiornik ten charakteryzują
następujące parametry:
•
•
•
•
pojemność;
ciśnienie;
prędkość przepływu krwi;
fala tętna.
W zbiorniku tętniczym dużym mieści się około 550 mL krwi. Stanowi to
około 11 % całkowitej objętości krwi krążącej w organizmie (TBV)
Ciśnienie tętnicze krwi
Ciśnienie tętnicze krwi (arterial blood pressure) zależy od dopływu i od
odpływu krwi ze zbiornika tętniczego. Przy zrównoważonym odpływie
i dopływie średnie ciśnienie panujące w tym zbiorniku nie ulega zmianom,
jeżeli nie zmienia się jednocześnie napięcie ścian tętnic wywołane skurczem
błony mięśniowej.
Krążenie duże
361
Ciśn ien ie w zb io rn ik u tę tn icz y m d uży m wa ha się w za leż nośc i od
okresu cyklu pracy serca. W okresie maksymalnego wyrzutu lewej komory
jest najwyższe i określane jako ciśnienie skurczowe (systolic arterial pres sure). W praktyce lekarskiej mierzone w tętnicy ramiennej na poziomie
ujścia lewej komory do aorty {ostium aortae) wynosi 16 kPa (120 mm Hg).
W rozkurczu i w okresie skurczu izowolu metrycz nego komór, przed
otworzeniem się zastawek aorty, ciśnienie jest najniższe, czyli rozkurczowe
(diastolic a rte rial pressu re) i wynosi 9,3 kPa (7 0 mm Hg).
- Tętnienie
Ryc. 180. Ciśnienie krwi w tętnicach, tętniczkach, naczyniach włosowatych i żyłach krążenia
dużego. Gradient ciśnienia w tętnicach, w sieci naczyń włosowatych i w żyłach. Średnie
ciśnienie: tętnicze — 12,0 kPa, w naczyniach włosowatych przytętniczych — 4,3 kPa, i w naczyniach włosowatych przyżylnych 1,6 kPa.
Ciśnienie w czasie rozkurczu serca nie obniża się do zera dzięki temu,
że ściany zbiornika tętniczego są sprężyste. Krew tłoczona przez lewą
komorę do aorty w czasie każdego skurczu serca rozciąga ściany zbiornika
tętniczego i napięcie sprężyste jego ścian zapewnia utrzymanie ciśnienia
w okresach rozkurczu serca. Jest to zgodne z zasadą powietrzni. Energia
skurczów serca jest magazynowana w elastycznych ścianach zbiornika
tętniczego (force of recoil). Średnie ciśnienie tętnicze (mean arterial
pressure) w pozycji leżącej wynosi około 12 kPa (90 mm Hg), amplituda
wahań ciśnienia wynosi zaś 6,7 kPa (50 mm Hg). Prawidłowe wartości
ciśnienia skurczowego/ciśnienia rozkurczowego wynoszą 16/9,3 kPa
(120/70 mm Hg) i odnoszą się do pomiaru wykonywanego w tętnicy
ramiennej na poziomie ujścia lewej komory do aorty u człowieka pozo-
362
Krążenie
stającego w spoczynku w pozycji leżącej. Po zmianie pozycji na stojącą,
w czasie ruchu, a zwłaszcza w czasie pracy fizycznej, ciśnienia skurczowe
i rozkurczowe odpowiednio podwyższają się.
W pozycji stojącej zaznaczają się wyraźnie różnice w ciśnieniu w zbiorniku tętniczym dużym wywołane siłą ciążenia. Powyżej serca ciśnienie
tętnicze opada, a poniżej wzrasta o 0,10241 kPa na każdy centymetr
różnicy poziomów. Przy średnim ciśnieniu w zbiorniku tętniczym dużym
w pozycji stojącej na poziomie ujścia lewej komory do aorty równemu
13,3 kPa średnie ciśnienie w tętnicach głowy wynosi około 9,3 kPa,
w tętnicach stopy zaś około 26,7 kPa.
Przepływ krwi
W ciągu minuty w spoczynku w pozycji leżącej dopływa do zbiornika
tętniczego dużego około 5,4 L krwi, co równa się pojemności minutowej
lewej komory. Tyle samo krwi odpływa ze zbiornika do sieci naczyń
włosowatych.
Ryc. 181. Przepływ burzliwy (linia przerywana) i przepływ laminarny krwi przez naczynie
krwionośne.
Krew przepływa w zbiorniku tętniczym dużym zgodnie z gradientem
ciśnienia od serca aż do naczyń włosowatych. Przepływ krwi ma charakter
pulsujący (pulsatile blood flow). Prędkość zwiększa się w czasie skurczu
izotonicznego komór w okresie maksymalnego wyrzutu i zmniejsza do
zera w czasie rozkurczu serca. Krew wtłaczana do aorty przepływa przez
nią ze średnią prędkością 0,6 m/s. W miarę oddalania się od serca średnia
prędkość przepływu krwi (mean velocity of blood) w tętnicach małego
kalibru zmniejsza się do kilku centymetrów na sekundę.
Krążenie duże
363
Ryc. 182. Ciśnienie i przepływ krwi w aorcie i w pniu płucnym.
Odpływ krwi ze zbiornika tętniczego dużego zależy przede wszystkim
od światła naczyń oporowych, czyli od światła małych tętniczek, oraz od
właściwości krwi — jej lepkości (viscosity of blood).
Fala tętna
Lewa komora, wtłaczając do aorty w czasie jednego skurczu objętość
wyrzutową krwi, powoduje jednoczesny wzrost ciśnienia i powstanie fali
ciśnieniowej oraz odkształcenie się ścian tętnic. Fala ciśnieniowa z towarzyszącym jej odkształceniem ścian tętnic, określana jako fala tętna
(arteriał pulse), rozchodzi się wzdłuż ścian zbiornika tętniczego dużego od
364
Krążenie
Ryc. 183. Powstawanie fali tętna w aorcie w czasie wyrzutu krwi z lewej komory serca.
Ryc. 184. Fonokardiogram (1), sfigmogram lewej tętnicy szyjnej wspólnej (2) i elektrokardiogram (3) odprowadzenia II kończynowego zarejestrowane w czasie trzech cykli pracy serca.
Fonokardiogram: I, II i III ton serca. Sfigmogram: D — załamek dykrotyczny. Elektrokardiogram: załamki P, Q, R, S i T.
Krążenie duże
365
serca aż do naczyń przedwłosowatych tętniczych, a nawet do naczyń
włosowatych. Prędkość rozchodzenia się fali tętna zależy od elastyczności
ścian tętnic oraz ich przebiegu i mieści się w granicach od 5 do 9 m/s.
W tętnicach o ścianach elastycznych fala tętna przesuwa się wolniej,
natomiast w tętnicach o ścianach stwardniałych, mniej elastycznych,
rozchodzi się szybciej. W tętnicach o prostym przebiegu fala tętna
przesuwa się szybciej, w tętnicach krętych zaś wolniej.
Ryc. 185. Fala tętna odebrana z: a — tętnicy szyjnej wspólnej; b — tętnicy promieniowej.
Odkształcanie się ścian tętnicy w czasie przechodzenia przez nią fali
tętna może być odebrane za pomocą specjalnych czujników i zarejestrowane na papierze, najczęściej za pomocą aparatu do elektrokardiografii.
Zapisana fala tętna, czyli sfigmogram. charakteryzuje się ramieniem
wstępującym i zstępującym. Na ramieniu zstępującym zaznacza się niewielka oscylacja, zwana załamkiem lub falą dykrotyczną (dicrotic notch),
spowodowana odbiciem się słupa krwi o zamykającą się zastawkę aorty.
Rola naczyń oporowych
Błona mięśniowa małych tętniczek pozostaje pod stałym wpływem
impulsacji nerwowej z ośrodków naczyniozwężających. W zależności od
zapotrzebowania na tlen w jakimś obszarze naczyniowym rozszerzają się
w nim małe tętniczki i przepływ krwi zwiększa się. Jednocześnie w innych obszarach naczyniowych w tym samym czasie dochodzi do dalszego
zwężania się światła małych tętniczek.
Czynność naczyń oporowych, czyli małych tętniczek, w dużym krążeniu można porównać do czynności kurków. Są one stale prawie
Ryc. 186. Czynność naczyń oporowych przedstawiona w postaci kurków: a — naczynia
oporowe całkowicie rozszerzone — przekrwienie; b — naczynia oporowe częściowo zwężone — prawidłowe krążenie; c — naczynia oporowe całkowicie zwężone — niedokrwienie.
Strzałki symbolizują filtrację i resorpcję przez ścianę naczynia włosowatego.
całkowicie zakręcone i przepuszczają tylko niewielką ilość krwi. W wyniku
zapotrzebowania na krew przez poszczególne narządy kurki regulujące
366
Krążenie
dopływ krwi zostają odkręcone, w tym samym czasie zaś kurki w innych
narządach zostają zakręcone. Pojemność krwi odpływająca ze zbiornika
tętniczego nie ulega zmianie.
Krążenie duże
367
Jednoczesne odkręcanie większej liczby kurków zwiększa odpływ krwi
i obniża ciśnienie w zbiorniku tętniczym. Mechanizmy kontrolujące układ
sercowo-naczyniowy przyspieszają natychmiast pracę serca i jego pojemność minutowa zwiększa się tak, aby odpływ krwi został zrównoważony
przez dopływ.
W obrębie naczyń oporowych zachodzi największy spadek ciśnienia
krwi. Krew dopływająca do małych tętniczek na poziomie serca ma średnie
ciśnienie około 12 kPa, czyli średnie ciśnienie w zbiorniku tętniczym. Po
przepłynięciu przez naczynie oporowe średnie ciśnienie krwi obniża się do
4,6 kPa.
Opór naczyniowy
Energia potencjalna związana z różnicą ciśnień pomiędzy zbiornikiem
tętniczym (Ras) i zbiornikiem żylnym (Rvs) zużywana jest na pokonanie
oporu naczyniowego. Całkowity obwodowy opór naczyniowy — TPR (total
peripheral vessels resistance) — obejmuje wszystkie naczynia krążenia
dużego: tętnice, tętniczki, naczynia przedwłosowate tętnicze, naczynia
włosowate i żyły. Decydujące jednak znaczenie dla wielkości oporu
naczyniowego mają naczynia oporowe, czyli małe tętniczki i naczynia
przedwłosowate tętnicze. Mała średnica naczyń oporowych jest powodem
największego spadku ciśnienia w obrębie tych naczyń.
Opór naczyniowy — R (blood vessels resistance) — jest wprost
proporcjonalny do różnicy ciśnień pomiędzy zbiornikami tętniczymi
i żylnymi (P) i odwrotnie proporcjonalny do pojemności minutowej serca
(Q).
Opór naczyniowy wyrażany jest w jednostkach obwodowego oporu
naczyniowego — PRU (peripheral resistance unit).
Jednostkę oporu naczyniowego stanowi stosunek różnicy ciśnień krwi
w kPa (mm Hg) — P, do ilości przepływającej krwi w mL/s — F.
Jednostka oporu naczyniowego może być wyrażona jako pochodna
stałej tętniczo-żylnej różnicy ciśnień:
368
Krążenie
lub jako pochodne stałego przepływu krwi:
Po dokonaniu zaokrągleń cyfr można przyjąć, że różnica średnicl
ciśnień (P) pomiędzy zbiornikami tętniczymi i żylnymi krążenia dużego
wynosi u człowieka 13,3 kPa. W czasie 1 sekundy przepływa ze zbiornik;
tętniczego dużego do zbiornika żylnego dużego 100 mL krwi. Te wartość
wyznaczają jedną jednostkę obwodowego oporu naczyniowego.
1 PPU — różnica ciśnień 13,3 kPa (100 mm Hg)
przepływ krwi 100 m L/s
Całkowity obwodowy opór naczyniowy w warunkach przeciętnych
życia człowieka wynosi jedną jednostkę oporu naczyniowego — TPR =
= 1 PRU. W czasie skurczu naczyń oporowych całkowity obwodowy
opór naczyniowy może zwiększyć się do 4 jednostek — TPR = 4 PRU,
w czasie ich rozkurczu zaś może się zmniejszyć do */ 4 jednostki —
TPR = 0,25 PRU.
Całkowity płucny opór naczyniowy (total pulmonary vessels resistance)
jest około dziesięciu razy mniejszy od całkowitego obwodowego oporu
naczyniowego (TPR). Wynika to stąd, że różnica pomiędzy średnim
ciśnieniem w zbiorniku tętniczym płucnym i zbiorniku żylnym płucnym
jest niewielka, wynosi bowiem około 1 kPa. Całkowity płucny opór
naczyniowy w stanach chorobowych może ulec zwiększeniu nawet do 1,0
PRU lub zmniejszeniu do 0,03 PRU.
I Zbiornik żylny duży
Zbiornik żylny duży (systemie venous reservoir — Rvs) tak jak zbiornik tętniczy jest również pojęciem czynnościowym. W zbiorniku tym
znajduje się krew wypełniająca duże, średnie i małe żyły krążenia dużego.
Zbiornik żylny duży charakteryzuje się:
• pojemnością;
• ciśnieniem;
• prędkością przepływu krwi.
W zbiorniku żylnym dużym gromadzi się około 2,7 L krwi, czyli 54%
całkowitej objętości krwi krążącej w organizmie (TBV).
Krążenie duże
369
Ciśnienie w zbiorniku żylnym dużym
Ciśnienie krwi w zbiorniku żylnym dużym zależy od miejsca pomiaru
i pozycji ciała. Ciśnienie w żyle głównej górnej (yena cava superior) i w żyle
głównej dolnej (yena cava inferior) przy ujściu do prawego przedsionka
(atrium dextrum) wynosi średnio około 0,5 kPa w pozycji leżącej badanego. Ciśnienie to przyjęto nazywać ciśnieniem żylnym centralnym (central
venous pressure) w odróżnieniu od ciśnienia żylnego obwodowego (peripheral venous pressure), które zależy od wielu czynników.
Poczynając od naczyń włosowatych i posuwając się w kierunku prawego przedsionka, ciśnienie krwi w żyłach obniża się. W małych żyłach
wynosi średnio około 2 kPa, w dużych żyłach zaś, na zewnątrz klatki
piersiowej, średnio około 0,6 kPa. Wartości te odnoszą się do pomiarów
ciśnienia w spoczynku w pozycji leżącej, w żyłach na poziomie prawego
przedsionka.
W pozycji stojącej ciśnienie żylne centralne nie zmienia się istotnie,
natomiast występują duże różnice ciśnień w zbiorniku żylnym powyżej
i poniżej prawego przedsionka. Poniżej prawego przedsionka ciśnienie
wzrasta dzięki sile ciążenia o 0,1 kPa na każdy centymetr różnicy
poziomów i osiąga w pozycji stojącej, w bezruchu, w obrębie żył stopy
około 13,3 kPa.
W żyłach powyżej serca ciśnienie jest niższe od ciśnienia atmosferycznego i żyły te są zapadnięte. W pozycji stojącej lub siedzącej w zatokach
opony twardej (sinus durae matris) panuje ciśnienie ujemne —1,3 kPa.
Ciśnienie żylne centralne podlega również niewielkim wahaniom zależnym od:
• ruchów oddechowych klatki piersiowej;
• pracy serca.
wO
370
Krążenie
W czasie wdechu ciśnienie w jamie opłucnej (cavum pleurale) obniża
się z —0,3 kPa do —0,8 kPa. Obniżenie się ciśnienia w jamie opłucnej
przenosi się na śródpiersie {mediastinum) i na przebiegające tam duże żyły.
W czasie wydechu ciśnienie w jamie opłucnej podwyższa się. Zgodnie
z rytmem oddechowym ciśnienie żylne centralne mieści się w granicach od
0,2 kPa w czasie wdechu do 0,8 kPa w czasie wydechu. Średnie ciśnienie
wynosi 0,5 kPa.
Ryc. 188. Rozkład ciśnień w zbiorniku tętniczym dużym (po prawej)
i w zbiorniku żylnym dużym (po
lewej) na różnych ich wysokościach
u człowieka stojącego, w wyniku
działania ciśnienia hydrostatycznego i siły ciążenia.
Obniżenie się podstawy serca w czasie skurczu izotonicznego komór
powoduje rozciąganie przedsionków i napływ do nich krwi z żył. Również
w rozkurczu, w okresie szybkiego wypełniania się komór krwią, dochodzi
do obniżenia się ciśnienia żylnego centralnego i szybkiego pulsującego
przepływu krwi w żyłach wpadających do prawego przedsionka.
Krążenie duże
371
Przepływ krwi w zbiorniku żylnym
Warunki przepływu krwi w zbiorniku żylnym całkowicie się różnią od
warunków przepływu w zbiorniku tętniczym.
Krew napływa do prawego przedsionka dzięki:
• ssącemu działaniu ruchów oddechowych klatki piersiowej i ssącemu
działaniu serca, czyli tzw. sile od przodu (vis a fronte);
• resztkowemu gradientowi ciśnienia od małych żył aż do prawego przed
sionka, wytworzonemu dzięki skurczom lewej komory serca, czyli tzw.
sile od tyłu (vis a tergo);
• pompie mięśniowej — skurczom mięśni szkieletowych, czyli tzw. sile
z boku (vis a latere), uciskającym żyły i wyciskającym krew z żył
w kierunku serca, ponieważ zastawki żylne nie pozwalają krwi cofnąć się
na obwód.
Przez zbiornik żylny przepływa około 5,4 L krwi na minutę, a średnia prędkość przepływu krwi w żyłach głównych w pobliżu serca wynosi
do 0,4 m/s.
I Krążenie krwi w naczyniach włosowatych
Przepływ krwi przez naczynia włosowate jest bardzo wolny i wynosi
około 0^5 mm/s. Ponieważ naczynia włosowate są bardzo krótkie, krew
przez nie przepływa w czasie kilku sekund (1—2 s).
Naczynia włosowate zawierają tylko około 5% całkowitej objętości
krwi krążącej. Mimo niewielkiej objętości krwi, która się w nich znajduje,
odgrywają zasadniczą rolę w krążeniu wszystkich związków w organizmie.
W obrębie naczyń włosowatych zachodzi cała wymiana związków pomiędzy krwią i wszystkimi tkankami na zasadzie dyfuzji, filtracji i resorpcji.
W naczyniach włosowatych przytętniczych na poziomie serca ciśnienie krwi wynosi 4,6 kPa, w naczyniach włosowatych przyżylnych zaś
tylko 2 kPa. Tym samym ciśnienie obniża się w naczyniach włosowatych
0 2,6 kPa.
Filtracja w naczyniach włosowatych
W naczyniach włosowatych przytętniczych zachodzi filtracja wody
1 składników drobnocząsteczkowych przez pory w ścianie naczyń włoso-
372
Krążenie
Tętniczka
Żyta
Ryc. 189. Krążenie krwi w sieci naczyń włosowatych.
+ 2,00kPa (+15 mm Hg) Ciśnienie hydrostatyczne + 4,61' kPa (+35mm Hg) 3,33k Pa (~ 25mm H g) Ci śnienie onk otyc zne - 3,33 k Pa (- 25mm H g) 0,27kPa (-2 mm Hg)
Ciśnienie międzykomórkowe - 0,27 kPa (-2 mm Hg)
Ciśnienie-160kPa (-12mm Hg)
Ciśnienie filtracyjne
+ 1, 07 kPa (+8 mm Hg)
resorpcyjne
.
\\
y
Naczynia
wiedwłosowate
Resorpcja
Filtracja
Ryc. 190. Filtracja i resorpcja wody, składników drobnocząsteczkowych w naczyniach
włosowatych przytętniczych i przyżylnych.
Krążenie duże
373
watych do płynu tkankowego (interstitial fluid). Ciśnienie hydrostatyczne
krwi w naczyniach włosowatych przytętniczych wynosi 4,6 kPa, ciśnienie
onkotyczne osocza 3,3 kPa i ciśnienie płynu tkankowego 0,2 kPa.
W związku z tym ciśnienie filtracyjne (filtration pressure) krwi w naczyniach włosowatych przytętniczych wynosi +1,1 kPa.
W naczyniach włosowatych przyżylnych zachodzi proces przeciwny do
filtracji — resorpcja wody i związków w niej rozpuszczonych. Ciśnienie
onkotyczne białek osocza jest wyższe od ciśnienia hydrostatycznego krwi
i woda ze składnikami w niej rozpuszczonymi zostaje wciągana, czyli
resorbowana z płynu tkankowego do krwi. Ciśnienie resorpcyjne (resorption pressure) w naczyniach włosowatych przyżylnych wynosi 1,6 kPa.
Średnica porów pomiędzy komórkami śródbłonka naczyń włosowatych nie jest jednakowa. Naczynia włosowate w kłębuszkach nerkowych
mają pory o średnicy około 10 nm. Pory w naczyniach włosowatych
mięśni szkieletowych mają mniejszą średnicę niż w naczyniach kłębuszkow
nerkowych, w wątrobie zaś większą średnicę.
W zależności od stanu błony mięśniowej małych tętniczek i naczyń
przedwłosowatych tętniczych może przeważać filtracja lub resorpcja.
Rozszerzenie się małych tętniczek i naczyń przedwłosowatych tętniczych,
czyli przekrwienie, powoduje otwieranie się naczyń włosowatych, wypełnianie ich krwią i przewagę filtracji nad resorpcja. Przeciwnie, skurcz
błony mięśniowej małych tętniczek i naczyń przedwłosowatych tętniczych
prowadzi do niedokrwienia — zamykania się naczyń włosowatych, a w naczyniach otwartych i wypełnionych krwią resorpcja przeważa nad filtracją.
U człowieka w ciągu doby filtruje się do przestrzeni międzykomórkowych około 0,25% objętości krwi przepływającej przez naczynia
włosowate. Większość przefiltrowanego płynu dzięki resorpcji powraca do
krwi, a od 2 do 4 L płynu dostaje się do naczyń chłonnych.
I Krążenie chłonki
Chłonka odpływająca z tkanek przez przewód piersiowy (ductus
thoracicus) i przewód chłonny prawy {ductus lymphaticus dexter) odprowadza część płynu tkankowego przefiltrowanego przez ściany naczyń
krwionośnych. W chłonce występują te same składniki co w osoczu
pozbawionym białek oraz związki wielkocząsteczkowe, które ze względu
na swoją wielkość nie mogą dostać się do krwi wraz z płynem resorbowanym.
374
Krążenie
Cząsteczki białek wydzielane przez komórki, zwłaszcza przez komórki
wątrobowe, dostają się do krwi za pośrednictwem naczyń chłonnych.
W obrębie błony śluzowej jelita cienkiego trafiają do naczyń chłonnych
kwasy tłuszczowe o długich łańcuchach. Są one zestryfikowane do
triacylogliceroli i otoczone warstwą lipoprotein, tworząc chylomikrony.
W tej postaci są transportowane przez naczynia chłonne do krwi.
Zwiększony przepływ krwi przez naczynia włosowate intensywnie
pracującego narządu wzmaga filtrację i więcej chłonki tworzy się w nim.
Naczynia krwionośne
włosowate
Naczynia
oporowe
tętnicze
Naczynia chłonne
Ryc. 191. Filtracja wody i składników drobnocząsteczkowych w naczyniach włosowatych
z krwi do płynu tkankowego, a następnie do naczyń chłonnych. Rvs — zbiornik żylny duży,
Ras — zbiornik tętniczy duży, ad — prawy przedsionek, as — lewy przedsionek, Vd —
komora prawa, Vs — komora lewa.
Chłonka przepływa w naczyniach chłonnych dzięki:
rytmicznym skurczom dużych naczyń chłonnych (lymph ducts rhythmic
contractions);
skurczom mięśni szkieletowych (skeletal muscle contractions);
ujemnemu ciśnieniu w klatce piersiowej (negative intrathoracic pressure).
Ośrodki kontrolujące krążenie krwi
375
Kurczące się mięśnie szkieletowe uciskają naczynia chłonne i przepychają chłonkę w kierunku dużych naczyń. Zastawki w naczyniach
chłonnych nie pozwalają chłonce cofnąć się i zapewniają jej jednokierunkowy przepływ. Wahania ujemnego ciśnienia w klatce piersiowej w czasie
wdechu i wydechu wywierają działanie ssące i zapewniają napływ chłonki
do naczyń chłonnych biegnących przez śródpiersie.
I Ośrodki kontrolujące krążenie krwi
Kontrola krążenia krwi w organizmie realizowana jest za pośrednictwem dwóch efektorów. Są to:
• mięsień sercowy;
• mięśnie gładkie i komórki mięśniowe gładkie w ścia nach naczyń
krwionośnych.
Te dwa efektory mają własne ośrodki, czyli ośrodek sercowy i ośrodek
naczynioruchowy.
I Ośrodek sercowy
Serce przyspieszając swoje skurcze, zwiększa pojemność minutową krwi
przetłaczanej ze zbiorników żylnych do zbiorników tętniczych. Przyspieszenie częstości skurczów serca prowadzi do zwiększenia pojemności
minutowej serca i do podwyższenia ciśnienia tętniczego w zbiorniku
tętniczym. Zwolnienie częstości skurczów serca daje zazwyczaj w ostatecznym wyniku zmniejszenie pojemności minutowej serca i obniżenie ciśnienia
tętniczego w zbiorniku tętniczym.
Neurony ośrodka kontrolującego pracę serca znajdują się w różnych
strukturach układu nerwowego i pod względem czynnościowym dzielą się
na neurony zwiększające pracę serca i na neurony zmniejszające pracę
serca.
Praca serca zostaje zwiększona przede wszystkim dzięki przyspieszeniu
skurczów serca. Z tego względu neurony to wywołujące określa się jako
ośrodek przyspieszający pracę serca (cardio-acceleratory center). Natomiast zmniejszenie pracy serca wiąże się zazwyczaj ze zwolnieniem jego
skurczów. Neurony zwalniające pracę serca obejmuje się nazwą ośrodka
zwalniającego pracę serca (cardio-inhibitory center).
376
Krążenie
Ryc. 192. Ośrodek zwalniający pracę serca w jąd-l
rze grzbietowym nerwu błędnego i ośrodek przy-1
spieszający pracę serca w rogach bocznych czc-1
ści piersiowej rdzenia kręgowego Thl—Th5 oraz I
włókna eferentne biegnące do serca.
Ośrodek rdzeniowy przyspieszający pracę serca znajduje się w rogach
bocznych rdzenia kręgowego w części piersiowej w segmentach od pierwszego do piątego: Thl—Th5. Neurony tego ośrodka wysyłają impulsy do
serca we włóknach przedzwoj owych do zwojów pnia współczulnego {ganglia
trunci sympathici) i zwojów współczulnych szyjnych górnego, środkowego
i dolnego {ganglia cervicalia superius, medium et inferius).
Ośrodki kontrolujące krążenie krwi
377
Komórki ze zwojów współczulnych przewodzą impulsy do serca przez
swoje wypustki stanowiące włókna Cs — zazwojowe współczulne. Najwięcej włókien zazwojowych biegnie do serca od zwoju szyjno-piersiowego, czyli gwiaździstego (ganglion cewicothoracicum sive stellatum),
utworzonego ze zwoju współczulnego szyjnego dolnego i pierwszego zwoju
piersiowego. Z zakończeń włókien zazwojowych typu Cs uwalnia się
noradrenalina (NA) działająca przyspieszająco na częstość skurczów
serca.
Ośrodek zwalniający pracę serca znajduje się w rdzeniu przedłużonym i
stanowią go neurony należące do jądra grzbietowego nerwu błędnego
(nucleus dorsalis nervi vagi). Neurony tego jądra zwalniają pracę serca
za pośrednictwem włókien eferentnych biegnących do serca w ner wach błędnych, czyli włókien typu B, i przywspółczulnych komórek
zwojowych znajdujących się w samym sercu. Pod wpływem impulsacji
z jądra grzbietowego nerwu błędnego przywspółczulne komórki zwojowe uwalniają ze swych zakończeń acetylocholinę (Ach), która działa na
układ przewodzący serca i na pozostałe komórki mięśnia przedsionków
i komór.
Ryc. 193. Schemat ośrodkowej kontroli pracy serca.
U człowieka ośrodek zwalniający pracę serca wykazuje stałą przewagę
nad ośrodkiem przyspieszającym pracę serca. Wyłączenie wpływu obu
ośrodków na serce, w wyniku jednoczesnego zablokowania receptorów
adrenergicznych i cholinergicznych, powoduje w spoczynku zwiększenie
częstotliwości skurczów serca z 1,2 do 1,7 Hz, czyli z 72 na minutę do
około 100 na minutę.
378
Krążenie
Ośrodek przyspieszający pracę serca i ośrodek zwalniający jego pracę
stanowią końcową wspólną drogę dla impulsacji biegnącej z:
• kory mózgu i z podwzgórza;
• receptorów w układzie sercowo-naczyniowym.
Z kory mózgu i z podwzgórza biegną impulsy nerwowe do ośrodków I
sercowych za pośrednictwem neuronów tworu siatkowatego.
Pobudzenie pól ruchowych w korze mózgu oraz ośrodków motywacyjnych w podwzgórzu powoduje hamowanie czynności neuronów jądra
grzbietowego nerwu błędnego. Ośrodek w części piersiowej rdzenia
kręgowego, przyspieszający pracę serca, uzyskuje przewagę i częstość j
skurczów serca zwiększa się.
Impulsacja z receptorów w układzie sercowo-naczyniowym na drodze
odruchowej zwalnia lub przyspiesza częstość skurczów serca. Rozciągnięcie ścian tętnic przez podwyższone ciśnienie krwi drażni baroreceptory
w ścianie łuku aorty i w zatokach tętnic szyjnych wewnętrznych. Pod
wpływem impulsacji biegnącej od baroreceptorów przez włókna aferentne
nerwów językowo-gardłowych (n. IX) i błędnych (n. X) ośrodek zwalniający pracę serca zostaje pobudzony i częstość skurczów serca maleje.
Przeciwnie — spadek ciśnienia tętniczego powoduje przyspieszenie pracy
serca na drodze odruchowej dzięki temu, że ośrodek zwalniający pracę
serca zostaje zahamowany.
Częstość skurczów serca ulega zmianie pod wpływem tzw. odruchu
Bainbridge'a oraz podrażnienia chemoreceptorów w ścianach naczyń
wieńcowych, czyli odruchu Bezolda-Jarischa. Odruch, czy też efekt Bainbridge'a — ponieważ łuk odruchowy nie został wykazany — polega na
przyspieszeniu skurczów serca w odpowiedzi na szybkie wypełnianie krwią
zbiornika żylnego dużego. W odruchu Bezolda-Jarischa z chemoreceptorów naczyń wieńcowych (coronary chemoreflex) serce zwalnia swoją
pracę wskutek podrażnienia receptorów w naczyniach wieńcowych przez
weratrynę lub nikotynę.
I Ośrodek naczynioruchowy
Skurcz lub rozkurcz mięśni gładkich w ścianie małych tętniczek zmienia
opór naczyniowy. Skurcz błony mięśniowej małych tętniczek zwiększa
opór naczyniowy, a tym samym zmniejsza przepływ krwi ze zbiornika
tętniczego do zbiornika żylnego i podwyższa ciśnienie w zbiorniku
tętniczym. Rozkurcz błony mięśniowej małych tętniczek prowadzi do
Ośrodki kontrolujące krążenie krwi
379
zjawiska przeciwnego. Opór naczyniowy zmniejsza się, przepływa więcej
krwi ze zbiornika tętniczego do żylnego i ciśnienie w zbiorniku tętniczym
obniża się.
Światło małych tętniczek jest kontrolowane ogólnie na drodze:
• nerwowej przez ośrodek naczynioruchowy (vasomotor center) za po
średnictwem nerwów naczynioruchowych;
• humoralnej przez ośrodki nerwowe kontrolujące wydzielanie hormo
nów, szczególnie przez układ reninowo-angiotensynowy;
i kontrolowane miejscowo przez:
• wyzwalane miejscowo odruchy i odruchy aksonowe;
• działające miejscowo czynniki naczyniorozszerzające i naczyniozwężające.
Ośrodek naczynioruchowy znajduje się w rdzeniu przedłużonym w tworze siatkowatym i składa się z dwóch części:
• presyjnej (pressor area) — zwężającej naczynia krwionośne;
• depresyjnej (depressor area) — rozszerzającej naczynia krwionośne.
Część presyjna
Neurony części presyjnej ośrodka naczynioruchowego w rdzeniu przedłużonym wysyłają wypustki do neuronów w rogach bocznych rdzenia
kręgowego w części piersiowej i lędźwiowej. Neurony rogów bocznych
przekazują pobudzenie do mięśni gładkich w ścianach naczyń krwionośnych za pośrednictwem neuronów w zwojach współczulnych. Włókna
nerwowe naczyniozwężające typu Cs przewodzą impulsy do naczyń
krwionośnych, utrzymując błonę mięśniową w stałym skurczu. Dzięki
temu przepływ krwi ze zbiornika tętniczego do zbiornika żylnego jest
regulowany na drodze nerwowej.
Pobudzenie neuronów części presyjnej ośrodka naczynioruchowego
wzmaga impulsację we włóknach naczyniozwężających, przepływ krwi ze
zbiornika tętniczego do żylnego zmniejsza się i ciśnienie w zbiorniku
tętniczym wzrasta.
Część presyjna ośrodka naczynioruchowego jest pobudzana przez:
• ośrodki z wyższych pięter mózgowia — z kory mózgu i układu
limbicznego, za pośrednictwem podwzgórza i tworu siatkowatego
śródmózgowia;
380
Krążenie
Naczynia
oporowe
tętnicze
Naczynia włosowate
Ryc. 194. Część presyjna ośrodka naczynioruchowego w rdzeniu przedłużonym, droga
impulsacji wywołującej skurcz błony mięśniowej naczyń oporowych. Rvs — zbiornik żylny
duży, Ras — zbiornik tętniczy duży, ad — przedsionek prawy, as — przedsionek lewy,
Vd — komora prawa, Vs — komora lewa.
Ośrodki kontrolujące krążenie krwi
381
Rdzeń
przedłużony
Pień współczulny
Naczynio
oporowe
tętnicze
Naczynia włosowate
Ryc. 195. Część depresyjna ośrodka naczynioruchowego w rdzeniu przedłużonym, hamująca
aktywność neuronów unerwiających błonę mięśniową naczyń oporowych. Rvs — zbiornik
żylny duży, Ras — zbiornik tętniczy duży, ad — przedsionek prawy, as — przedsionek lewy,
Vd — komora prawa, Vs — komora lewa.
382
Krążenie
ośrodek oddechowy w rdzeniu przedłużonym;
aferentne impulsy: bólowe i z chemoreceptorów kłębuszków szyjnych
i kłębków aortowych;
zmniejszenie prężności tlenu (P O2) we krwi tętniczej;
zwiększenie prężności dwutlenku węgla (Pco2) we krwi tętniczej.
Część depresyjna
Pobudzone neurony części depresyjnej ośrodka naczynioruchowego
hamują aktywność neuronów w rogach bocznych rdzenia kręgowego
i tym samym hamują impulsację we włóknach nerwowych naczyniozwężających. Małe tętniczki rozszerzają się, opór naczyniowy zmniejsza się
i więcej krwi przepływa ze zbiornika tętniczego do zbiornika żylnego.
Ciśnienie krwi w zbiorniku tętniczym obniża się.
Część depresyjna ośrodka naczynioruchowego w rdzeniu przedłużonym jest aktywowana pod wpływem:
• impulsacji z baroreceptorów ze ścian łuku aorty i zatoki tętnicy szyjnej
wewnętrznej;
• zmniejszonej prężności dwutlenku węgla (Pco2) we krwi tętniczej.
W pewnej niezależności od ośrodka naczynioruchowego w rdzeniu
przedłużonym pozostaje współczulny układ rozszerzający naczynia krwionośne w mięśniach szkieletowych (sympathetic vasodilator system). Zaczyna się on w korze mózgu, skąd impulsy biegną za pośrednictwem
neuronów podwzgórza i tworu siatkowatego śródmózgowia do neuronów rogów bocznych rdzenia kręgowego. Impulsy biegnące od tych
neuronów powodują rozszerzenie naczyń krwionośnych w mięśniach
szkieletowych i zwiększają przepływ krwi przez nie.
Regulacja ciśnienia
tętniczego krwi
Ciśnienie w zbiorniku tętniczym dużym stanowi wypadkową:
dopływu do zbiornika krwi tłoczonej przez serce;
odpływu krwi, który w znacznym stopniu zależy od stanu błony
mięśniowej małych tętniczek.
Ośrodki kontrolujące krążenie krwi
383
Ryc. 196. Schemat ośrodkowej kontroli przepływu krwi ze zbiornika tętniczego dużego
(Ras) do zbiornika żylnego dużego (Rvs). Impulsacja wysyłana przez neurony w rogach
bocznych rdzenia kręgowego i wydzielona adrenalina wywołują skurcz błony mięśniowej
naczyń oporowych ( + ), zwiększają całkowity obwodowy opór naczyniowy i zmniejszają
przepływ krwi pomiędzy zbiornikami.
Odruchy i ośrodki
Dopływ krwi do zbiornika tętniczego przede wszystkim jest kontrolowany przez ośrodek sercowy, odpływ zaś przez ośrodek naczynioruchowy. Oba te ośrodki współdziałają ze sobą, pozostając stale pod
wpływem impulsacji aferentnej z baroreceptorow.
Nasilenie impulsacji z baroreceptorow pobudza:
• ośrodek zwalniający pracę serca;
• część depresyjną ośrodka naczynioruchowego.
Jednocześnie hamuje:
• ośrodek przyspieszający pracę serca;
• część presyjną ośrodka naczynioruchowego.
Przeciwny skutek występuje po zmniejszeniu się impulsacji z baroreceptorow.
Po każdym skurczu serca i przesunięciu się fali tętna wzdłuż tętnic
biegnie salwa impulsów od baroreceptorow do rdzenia przedłużonego.
384
Krążenie
Dzięki temu ciśnienie w zbiorniku tętniczym stale jest regulowane w zależności od zapotrzebowania organizmu. W stanie spoczynku jest ono
utrzymywane na stałym poziomie. Największe skupienia baroreceptorów
znajdują się w zatokach tętnic szyjnych wewnętrznych i w łuku aorty, ale
występują one również w innych miejscach układu sercowo-naczyniowego: w ścianach przedsionków, w ścianach lewej komory i w ścianach
naczyń krążenia płucnego. Baroreceptory są wrażliwe na rozciąganie.
W związku z tym wskutek zwyżki ciśnienia krwi zostają podrażnione
rozciągnięciem ścian naczyń krwionośnych i jam serca.
Odruchy neurohormonalne
Zapotrzebowanie ze strony organizmu prowadzi do wydzielania hormonów działających na układ sercowo-naczyniowy. Silne emocje, znaczny
wysiłek fizyczny, utrata krwi, oziębienie wywołują wydzielanie adrenaliny
i noradrenaliny przez rdzeń nadnerczy. Impulsy z kory mózgu i układu
limbicznego biegną przez podwzgórze, śródmózgowie do części piersiowej
rdzenia kręgowego, gdzie znajdują się neurony, których aksony biegną
w nerwach trzewnych do rdzenia nadnerczy {medulla glandulae suprarenalis). Pod wpływem wydzielonych do krwi hormonów rdzenia
nadnerczy ciśnienie skurczowe w zbiorniku tętniczym podwyższa się. Choć
opór naczyń obwodowych może zmniejszyć się, to jednak częstość skurczów serca ulega przyspieszeniu i pojemność minutowa serca zwiększa się.
W czasie silnych emocji lub utraty znacznej ilości krwi, połączonej
z dużym obniżeniem ciśnienia w zbiorniku tętniczym, zostaje również
wydzielony w większych ilościach hormon podwzgórza — wazopresyna.
Ośrodki w podwzgórzu wyzwalające stany emocjonalne jednocześnie
powodują uwalnianie wazopresyny z części nerwowej przysadki (pars
nervosa hypophysis).
Utrata krwi prowadzi do zmniejszenia się objętości krwi w zbiorniku
żylnym i w zbiorniku tętniczym, co powoduje brak stałej impulsacji z:
• receptorów objętościowych w ścianach dużych żył;
• baroreceptorów zatok tętnic szyjnych i łuku aorty.
Następuje odruchowe wydzielanie wazopresyny. Hormon ten, działając
kurcząco na mięśnie gładkie naczyń krwionośnych, zwiększa całkowity
obwodowy opór naczyniowy (TPR) oraz zmniejsza pojemność zbiorników
krwi: tętniczego i żylnego, dostosowując je do objętości krwi krążącej
w łożysku krwionośnym (patrz ryc. 120, str. 230).
Ośrodki kontrolujące krążenie krwi
385
Układ reninowo-angiotensynowy
Układ reninowo-angiotensynowy (renin-angiotensyn system) stale
uczestniczy w ogólnej regulacji ciśnienia tętniczego krwi. Komórki wydzielające pro-reninę występują w wątrobie, w mózgowiu i w innych
narządach, ale zasadniczym miejscem wydzielania aktywnej reniny są
komórki aparatu przykłębuszkowego nerek (apparatus iuxtaglomerularis).
Komórki wydzielające reninę znajdują się pomiędzy plamką gęstą {macula
densa) kanalika nerkowego krętego dalszego oraz błoną mięśniową
tętniczki doprowadzającej i tętniczki odprowadzającej kłębuszka nerkowego (patrz ryc. 253, str. 476).
Renina jest glikoproteiną o masie cząsteczkowej 37,3 kDa i 340
aminokwasach, o właściwościach enzymatycznych i okresie połowicznego
rozpadu (T7 2) wynoszącym 80 minut. Renina odcina dziesięciopeptydowy łańcuch od angiotensynogenu — białka osocza należącego do
alpha-2 globulin. Dziesięciopeptyd, czyli angiotensyna I, jest związkiem
nieczynnym fizjologicznie. Przepływając we krwi przez naczynia krwionośne w płucach, jest zamieniana na ośmiopeptyd — angiotensynę II,
fizjologicznie aktywną. Zachodzi to pod wpływem enzymu konwertującego
angiotensynę I do angiotensyny II (ACE — angiotensin-converting enzyme) znajdującego się na komórkach śródbłonka naczyniowego.
Angiotensyna II wiąże się z kilkoma typami receptorów błonowych.
Receptorem występującym w błonie komórkowej komórek mięśniowych
gładkich w ścianach naczyń krwionośnych jest receptor AT1A. Angiotensyna II, wiążąc się z tym receptorem aktywuje fosfolipazę C za pośrednictwem białka błonowego G i wywołuje zwiększenie stężenia wolnych jonów
Ca2+ w cytoplazmie komórek mięśniowych oraz skurcz tych komórek.
Angiotensyna II jest hormonem najsilniej kurczącym błonę mięśniową
naczyń krwionośnych, zwiększającym całkowity obwodowy opór naczyniowy (TPR) i podwyższającym ciśnienie tętnicze krwi zarówno skurczowe, jak i rozkurczowe.
Angiotensyna II szybko jest rozkładana we krwi. Okres jej połowicznego rozpadu (T J/ 2) wynosi 1—2 minuty i zachodzi pod wpływem
enzymów objętych wspólną nazwą angiotensynaz krwi. Powstający pod
wpływem angiotensynaz jeden z peptydów — siedmiopeptyd — zachowuje część aktywności angiotensyny II i nosi nazwę angiotensyny III.
Bezpośrednim czynnikiem wywołującym wydzielanie reniny z nerek jest
spadek ciśnienia tętniczego krwi w tętniczkach nerkowych. Istnieją liczne
czynniki pośrednie, które obniżają ciśnienie tętnicze w tętniczkach nerkowych. Są to:
13 — Fizjologia człowieka
386
Krążenie
•
•
•
•
zmniejszenie objętości płynu zewnątrzkomórkowego (ECF);
zmniejszenie całkowitej objętości krwi krążącej (TBV);
zmniejszenie stężenia jonów Na + w płynach ustrojowych;
obniżenie ciśnienia tętniczego krwi w zbiorniku tętniczym dużym,
występujące nawet przy zmianie pozycji ciała z leżącej na stojącą;
• zmniejszenie przepływu krwi tętniczej przez nerki na skutek skurczu
błony mięśniowej tętnic nerkowych pod wpływem impulsacji współczulnej lub krążącej we krwi adrenaliny i noradrenaliny.
I
Miejscowa regulacja przepływu
krwi
Miejscowe mechanizmy regulujące (local regulatory mechanisms) przepływ krwi przez poszczególne narządy wiążą się z:
• czynnikami nerwowymi;
• czynnikami humoralnymi;
• miejscową autoregulacją.
Stopień rozciągnięcia ścian naczyń tętniczych warunkuje siłę skurczu
mięśni gładkich. Zróżnicowana wrażliwość na rozciąganie błony mięśniowej tętnic doprowadzających krew do poszczególnych narządów stabilizuje
przepływ krwi przez te narządy i w pewnym stopniu uniezależnia go od
ogólnego ciśnienia tętniczego.
Podrażnienie interoreceptorow w narządach wewnętrznych wywołuje
na drodze odruchowej rozszerzenie lub zwężenie światła naczyń tętniczych. Zazwyczaj następuje osłabienie eferentnej impulsacji naczyniozwężającej, wysyłanej przez ośrodki nerwowe, i błona mięśniowa tętnic
rozkurcza się. Impulsacja od receptorów, biegnąca we włóknach aferentnych, może również być przewodzona antydromowo do mięśni gładkich
naczyń krwionośnych i wywołać ich rozkurcz na drodze odruchu aksonowego (axon reflex). Rozszerzenie naczyń skórnych po podrażnieniu
skóry zachodzi na zasadzie odruchu aksonowego.
Pod wpływem wielu czynników fizycznych i chemicznych błona mięśniowa małych tętniczek rozkurcza się i światło tych naczyń powiększa się,
lub też kurczy i światło tych naczyń zmniejsza się.
Do czynników rozkurczających (vasodilators) działających miejscowo
bezpośrednio na komórki mięśniowe gładkie należą:
Miejscowa regulacja przepływu krwi
387
wzrost temperatury;
zwiększenie prężności dwutlenku węgla (Pco2);
zwiększenie wartości pH; wzrost ciśnienia
osmotycznego;
miejscowe zwiększenie stężenia mleczanów, histaminy, adenozyny,
jonów K+, prostaglandyn (PGE 2). prostacykliny (PGI2) i przedsionkowego peptydu natriuretycznego (ANP).
Zmniejszenie prężności tlenu (Po2) w tkance działa również rozkurczająco na błonę mięśniową małych tętniczek.
Działanie przeciwne, to jest bezpośrednio kurczące błonę mięśniową
małych tętniczek (vasoconstrictors), mają:
• miejscowe obniżenie temperatury;
• zmniejszenie prężności dwutlenku węgla (Pco2);
• zmniejszenie wartości pH;
• zmniejszenie stężenia mleczanów, histaminy, adenozyny, jonów K +,
prostaglandyny, prostacykliny i przedsionkowego peptydu natriurety
cznego.
Zwiększenie prężności tlenu i stężenia serotoniny ma działanie kur czące.
2+
Jony Ca
2
Ca *'
w ptynie
_____
cytoplazmie
zewnątrzkomórkowym
Jony
w
komórek
śródbfon
ka
naczyniowego
błony
mięśniowej
naczyń
krwionośnych
Ryc. 197. Schemat tworzenia
się tlenku azotu (NO) w
komórkach śródbłonka
naczyniowego.
------------ ►■ błony
mięśniowej
naczyń
krwionośnych
388
Krążenie
Przepływające wraz z krwią przez tętniczki substancje fizjologicznie
aktywne działają za pośrednictwem przekaźników chemicznych uwalnianych z komórek śródbłonka naczyniowego. Pod wpływem przekaźników
chemicznych rozkurczających miał się ze śródbłonka uwalniać śródbłonkowy czynnik rozkurczający — EDRF (endothelium-derived relaxant
factor), pod wpływem przekaźników chemicznych kurczących zaś śródbłonkowy czynnik kurczący — EDCF (endothelium-derived contracting
factor). Dalsze badania wykazały, że śródbłonkowym czynnikiem rozkurczającym jest powstający w komórkach śródbłonka tlenek azotu (NO),
śródbłonkowym czynnikiem kurczącym zaś są trzy polipeptydy o 21
aminokwasach i zbliżonej budowie: endotelina pierwsza — ET-1 (endoTrombina
Adrenalina
Interleukina 1
Wazopresyna
Anghłensyna II
Komórki
śródbłonka
naczyń
krwionośnych
-ENDOTELINA 1
Prosłacyklina
Tromboksan
Tlenek azotu
Skurcz btony
mięśniowej w
ścianach
naczyń
krwionośnych
Przedsionkowy
peptyd
nałriurełyczny
Ryc. 198. Schemat działania czynników pobudzających i hamujących wytwarzanie endoteliny przez komórki śródbłonka naczyniowego.
Krążenie płucne
389
thelin 1), endotelina druga — ET-2 (endothelin 2) i endotelina trzecia —
ET-3 (endothelin 3).
Pod wpływem acetylocholiny (Ach), bradykininy, wazoaktywnego
peptydu jelitowego (VIP) i Substancji P uwalnia się z komórek śródbłonka
naczyniowego tlenek azotu (NO) i tętniczki rozkurczają się. Przeciwnie —
adrenalina, wazopresyna, angiotensyna II i interleukina 1 (IL-1) pobudzają
komórki śródbłonka naczyniowego do uwalniania endoteliny pierwszej
(ET-1) i wywołują skurcz tętniczek. Endotelina pierwsza jest przekaźnikiem chemicznym najsilniej kurczącym błonę mięśniową tętniczek
w porównaniu z innymi fizjologicznie aktywnymi przekaźnikami chemicznymi.
Autoregulacja
Autoregulacja, czyli samoregulacja, dotyczy zarówno odpływu krwi
z całego zbiornika tętniczego dużego, jak i przepływu krwi przez poszczególne obszary naczyniowe. Wraz ze wzrostem ciśnienia tętniczego
ściany małych tętniczek są silniej rozciągane i w odpowiedzi na to silniej
się kurczą. Dzięki temu wzrost ciśnienia w zbiorniku tętniczym wywołuje
dalsze zwężenie światła naczyń oporowych i ilość krwi odpływająca ze
zbiornika nie ulega zmianie.
I Krążenie płucne
Prawa komora serca tłoczy do pnia płucnego około 5,4 L krwi na
minutę (SI: 90 mL/s), czyli tyle samo co lewa komora do aorty. Pojemność
minutowa obu komór serca jest jednakowa zarówno w czasie spoczynku,
jak i w czasie wysiłku fizycznego. Natomiast objętość krwi, która wypełnia
krążenie płucne, jest mniejsza niż w krążeniu dużym. W krążeniu płucnym,
to jest w zbiorniku tętniczym płucnym (pulmonary arterial reservoir),
w zbiorniku żylnym płucnym (pulmonary venous reservoir) i w sieci naczyń włosowatych, znajduje się tylko około 18% całkowitej objętości krwi
krążącej w organizmie (TBV).
Ciśnienie w zbiorniku tętniczym płucnym wynosi: skurczowe — 3,3
kPa, rozkurczowe — 0,9 kPa i średnie ciśnienie — około 2,0 kPa.
W zbiorniku żylnym płucnym średnie ciśnienie jest prawie takie samo jak
ciśnienie skurczowe w lewym przedsionku, czyli wynosi około 0,9 kPa.
Różnica ciśnień pomiędzy obu zbiornikami płucnymi, tętniczym i żylnym,
jest niewielka — średnio około 1,0 kPa.
390
Krążenie
Sieć naczyń włosowatych łączących oba zbiorniki oplata pęcherzyki
płucne. Ciśnienie krwi w naczyniach włosowatych zależy od pozycji ciała
i okolicy płuc. W pozycji pionowej w górnych partiach płuc większość
naczyń włosowatych jest zamknięta. To powoduje mniejszy przepływ krwi
w porównaniu z partiami środkowymi i dolnymi płuc.
W czasie wydechu ciśnienie powietrza w pęcherzykach płucnych jest
wyższe od ciśnienia atmosferycznego, uciska naczynia włosowate i zmniejsza przepływ krwi. W górnych partiach płuc ciśnienie powietrza pęcherzykowego (PA) przewyższa ciśnienie krwi w naczyniach włosowatych przytętniczych (Pa), a to z kolei jest wyższe od ciśnienia w naczyniach włosowatych przyżylnych (Pv), czyli PA>Pa>Pv. W środkowych i dolnych
partiach płuc rozkład ciśnień jest inny. W środkowych: Pa>PA>Pv,
Serce
Ryc. 200. Schemat krążenia płucnego,
czyli małego. Rap — zbiornik tętniczy
płucny, Rvp — zbiornik żylny płucny,
s.n.p. — sieć naczyń włosowatych w pęcherzykach płucnych, ad — prawy
przedsionek serca, as — lewy przedsionek serca, Vd — komora prawa
serca, Vs — komora lewa serca, Rvs —
krew napływająca ze zbiornika żylnego
dużego, Ras — krew odpływająca do
zbiornika tętniczego dużego.
Krążenie płucne
391
w dolnych zaś Pa > Pv > PA. W dolnych partiach płuc naczynia włosowate
są rozszerzone i tamtędy przepływa większość krwi. W pozycji leżącej na
plecach w tylnych partiach płuc naczynia są rozszerzone i większość krwi
przepływa przez nie.
Małe tętniczki w przeciwieństwie do krążenia dużego, mają słabo
rozwiniętą błonę mięśniową i nie spełniają funkcji naczyń oporowych.
Krew wtłoczona do zbiornika tętniczego płucnego przepływa do zbiornika
żylnego płucnego, natrafiając jedynie na opór ze strony sieci naczyń
włosowatych.
Ryc. 201. Przepływ krwi przez sieć naczyń włosowatych w górnych, środkowych i dolnych partiach płuc. PA — ciśnienie powietrza pęcherzykowego, Pv — ciśnienie krwi
w naczyniach włosowatych przyżylnych i Pa — ciśnienie krwi w naczyniach włosowatych
przytętniczych.
W naczyniach włosowatych płuc w warunkach prawidłowych nie
dochodzi do filtracji osocza. Ciśnienie onkotyczne białek osocza jest stale
wyższe od ciśnienia hydrostatycznego krwi i może zachodzić wyłącznie
resorpcja płynu tkankowego. Wzrost ciśnienia w krążeniu płucnym
powyżej granicy fizjologicznej stwarza warunki do wystąpienia ciśnienia
filtracyjnego i do gromadzenia się przefiltrowanego płynu w świetle
pęcherzyków płucnych. Stan ten nazywa się obrzękiem płuc (oedema
pulmonum).
392
Krążenie
I Krążenie krwi w mózgowiu
Przepływ krwi przez całe mózgowie — CBF (cerebral blood flow),
w przeciwieństwie do przepływu krwi przez inne narządy, nie zmienia się
istotnie w czasie pracy fizycznej i umysłowej, jak również w okresie
czuwania i snu. Przez mózgowie przepływa około 750 mL krwi na minutę.
Choć przepływ krwi przez całe mózgowie jest stały, to jednak w zależności
od stanu fizjologicznego organizmu występują zmiany w miejscowym
przepływie krwi przez poszczególne pola w korze mózgu.
Przepływ krwi przez całe mózgowie zależy przede wszystkim od
ciśnienia śródczaszkowego (intracranial pressure). Wzrost ciśnienia śródczaszkowego zmniejsza przepływ krwi przez mózgowie.
Ciśnienie śródczaszkowe wzrasta wskutek:
•
•
•
•
•
wzrostu ciśnienia tętniczego w obrębie tętnic mózgowych;
wzrostu ciśnienia żylnego w naczyniach żylnych mózgowych;
zwiększenia lepkości krwi;
miejscowego zwiększenia prężności dwutlenku węgla (Pco2);
miejscowego zmniejszenia prężności tlenu (Po2)Zwiększenie PCo2 i zmniejszenie PO2 w obrębie mózgowia wpływają silnie
rozkurczająco na mięśnie naczyń tętniczych. Naczynia rozszerzają się i
przepływ krwi znacznie zwiększa się. Po pewnym czasie wzrasta ciśnienie
śródczaszkowe i przepływ krwi zmniejsza się. Zmniejszenie w mózgu P Co2
Krążenie krwi w mózgowiu
393
i zwiększenie P Q 2 wywołują umiarkowany skurcz naczyń mózgowych
i zmniejszenie przepływu krwi.
Prężność dwutlenku węgla i tlenu w tkance mózgowej stanowi zasadniczy czynnik regulujący przepływ krwi przez mózgowie (CBF). Unerwienie współczulne i przywspółczulne naczyń mózgowych ma znaczenie
drugorzędne w regulacji przepływu krwi.
Krążenie płynu mózgowo-rdzeniowego
W obrębie splotów naczyniówkowych (plexus chorioideus) komór
mózgowych bocznych, komory trzeciej i komory czwartej oraz wokół
naczyń włosowatych w tkance mózgowej tworzy się płyn mózgowo-rdzeniowy (cerebrospinal fluid). Płyn powstający wokół naczyń włosowatych odpływa wzdłuż przestrzeni Virchowa-Robina wokół naczyń
krwionośnych jamy podpajęczynowkowej. Płyn wytworzony w komorach
-Opona
twarda
-Kora mózgu
Astrocyty
wytw arzające bfonę
graniczną
glejową
Ryc. 203. Budowa schematyczna opon mózgowo-rdzeniowych i jamy podpajęczynowkowej
wypełnionej płynem mózgowo-rdzeniowym. (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
394
Krążenie
mózgowych odpływa również do jamy podpajęczynówkowej, skąd jest
wchłaniany do zatok żylnych w obrębie ziarnistości pajęczynówki (Pacchioniego).
Płyn mózgowo-rdzeniowy jest izotoniczny w stosunku do osocza,
różni się jednak pod względem zawartości niektórych składników. Zawiera nieznaczne ilości białek i cholesterolu oraz mniej jonów wapnia,
glukozy i kwasu moczowego. Bierze udział w wymianie składników
pomiędzy tkanką mózgową i krwią oraz spełnia funkcję amortyzatora dla
całego mózgowia.
Bariera mózgowa
Naczynia włosowate w mózgowiu są dodatkowo otoczone wypustkami komórek neurogleju. Są to astrocyty, które za pomocą wypustek
szczelnie otaczają od zewnątrz naczynia włosowate. Dzięki temu astrocyty tworzą dodatkową warstwę, którą muszą pokonać związki chemiczne
krążące we krwi. Wiele związków pokonuje barierę mózgową (blood-brain
barrier) dość szybko, inne wolno, jeszcze inne bardzo wolno. Woda,
dwutlenek węgla, tlen i glukoza przenikają przez barierę szybko. Większość jonów przechodzi przez barierę mózgową znacznie wolniej, a niektóre związki, jak np. aminy katecholowe (adrenalina, noradrenalina)
przenikają przez barierę w minimalnych ilościach. Bariera mózgowa
spełnia funkcję ochronną. Zabezpiecza tkankę mózgową przed wahaniami w stężeniu poszczególnych składników, występującymi w osoczu krwi,
oraz przed związkami szkodliwymi krążącymi we krwi.
I Krążenie wrotne
W obrębie jamy brzusznej krew po przepłynięciu przez pierwotną sieć
naczyń włosowatych żołądka, dwunastnicy, jelita cienkiego, jelita grubego,
trzustki i śledziony wpada do żyły wrotnej (yena portae), poprzez którą
dostaje się do wątroby i tam przepływa przez wtórną sieć naczyń
włosowatych. Z wątroby krew kieruje się do żyły głównej dolnej (yena cava
inferior) przez żyły wątrobowe (venae hepaticae).
W spoczynku, w pozycji leżącej przez wątrobę przepływa około 1,5 L
krwi na minutę, czyli około 28% pojemności minutowej lewej komory
serca (cardiac output). Przeciętnie 4/5 krwi dopływa do wątroby przez żyłę
Krążenie wrotne
395
Ż. główna dolna
ŚLEDZIONA
JELITO GRUBE
cz.
WSTĘPUJĄCA
OKRĘŻNICY
JELITO
GRUBE cz.
ZSTĘPUJĄCA
OKRĘŻNICY
OKRĘŻNICA
ESOWATA
ODBYTNICA
Ryc. 204. Krążenie wrotne. Krew dopływająca przez naczynia tętnicze do żołądka,
pęcherzyka żółciowego, jelita cienkiego i grubego i odpływająca przez naczynia żylne i żyłę
wrotną do wątroby.
wrotną i 1/i przez tętnicę wątrobową (arteria hepatica). Stosunki te zmieniają się w zależności do fazy trawienia pokarmów w przewodzie pokarmowym, od pozycji ciała i aktywności fizycznej.
W pozycji leżącej ciśnienie w żyle wrotnej wynosi od 0,9 do 1,3 kPa,
w żyłach wątrobowych zaś około 0,6 kPa. Przepływ krwi przez wątrobę
zwiększa się w czasie trawienia pokarmów w przewodzie pokarmowym,
zmniejsza się zaś w pozycji wyprostnej. W czasie pracy fizycznej przepływ krwi przez wątrobę znacznie się zmniejsza — aż do 350 raL na minutę.
Oddychanie
Istotą procesu oddychania jest wyzwolenie energii zgromadzonej w organizmie. Do wyzwolenia energii ze związków chemicznych w organizmie
człowieka niezbędny jest tlen atmosferyczny. Proces ten, czyli oddychanie,
dzieli się na:
• oddychanie zewnętrzne (external respiration) — polegające na do
prowadzeniu cząsteczek tlenu atmosferycznego do wnętrza komórek;
• oddychanie wewnętrzne (internal respiration), czyli wewnątrzkomórko
we — w czasie którego cząsteczki tlenu wchodzą w reakcje chemiczne.
I Oddychanie zewnętrzne
Oddychanie zewnętrzne jest procesem złożonym, w którym biorą udział:
układ oddechowy, składający się z dróg oddechowych i płuc, mięśnie
poprzecznie prążkowane szkieletowe, krew i układ sercowo-naczyniowy
oraz ośrodki nerwowe sterujące oddychaniem.
Oddychanie zewnętrzne polega na doprowadzeniu tlenu atmosferycznego do komórek zgodnie z gradientem ciśnienia parcjalnego tlenu.
Jednocześnie zostaje usuwany z komórek dwutlenek węgla powstający
w wyniku utleniania komórkowego związków organicznych. Dwutlenek
węgla jest usuwany również zgodnie z gradientem ciśnienia parcjalnego.
Oddychanie zewnętrzne dzieli się na szereg procesów:
•
•
•
•
wentylacja płuc;
dyfuzja gazów pomiędzy powietrzem pęcherzykowym i krwią;
transport gazów za pośrednictwem krwi;
dyfuzja gazów pomiędzy krwią i komórkami.
Oddychanie zewnętrzne
397
I Wentylacja płuc
Wdechy i wydechy
W czasie wentylacji płuc (pulmonary ventilation) do pęcherzyków
płucnych {aheoli pulmonis) jest wciągane powietrze atmosferyczne zawierające tlen i inne gazy oraz bardzo mało dwutlenku węgla. Powietrze
wydychane zawiera mniej tlenu, a zawartość dwutlenku węgla jest w nim
znacznie większa niż w powietrzu wdychanym.
Wentylacja płuc jest uzależniona od ruchów oddechowych klatki piersiowej. Polegają one na wdechu i wydechu. W czasie wdechu powiększa się
objętość klatki piersiowej. Skurcz mięśni wdechowych, którymi są: przepona (diaphragma) i mięśnie międzyżebrowe zewnętrzne {musculi intercostales externi), powoduje powiększenie wymiarów wewnętrznych klatki
piersiowej: pionowego, strzałkowego i czołowego.
Opłucna płucna (pleura pulmonalis) przylega do opłucnej ściennej
{pleura parietalis) i w czasie wdechu podąża za nią, wypełniając całą
jamę opłucnej (cavum pleurale), w której panuje ujemne ciśnienie w czasie
spokojnego oddychania od 0,3 do 0,8 kPa. Powoduje to rozciągnięcie
tkanki płucnej, obniżenie się ciśnienia w pęcherzykach płucnych, w dro-
Ryc. 205. Zmiany ciśnienia w płucach i w jamie opłucnej w czasie wdechu i wydechu.
398
Oddychanie
gach oddechowych i napływ powietrza do płuc w celu wyrównania
powstałej różnicy ciśnień.
Na szczycie wdechu mięśnie wdechowe rozkurczają się i klatka piersiowa zaczyna zmniejszać swoją objętość dzięki sile wywieranej przez
rozciągnięte elementy sprężyste w tkance płucnej. Ciśnienie w pęcherzykach płucnych wzrasta powyżej ciśnienia atmosferycznego i powietrze jest
usuwane na zewnątrz.
Spokojny wydech jest aktem biernym, nie wymagającym skurczu
mięśni, natomiast w czasie nasilowego wydechu kurczą się mięśnie międzyżebrowe wewnętrzne {musculi intercostales interni) oraz mięśnie przedniej ściany jamy brzusznej, przede wszystkim mięśnie proste brzucha
{muscli recti abdominis).
W nasilonych wdechach biorą udział mięśnie wdechowe dodatkowe:
mięśnie mostkowo-obojczykowo-sutkowe {musculi sternocleidomastoidei),
mięśnie piersiowe mniejsze (musculi pectorales minores), mięśnie zębate
przednie (musculi serrati anteriores), mięśnie czworoboczne (musculi
trapezii), mięśnie dźwigacze łopatki (musculi levatores scapulae), mięśnie
równoległoboczne większe i mniejsze (musculi rhomboidei maiores et
minores) i mięśnie pochyłe (musculi scaleni).
W okresie spoczynku klatka piersiowa wykonuje rytmiczne ruchy
wdechowe i wydechowe o częstotliwości 0,26 Hz, czyli średnio 16
oddechów na minutę.
Pojemność płuc
U mężczyzn w płucach na szczycie najgłębszego wdechu znajduje się
około 6 L powietrza. Jest to pojemność płuc całkowita — TLC (total lung
capacity). Pojemność płuc całkowita dzieli się na pojemność wdechową —
IC (inspiratory capacity) — i pojemność zalegającą czynnościową — FRC
(functional residual capacity).
Pojemność wdechową (IC) stanowi powietrze wciągane do płuc w czasie
najgłębszego wdechu po spokojnym wydechu. Pojemność zalegająca
czynnościowa (FRC) jest to powietrze pozostające w płucach po spokojnym wydechu. Każda z tych dwóch pojemności (IC i FRC) dzieli się z kolei
na dwie objętości.
Objętość oddechowa — TV (tidal volume) — wdychana i wydychana
w czasie swobodnego wdechu i wydechu, oraz objętość zapasowa wdechowa — IRV (inspiratory reserve volume) — wciągana do płuc w czasie
maksymalnego wdechu wykonywanego na szczycie swobodnego wdechu,
tworzą łącznie pojemność wdechową (IC).
Oddychanie zewnętrzne
399
Maksymalna
pojemność
wdechowa
Spoczynkowa
objętość
wydechowa
Maksymalna
pojemność
wydechowa
Ryc. 206. Pojemność płuc całkowita (TLC): IC — pojemność wdechowa, FRC — pojemność zalegająca czynnościowa, TV — objętość oddechowa, IRV — objętość zapasowa
wdechowa, ERV — objętość zapasowa wydechowa, RV — objętość zalegająca, VC — pojemność życiowa.
Po swobodnym wdechu można wykonać maksymalny wydech, usuwając z płuc objętość zapasową wydechową — ERV (expiratory reserve
volume). W czasie maksymalnego wydechu zawsze pozostaje w płucach
objętość zalegająca — RV (residual volume). Objętość zapasowa wydechowa i objętość zalegająca łącznie tworzą pojemność zalegającą czynnościową
(FRC).
Po najgłębszym wydechu wykonując maksymalny wdech, wciąga się do
płuc powietrze stanowiące pojemność życiową wdechową — IVC (inspiratory vital capacity). Jest ona nieco większa od pojemności życiowej —
VC (vital capacity), a więc od ilości powietrza, które można usunąć z płuc
po maksymalnym wdechu w czasie maksymalnego wydechu. Wykonując
maksymalny wydech poprzedzony maksymalnym wdechem, przewodziki
pęcherzykowe zamykają się wcześniej, zanim powietrze wypełniające
pęcherzyki zostanie usunięte. Z tego względu pojemność życiowa wdecho-
400
Oddychanie
wa (IVC) może być większa od pojemności życiowej (VC) mierzonej
w czasie wydechu.
Objętość zalegająca (RV) obejmuje powietrze znajdujące się w pęcherzykach płucnych (afoeolipulmonis) i przewodzikach pęcherzykowych {ductuli
aheolares), a więc tam, gdzie istnieją anatomiczne warunki do wymiany
gazów.
Ryc. 207. Zapis objętości powietrza wdychanego i wydychanego (spirogram) pr2y użyciu
aparatu (spirografu) do zapisywania objętości powietrza wdychanego i wydychanego. Zapis
objętości wdechów i wydechów został wykonany w czasie oddychania spoczynkowego
i w czasie maksymalnej dowolnej wentylacji płuc (MW).
W czasie swobodnego wdechu wprowadzane jest do dróg oddechowych
około 500 mL powietrza stanowiące objętość oddechową (TV). Z tej
objętości powietrza wdychanego do pęcherzyków płucnych dostaje się
około 350 mL, a pozostałe 150 mL wypełnia przestrzeń martwą anatomiczną (anatomical respiratory dead space). Przestrzeń tę tworzą drogi
oddechowe, w których nie ma warunków anatomicznych do wymiany
gazów pomiędzy powietrzem a krwią: jama nosowa (cavum nasi), gardło
(pharynx), krtań (Iarynx), tchawica (trachea), oskrzela (bronćhi) i oskrzeliki {bronchioli).
Oddychanie zewnętrzne
401
Część powietrza wdychanego może dostawać się do pęcherzyków
płucnych i nie podlegać wymianie gazowej. W naczyniach włosowatych
w ścianie tych pęcherzyków krew nie przepływa lub przepływa jej zbyt
mało, aby całe wprowadzone powietrze mogło być wykorzystane do
wymiany gazowej. Powietrze pęcherzykowe nie podlegające wymianie
stanowi tzw. przestrzeń martwą fizjologiczną (physiological respiratory
dead space). U ludzi zdrowych przestrzeń martwa fizjologiczna jest mała,
natomiast znacznie zwiększa się u chorych.
W czasie spoczynku jest wdychane i wydychane około 8 L powietrza na
minutę — 16 oddechów x 500 mL powietrza objętości oddechowej. Jest to
wentylacja płuc minutowa (pulmonary ventilation). Może ona znacznie
zwiększyć się w czasie wykonywania szybkich i głębokich oddechów.
Maksymalna dowolna wentylacja płuc — MW (maximal voluntary
ventilation) — może być od kilkunastu do dwudziestu kilku razy większa
od wentylacji płuc minutowej w czasie spoczynku.
W celach diagnostycznych do badania sprawności układu oddechowego stosuje się pomiar objętości powietrza wydychanego w czasie
pierwszej sekundy po najgłębszym wdechu, czyli natężonej objętości
wydechowej w pierwszej sekundzie — FEV, 0 (forced expiratory volume in
the first sec).
Ryc. 208. Zapis objętości powietrza wdychanego i wydychanego w czasie oddychania
w spoczynku i w czasie pomiaru natężonej objętości wydechowej w pierwszej sekundzie
(FEV, 0). W czasie wykonania pomiarów FEV, 0 przesuw taśmy rejestrującej spirografu
przyspieszono dziesięciokrotnie z 1 mm na sekundę do 10 mm na sekundę.
402
Oddychanie
Wentylacja pęcherzyków
Suche powietrze atmosferyczne, bez pary wodnej, wciągane do płuc
zawiera 21% tlenu, 78% azotu i 0,04% dwutlenku węgla (patrz. tab. 30).
Powietrze wciągane do płuc, stanowiące objętość oddechową (TV), miesza się z powietrzem stanowiącym pojemność zalegającą czynnościową
(FRC), jednocześnie ogrzewa się i wysyca parą wodną.
W czasie każdego wdechu do pęcherzyków płucnych dostaje się tylko
około 350 mL powietrza oddechowego. Pozostała część objętości oddechowej, stanowiąca około 150 mL powietrza, wypełnia przestrzeń
martwą anatomiczną. Ostatecznie powietrze pęcherzykowe ma średnie
ciśnienie parcjalne gazów *:
PO2= 13,3 kPa, PN = 76,4 kPa, PH2o = 6,3 kPa i PCO2 = 5,3 kPa.
Powietrze o takim składzie styka się ze ścianą pęcherzyków płucnych
i przewodzików pęcherzykowych, których łączna powierzchnia wynosi
około 70 m2. Jest to powierzchnia, przez którą dyfundują gazy.
Ryc. 209. Schemat budowy pneumocytu typu II — komórki nabłonka oddechowego
pęcherzyków płucnych wydzielającej czynnik powierzchniowy (surfaktant). (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL,
Warszawa 1997.)
Prężność gazów — w cieczach, ciśnienie parcjalne — w powietrzu.
Oddychanie zewnętrzne
403
Tkanka płucna jest elastyczna, co wiąże się z:
• elementami sprężystymi występującymi pomiędzy pęcherzykami;
• napięciem powierzchniowym cienkiej warstwy płynu wyściełającego
wnętrze pęcherzyków płucnych.
W czasie wdechu napięcie powierzchniowe w pęcherzykach wzmaga się,
a w czasie wydechu maleje. Zapobiega to zlepianiu się ścian pęcherzyków
na szczycie wydechu. Napięcie powierzchniowe w pęcherzykach zmniejsza
czynnik powierzchniowy, czyli surfaktant (surfactant). Czynnik ten stanowią cząsteczki lipoprotein (wśród których najwięcej jest dipalmitoilo-fosfatydylocholiny) wydzielane przez ziarniste pneumocyty, czyli komórki pęcherzyka oddechowego duże. W czasie wdechu powierzchnia pęcherzyków zwiększa się, cząsteczki czynnika powierzchniowego ulegają
rozsunięciu i napięcie powierzchniowe się wzmaga. Zjawiska przeciwne
zachodzą w czasie wydechu.
I Dyfuzja gazów w płucach
W pęcherzykach płucnych zachodzi wymiana gazów pomiędzy powietrzem i krwią przepływającą przez sień naczyń włosowatych otaczających
pęcherzyki. W tych naczyniach stale znajduje się około 100 mL krwi. Ilość
ta przepływa przez naczynia włosowate w czasie około 0,8 s. Podczas pracy
fizycznej i związanego z nią zwiększenia pojemności minutowej serca krew
przepływa znacznie szybciej przez naczynia włosowate pęcherzyków
płucnych.
Dyfuzja gazów przez ścianę pęcherzyków odbywa się zgodnie z gradientem prężności cząsteczek gazów. Cząsteczki tlenu dyfundują ze światła
pęcherzyków do krwi, ponieważ w powietrzu pęcherzykowym ciśnienie
parcjalne tlenu jest większe, we krwi dopływającej ze zbiornika tętniczego
płucnego zaś jest mniejsze. W przeciwnym kierunku dyfundują cząsteczki
dwutlenku węgla. We krwi dopływającej do naczyń włosowatych pęcherzyków PCQ2 jest większe, w powietrzu pęcherzykowym zaś PCo2 jest mniejsze
(patrz tab. 30, str. 408).
Częsteczki O2, dyfundując do krwi, muszą pokonać ścianę pęcherzyka
płucnego i ścianę naczynia włosowatego. Grubość tej przegrody nie
przekracza 1 um i — poczynając od światła pęcherzyka płucnego —
jest utworzona przez: warstwę płynu pokrywającą powierzchnię pęcherzyków, nabłonek pęcherzyków, błonę podstawną i śródbłonek naczyń
włosowatych. Cząsteczki O 2 po przejściu przez tę przegrodę rozpusz-
404
Oddychanie
Ryc. 210. Budowa schematyczna ściany pęcherzyka płucnego oddzielającej powietrze
pęcherzykowe od krwi w naczyniach włosowatych. Cząsteczki tlenu ze światła pęcherzyków
płucnych dyfundują przez ich ścianę do osocza w naczyniach włosowatych i z osocza do
erytrocytów. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Ryc. 211. Ciśnienie parcjalne tlenu i dwutlenku węgla w pęcherzykach płucnych w górnych,
środkowych i dolnych partiach płuc.
Oddychanie zewnętrzne
405
czają się w osoczu wypełniającym naczynia włosowate na zasadzie
rozpuszczalności fizycznej. Z osocza O2 natychmiast dyfunduje do erytrocytów.
Cząsteczki CO2 dyfundują z osocza krwi przepływającej przez naczynia
włosowate do światła pęcherzyków, tj. w kierunku przeciwnym niż
cząsteczki O2.
Wartości średnie Po2 i Pco2 występują w powietrzu pęcherzykowym
wypełniającym środkowe partie płuc. W górnych partiach płuc powietrze
pęcherzykowe ma wyższe PO2 i niższe PCo2 niż wartości średnie. Przeciwnie, w
dolnych partiach płuc powietrze pęcherzykowe ma niższe Po2 i wyższe
P Co2- Różnice w ciśnieniu parcjalnym w powietrzu pęcherzykowym O 2 i
CO2 zależą od ilości krwi przepływającej przez sieć naczyń włosowatych
w górnych, środkowych i dolnych partiach płuc.
I Transport gazów
Cząsteczki O2 dyfundujące do osocza krwi w niewielkiej ilości rozpuszczają się w nim na drodze fizycznej. Przy ciśnieniu parcjalnym
tlenu w pęcherzykach = 13,3 kPa w 1 L krwi odpływającej do zbiornika
żylnego płucnego tylko 3 mL O2 jest rozpuszczone na zasadzie rozpuszczalności fizycznej. Jest to ilość niedostateczna do zapewnienia oddychania
wewnętrznego na minimalnym poziomie. Podobnie w tkankach — przy
prężności dwutlenku węgla równej 6,1 kPa na drodze fizycznej rozpuszcza
się w 1 L krwi tylko 35 mL CO 2 (patrz tab. 30).
Zarówno cząsteczki O 2, jak i CO2 wiążą się we krwi na drodze
chemicznej i w tej postaci są przede wszystkim transportowane zgodnie
z gradientem prężności.
Transport tlenu
Cząsteczki O2 rozpuszczone w osoczu na drodze fizycznej dyfundują
przez otoczkę do erytrocytów i wiążą się z hemoglobiną, tworząc hemoglobinę utlenowaną, czyli oksyhemoglobinę (oxygen-hemoglobin). Jedna cząsteczka hemoglobiny Hb4 wiąże się z czterema cząsteczkami tlenu,
tworząc hemoglobinę utlenowaną:
406
Oddychanie
Dzięki występowaniu hemoglobiny zdolność krwi do transportu tlenu
wzrasta około 70 razy. W powietrzu pęcherzykowym w warunkach
P02= 13,3 kPa w 1 L krwi odpływającej z naczyń włosowatych pęcherzyków płucnych jest rozpuszczone na drodze fizycznej 3 mL O 2, a około
190 mL O2 jest związane z hemoglobiną.
Krew w zbiorniku żylnym płucnym ma Po2= 12,7 kPa i hemoglobina
jest wysycona tlenem w 97%. Mniejsza PO2 w zbiorniku żylnym płucnym
w stosunku do P02 w naczyniach włosowatych pęcherzyków płucnych
związane jest z przepływem pewnej ilości krwi ze zbiornika tętniczego
dużego do zbiornika żylnego płucnego przez naczynia krwionośne oskrzeli
z pominięciem pęcherzyków płucnych, czyli z przeciekiem płucnym krwi
(pulmonary shunt).
Krew zawierająca hemoglobinę wysycona tlenem (hemoglobin saturated) odpływa z płuc, kierując się przez zbiornik żylny płucny, lew y
Pęcherzyk płucny
I
Ryc. 212. Transport gazów pomiędzy
powietrzem pęcherzykowym i
tkankami
Po2= 13,3 kPa (100 mm Hg)
Pco 2 = 5,3 kPa(40mm Hg)
0,
Krew
tętnicza
Krew
żytna
Tkanki
Po 2 =4,7kPa (35mmHg)
Pco2=6,1 kPa(4B mmHg)
Oddychanie zewnętrzne
407
przedsionek serca, lewą komorę serca, zbiornik tętniczy duży do sieci
naczyń włosowatych krążenia dużego. W naczyniach włosowatych w tkankach prężność tlenu jest mała i z hemoglobiny utlenowanej znajdującej
się w erytrocytach uwalnia się około 7 4 transportowanego tlenu. Krew
żylna odpływająca do zbiornika żylnego dużego zawiera hemoglobinę
wysyconą tlenem średnio w 75% i Po2 = 5,3 kPa.
Zależność pomiędzy ciśnieniem parcjalnym tlenu a stopniem wy sycenia
hemoglobiny tlenem przedstawia krzywa dysocjacji hemoglobiny utlenowanej. Krzywa ta przesuwa się w lewo i staje się bardziej stroma w miarę
obniżania się temperatury, zwiększania wartości pH i zmniejszenia się
prężności dwutlenku węgla we krwi. Warunki, w których krzywa dysocjacji hemoglobiny utlenowanej przesuwa się w lewo i staje się bardziej
stroma, panują w naczyniach włosowatych pęcherzyków płucnych. Przy
tym samym Po2 więcej hemoglobiny wiąże się z tlenem. Przeciwnie —
w sieci naczyń włosowatych krążenia dużego panują warunki przesuwające w prawo i spłaszczające krzywą dysocjacji hemoglobiny utlenowanej,
które sprzyjają uwalnianiu się cząsteczek O 2 ze związku z hemoglobiną
(patrz ryc. 152, str. 302).
Transport dwutlenku węgla
We krwi przepływającej przez sieć naczyń włosowatych krążenia dużego
zawar toś ć dwut lenku węgla zwięks za się o 5 0 mL CO 2 /L kr wi
i jego prężność zwiększa się o 0,8 kPa. Krew tętnicza w zbiorniku tętniczym
dużym zawiera około 470 mL CO2/L krwi i PCo2 = 5,3 kPa, a w zbiorniku
żylnym dużym krew zawiera około 520 mL CO2/L krwi i Pco2 = 6,l kPa
(patrz tab. 30).
Dwutlenek węgla dyfundujący z tkanek do krwi przepływającej przez
naczynia włosowate jest transportowany do płuc:
• około 6% w postaci CO2 rozpuszczonego na zasadzie rozpuszczalności
fizycznej w osoczu i w cytoplazmie erytrocytów;
• około 88% w postaci jonów HCO3 związanych przez wodorowęglanowy układ buforowy osocza i erytrocytów;
• około 6% w postaci karbaminianów, CO 2 związanego z wolnymi
grupami aminowymi białek osocza i hemoglobiny.
Cząsteczki CO2 dyfundujące z tkanek do krwi rozpuszczają się w osoczu na zasadzie rozpuszczalności fizycznej i przenikają w tej postaci do
wnętrza erytrocytów. Tam pod wpływem enzymu anhydrazy węglanowej
408
Oddychanie
(carbonic anhydrase) dwutlenek węgla wiąże się z wodą i powstaje kwa
węglowy:
CO2 + H2O *± H2CO3 <=> H+ + HCO;
Kwas węglowy dysocjuje na wolne jony H+ i HCO]. Jony H+ wiążą się
z hemoglobiną, większość jonów HCO3 dyfunduje zaś do osocza.
Zwiększenie stężenia jonów HCO3 w osoczu krwi żylnej i zmniejszenie
we krwi tętniczej powoduje wędrówkę jonów Cl~ przez otoczkę erytrocytów. We krwi żylnej jony HCO~ przechodzą z erytrocytów do osocza,
jony Cl~ wchodzą zaś do ich wnętrza. We krwi tętniczej jony C\~ wychodzą z erytrocytów do osocza.
Tabela 30
Ciśnienie parcjalne O2 i CO2 w powietrzu oraz prężność O2 i CO2 we krwi i w tkankach
P
o2
Pco2
SI: kPa
mm Hg
SI: kPa
mm Hg
Powietrze atmosferyczne
Powietrze pęcherzykowe
Krew w naczyniach włosowatych pęcherzyków płucnych
21,3
13,3
160
100
0,04
5,3
0,3
40
13,3
100
5,3
40
Krew tętnicza
Krew żylna
12,7
5,3
95
40
5,3
6,1
40
46
Tkanki
4,7
35
6,1
46
Dwutlenek węgla rozpuszczony w osoczu i znajdujący się w erytrocytach wiąże się z grupami aminowymi aminokwasów, z których są
zbudowane białka osocza i hemoglobina. W wyniku reakcji:
, + R -N H, - R -NH CO OH
tworzą się karbaminiany. Większość karbaminianów powstaje w eryt rocytach po połączeniu się CO 2 z grupami aminowymi hemoglobiny.
Oddychanie zewnętrzne
409
I Dyfuzja gazów w tkankach
Krew tętnicza dopływająca do wszystkich tkanek ma większą prężność
tlenu i mniejszą prężność dwutlenku węgla w porównaniu z odpływającą
krwią żylną. Zgodnie z gradientem prężności uwolniony z hemoglobiny
tlen dyfunduje do komórek, dwutlenek węgla zaś dyfunduje w kierunku
przeciwnym z komórek do osocza.
Cząsteczki O2 uwolnione z hemoglobiny przechodzą przez otoczkę
erytrocytów do osocza, następnie przez komórki śródbłonka naczyń
włosowatych do płynu międzykomórkowego i dopiero z tego płynu
dyfundują przez błonę komórkową do poszczególnych komórek.
W zależności od intensywności metabolizmu wewnątrzkomórkowego
występują dość znaczne różnice w "prężności tlenu w poszczególnych
tkankach. W tkankach o intensywnym metabolizmie prężność tlenu
w komórkach jest mała i jednocześnie te tkanki zużywają więcej tlenu.
Krew żylna odpływająca z tkanek o intensywnym metabolizmie zawiera
mniej tlenu i więcej dwutlenku węgla.
Tabela 31
Przepływ krwi i zużycie tlenu przez narządy i tkanki
Przepływ krwi
Różnica tętniczo-Tkanka
(mL krwi/kg
żylna zawartości
tkanki/s)
tlenu (mL O2/L
krwi)
Zużycie tlenu
(mL O2/kg
tkanki/s)
Mięsień sercowy
Mięśnie szkieletowe
(w spoczynku)
14,00
0,45
114,0
60,0
1,62
0,03
Nerki
70,00
Wątroba
9,62
14,0
34,0
1,00
0,33
Mózg
9,0
62,0
0,55
Stopień zużycia tlenu (oxygen consumption) przez poszczególne tkanki
wyraża się różnicą tętniczo-żylną w zawartości tlenu (arteriovenous oxygen
difference). Krew tętnicza dopływająca do wszystkich tkanek ma jednakową zawartość tlenu, natomiast krew żylna może zawierać go więcej
lub mniej.
410
Oddychanie
W spoczynku pojemność minutowa serca wynosi około 5,4 L, a różnica tętniczo-żylna w zawartości tlenu we krwi pomiędzy zbiornikiem
tętniczym dużym a zbiornikiem żylnym dużym wynosi około 46 mL O 2 na
1 L krwi. Na tej podstawie można obliczyć zużycie tlenu przez cały
organizm:
5,4 L krwi x 46 mL O 2/L krwi = 248 mL O2/l min.
W okresie aktywności ruchowej, a zwłaszcza w czasie pracy fizycznej,
zwiększa się znacznie pojemność minutowa serca i wentylacja minutowa
płuc. Mimo to zużycie tlenu przez pracujące mięśnie szkieletowe jest
tak duże, że zwiększa się różnica tętniczo-żylna w zawartości tlenu we
krwi pomiędzy zbiornikiem tętniczym dużym a zbiornikiem żylnym
dużym. Zużycie spoczynkowe tlenu na minutę może zwiększyć się do 16
razy w czasie wysiłku fizycznego.
I Regulacja oddychania
I
Ośrodek oddechowy
Regulacja oddychania, czyli częstotliwość i głębokość oddechów,
odbywa się za pośrednictwem ośrodka oddechowego (respiratory center)
w rdzeniu przedłużonym (medulla oblongata). W skład tego ośrodka
wchodzą dwa rodzaje neuronów tworzące dwa ośrodki o przeciwnej
funkcji. Ich umiejscowienie częściowo pokrywa się. Oba rodzaje neuronów należą do tworu siatkowatego rdzenia przedłużonego (formatio
reticularis medullae oblongatae). Są to neurony wdechowe tworzące ośrodek
wdechu (inspiratory center), znajdujące się w jądrze samotnym (nucleus
solitarius) i w części przedniej jądra tylno-dwuznacznego nerwu błędnego
{pars anterior nucleus retroambigualis nervi vagi) oraz ośrodek wydechu
(expiratory center) w jądrze dwuznacznym nerwu błędnego (nucleus
ambiguus nervi vagi) i w części tylnej jądra tylno-dwuznacznego nerwu
błędnego (pars posterior nucleus retroambigualis nervi vagi).
Ośrodek wdechu wysyła impulsy nerwowe do rdzenia kręgowego, do
neuronów ruchowych unerwiających mięśnie wdechowe, ośrodek wydechu pobudza zaś neurony ruchowe unerwiające mięśnie wydechowe.
Neurony ośrodka wdechu stanowią rozrusznik dla czynności oddechowej. Kilkanaście razy na minutę, średnio 16 razy, neurony ośrodka wdechu pobudzają się i wysyłają salwę impulsów nerwowych. Impulsy od
neuronów wdechowych biegną przez gałązkę zstępującą aksonu do
Regulacja oddychania
411
Jądro
Jądro
tylnodwuzraczne
część
przednia
Jądro
łylnodwuznaczne
część
przednia
samotne
Ryc. 213. Schemat budowy rdzenia przedłużonego z zaznaczonym umiejscowieniem neuronów wdechowych: a — w jądrze samotnym; b — w jądrze tylno-dwuznacznym w części
przedniej (pola zakropkowane na czerwono). (Nieznacznie zmodyfikowano i reprodukowano za zezwoleniem z E. G. Merrill Where are the real respiratory neurones? Fed. Proc.
Fed. Am. Soc. Exp. Biol., 1981, 40, 2389—2394.)
Jądro
dwuznaczne
Jądro
tylno-
dwuznaczne
część
tylna
Ryc. 214. Schemat budowy rdzenia przedłużonego z zaznaczonym umiejscowieniem neuronów wydechowych w jądrze dwuznacznym
i w jądrze tylno-dwuznacznym w części tylnej (pola zakreskowanego poziomymi czerwonymi kreskami). (Nieznacznie zmodyfikowano i reprodukowano za zezwoleniem
z E. G. Merrill Where are the real respiratory
neurones? Fed. Proc. Fed. Am. Soc. Exp. Biol.,
1981, 40, 2389—2394.)
412
Oddychanie
neuronów ruchowych w rdzeniu kręgowym i jednocześnie przez gałązkę
wstępującą aksonu do neuronów tworu siatkowatego mostu (pons), tworzących ośrodek pneumotaksyczny (pneumotaxic center). Ośrodek pneumotaksyczny hamuje zwrotnie ośrodek wdechu na 1—2 s, po czym neurony ośrodka wdechu ponownie pobudzają się i wysyłają salwę impulsów
do rdzenia kręgowego. Rytmiczność oddechów związana jest z występującymi po sobie kolejno okresami pobudzania i hamowania ośrodka
wdechu.
Modulacja aktywności ośrodka wdechu
Pobudzenie powstające samoistnie w ośrodku wdechu jest modulowane, a więc oddechy przyspieszają się i są pogłębione lub zwalniają się
i spłycają na skutek:
• impulsów wysyłanych przez receptory i odbieranych przez neurony
wdechowe;
• zmiany wartości pH w bezpośrednim sąsiedztwie neuronów wdecho
wych, czyli po podrażnieniu chemodetektorów.
Impulsy nerwowe modulujące aktywność neuronów ośrodka wdechu
biegną od:
• chemoreceptorów kłębuszków szyjnych (glomus caroticum) i kłębków
aortowych (glomus aorticum);
• interoreceptorów w tkance płucnej oraz proprioreceptorów klatki
piersiowej;
• ośrodków znajdujących się w wyższych piętrach mózgowia: z kory
mózgu, układu limbicznego i ośrodka termoregulacji w podwzgórzu.
Chemoreceptory
Zasadniczym modulatorem aktywności ośrodka wdechu są impulsy
aferentne biegnące od chemoreceptorów kłębuszków szyjnych i kłębków
aortowych. Przez kłębki stale przepływają duże, w stosunku do niewielkiej ich masy, ilości krwi tętniczej. Bodźcem drażniącym chemoreceptory
jest nieznaczne zwiększenie PCo2 i koncentracji jonów wodorowych lub
znaczne zmniejszenie Po2 we krwi tętniczej.
Impulsacja aferentna jest przewodzona od kłębków aortowych do
rdzenia przedłużonego za pośrednictwem włókien biegnących w nerwie
Regulacja oddychania
Nerw błędny
41 3
Nerw biedny
Tętnico szyjna
wewnętrzna prawa
Tętnica szyjna
wewnętrzna lewa
Zatoka szyjna
tętnicza
Klębuszek szyjny
Zatoka szyjna
tętnicza
Ktębuszek szyjny
Tętnica szyjna
wspólna
Tętnica
podobojczykowa
prawa
Tętnica
podobojczykowa
lewa
Kłębek aortowy
Ryc. 215. Chemoreceptory w kłębuszkach szyjnych i w kłębkach aortowych i ich uner wienie.
językowo-gardłowym (n. IX) i w nerwie błędnym (n. X). Impulsacja
wysyłana przez podrażnione chemoreceptory pobudza ośrodek wdechu
i oddechy stają się przyspieszone i pogłębione.
Pod wpływem impulsacji współczulnej komórki chromochłonne kłębuszka szyjnego uwalniają dopaminę, która zmniejsza pobudliwość chemoreceptorów stanowiących zakończenia gałązki nerwu językowo-gardłowego. Przez zmianę pobudliwości chemoreceptorów dochodzi do zmiany
aferentnej impulsacji biegnącej do ośrodka oddechowego w rdzeniu
przedłużonym.
Interoreceptory i proprioreceptory
Rozciągnięcie tkanki płucnej pobudza interoreceptory — mechanoreceptory inflacyjne (inflation receptors) — znajdujące się pomiędzy
mięśniami gładkimi oskrzeli i wyzwala wydech. Przeciwnie — zmniejszę-
414
Oddychanie
nie stopnia rozciągnięcia płuc w czasie wydechu pobudza inne mechanoreceptory deflacyjne (deflation receptors) i wyzwala wdech. Są to odruchy
Heringa-Breuera. Podrażnione receptory pod nabłonkiem dróg oddecho-
wych wyzwalają odruch kaszlu, receptory J zaś, występujące pomiędzy
pneumocytami i naczyniami włosowatymi otaczającymi pęcherzyki, są
wrażliwe na pojawienie się płynu międzykomórkowego. Podrażnienie
płuc, któremu towarzyszy ich przekrwienie, pobudza te receptory i powoduje początkowy bezdech, po którym występują szybkie i płytkie oddechy. Impułsacja od interoreceptorów w płucach przewodzona jest do
rdzenia przedłużonego przez aferentne włókna nerwu błędnego.
Wdechowe lub wydechowe ustawienie klatki piersiowej drażni odpowiednie proprioreceptory i wpływa modulująco na częstość i głębokość
oddechów. Im głębszy jest wdech, tym głębszy wydech po nim następuje.
Chemodetektory w rdzeniu przedłużonym
W rdzeniu przedłużonym na powierzchni brzusznej znajdują się neurony wrażliwe na zmianę wartości pH płynu mózgowo-rdzeniowego. Zwięk-
szona dyfuzja CO 2 z krwi do płynu mózgowo-rdzeniowego powoduje
n.X
Ryc. 216. Schemat regulacji
oddychania.
Oddychanie wewnętrzne
41 5
zwiększenie w nim stężenia kwasu węglowego (H2CO3) i zwiększenie
koncentracji jonów wodorowych w bezpośrednim otoczeniu chemodetektorów. Zwiększenie koncentracji jonów wodorowych podrażnia chemodetektory, które z kolei pobudzają ośrodek wdechu.
Wrażliwość chemodetektorów na zmianę wartości pH zmniejsza się
w czasie snu oraz w czasie ogólnej narkozy.
I Od d yc h a n i e w ew n ętrzn e
I Przemiana pośrednia w komórkach
Cząsteczki O2 dyfundujące do wnętrza komórki wychwytywane są
przez mitochondria. W obrębie błony zewnętrznej mitochondriów znajdują się wszystkie enzymy cyklu kwasów trikarboksylowych, czyli cyklu
Krebsa, na błonie wewnętrznej zaś enzymy łańcucha oddechowego.
Metabolity heksoz, aminokwasów i kwasów tłuszczowych w cyklu
kwasów trikarboksylowych utleniane są do dwutlenku węgla i wody
z jednoczesnym uwolnieniem wolnych atomów wodoru lub wolnych
elektronów. Enzymy łańcucha oddechowego przenoszą atomy wodoru na
tlen. W procesie tym powstają cząsteczki wody.
Gromadzenie energii w komórce
Komórki nie czerpią bezpośrednio energii z utleniania produktów
energetycznych wprowadzonych do organizmu z zewnątrz. Związki, takie
jak węglowodany, aminokwasy i tłuszcze, utleniając się wyzwalają energię
do syntezy związków fosforowych wysokoenergetycznych. Wśród związków fosforowych wysokoenergetycznych najważniejszym, uniwersalnym
przenośnikiem energii jest adenozynotrifosforan — ATP. Adenozynotrifosforan rozszczepia się w komórce na drodze hydrolizy na adenozynodifosforan (ADP) i ortofosforan. Uwolnienie ortofosforanu wyzwala znaczne
ilości energii.
Komórki czerpią energię z ATP do biosyntezy związków, do aktywnego
transportu wewnątrzkomórkowego przez błony komórkowe przeciw gradientowi stężeń oraz do ruchu całej komórki.
Do resyntezy ATP z ADP i ortofosforanu dostarczana jest energia
z katabolizmu węglowodanów, białek i tłuszczów. Zwiększenie zawartości
w komórce ADP przyspiesza transport elektronów i atomów wodoru
416
Oddychanie
Ryc. 217. Cykl kwasów trikarboksylowych.
Pirogronian
Acetylo - koenzym A
Bursztynian
w jednostce czasu przez łańcuch oddechowy i przyspiesza przenoszenie ich
na tlen. Zwiększa się zużycie tlenu w komórce wraz z resynteza ATP.
Resynteza 3 cząsteczek ATP z ADP i ortofosforanu wiąże się z wytworzeniem 1 cząsteczki wody, do czego potrzebne jest pół cząsteczki
tlenu.
Glikoliza
Niezbędnym materiałem energetycznym zużywanym przez komórki
w organizmie jest glukoza. Poza rozpadem glukozy pewne ilości energii są
Oddychanie wewnętrzne
41 7
czerpane przez komórki z innych cukrów prostych oraz aminokwasów
i kwasów tłuszczowych.
Rozpad glukozy zachodzi w warunkach:
• beztlenowych w cytoplazmie — ostatecznym produktem jest mleczan;
• tlenowych w cytoplazmie i w mitochondriach — ostatecznymi produk
tami są dwutlenek węgla i woda.
W czasie glikolizy beztlenowej według schematu Embdena-Meyerhofa
w stadiach początkowych następuje zużycie energii czerpanej z ATP.
Natomiast w stadiach końcowych znacznie więcej ATP jest resyntetyzowane. W wyniku ostatecznym energia wyzwolona z rozpadu glukozy do
mleczanu przenosi się na ATP. Z rozpadu jednej cząsteczki glukozy
w warunkach glikolizy beztlenowej są resyntetyzowane netto 2 cząsteczki
ATP.
Glikoliza w warunkach tlenowych przebiega do pirogronianu. Pirogronian jest wprowadzany do cyklu kwasów trikarboksylowych przy
udziale acetylo-koenzymu A. Glikoliza w warunkach tlenowych jest
znacznie wydajniejsza energetycznie od glikolizy beztlenowej. Z rozpadu
jednej cząsteczki glukozy w warunkach tlenowych do dwutlenku węgla
i wody resyntetyzuje się 38 cząsteczek ATP. W warunkach tlenowych
glikoliza przebiega również przez cykl pentozowy.
Glukoza
Glukozo -6- fosforan
I
Fruktozo -6- fosforan
\ Fruktozo-1,6-di
fosforan
\ Aldehyd-3-fosfoglicery
nowy
\ 1,3difosfoglicerynian
3- fosfoglicerynian
Fosfoenolopirogronian
I
Pirogronian
I
Mleczan
Ryc. 220. Glikoliza beztlenowa.
Ryc. 222. Resynteza ATP w czasie glikolizy
tlenowej.
14 — Fizjologia człowieka
418
Oddychanie
Ryc. 223. Glikoliza, cykl pentozowy.
I Przemiana w organizmie
Wyzwolenie energii z węglowodanów, białek i tłuszczów wiąże się
z utlenianiem produktów ich rozpadu. Mierząc zużycie tlenu w jednostce
czasu, można pośrednio oznaczyć energię wytworzoną w organizmie.
Zużycie 1 L tlenu przez organizm wyzwala przeciętnie około 20,2 kJ. Jest
to energetyczny równoważnik tlenu oznaczony metodą kalorymetrii pośredniej, w odróżnieniu od kalorymetrii bezpośredniej polegającej na
oznaczeniu energii wytworzonej w organizmie na podstawie wartości
energetycznej spożytych pokarmów.
Współczynnik oddechowy
Współczynnik oddechowy — RQ (respiratory ąuotient) — wskazuje na
stosunek objętościowy pomiędzy uwolnionym z organizmu dwutlenkiem
węgla i pochłoniętym tlenem w jednostce czasu. Współczynnik oddechowy
dla węglowodanów wynosi 1,0, dla białek średnio około 0,82 i dla
tłuszczów około 0,7. Węglowodany zawierają w każdej cząsteczce jednakowy stosunek atomów wodoru do atomów tlenu. Cały tlen pochłaniany w czasie utleniania węglowodanów jest zużywany do wytwarzania dwutlenku węgla. Utlenianie tłuszczów wymaga więcej tlenu.
Jest on zużywany w organizmie do wytwarzania jednocześnie dwutlenku
węgla i wody.
Przemiana w organizmie
C02
250 mL /min
02
200
41 9
Ryc. 224. Schemat układów sercowo-naczyniowego i oddechowego. Tlen pochłonięty z powietrza atmosferycznego transportowany jest do komórek, dwutlenek
węgla tworzący się w komórkach transportowany jest od komórek do powietrza
atmosferycznego. U „standardowego" mężczyzny w spoczynku w celu zachowania
podstawowej przemiany materii pochłaniane jest 200 mL tlenu i wydalane 250 mL
dwutlenku węgla na minutę.
mL/min
Tkanki
I Podstawowa przemiana materii
Mierząc zużycie tlenu przez organizm w ściśle określonych warunkach,
można uzyskać dane porównawcze odnoszące się do przemiany materii
w organizmach różnych ludzi.
Warunki pomiaru zużycia tlenu w jednostce czasu są następujące:
• całkowity spoczynek fizyczny i psychiczny, pozycja leżąca;
• od 12 do 14 godzin po ostatnim posiłku;
• temperatura otoczenia +20°C.
420
Oddychanie
Zużycie tlenu w jednostce czasu w tych warunkach wiąże się z wyzwoleniem energii dla procesów fizjologicznych niezbędnych do utrzymania człowieka przy życiu i nosi nazwę podstawowej przemiany materii
— BMR (basal metabolic ratę).
Podstawowa przemiana materii zależy od:
• powierzchni ciała;
• wieku badanego człowieka;
• płci.
Powierzchnię ciała u ludzi oblicza się na podstawie wzrostu i masy ciała
według wzoru:
S = 0,007184xW°-*"xH°'725
gdzie:
S — powierzchnia ciała w metrach kwadratowych,
W — masa ciała w kilogramach,
H — wzrost w centymetrach.
Podstawowa przemiana materii w przeliczeniu na metr kwadratowy
powierzchni ciała zmniejsza się w miarę przybywania lat. U płci męskiej jest
ona większa, a u płci żeńskiej mniejsza. U mężczyzn w wieku 20 lat
podstawowa przemiana wynosi 48,15 J/m2/s, a u kobiet w tym samym
wieku wynosi 41,98 J/m2/s. Przeciętnie u ludzi dorosłych podstawowa
przemiana materii wynosi u mężczyzn około 7,28 MJ na dobę, u kobiet
zaś 5,62 MJ na dobę.
Czynniki wpływające na przemianę materii
Wiele czynników pobudza przemianę materii i zwiększa zużycie tlenu
przez organizm w jednostce czasu. Są to:
• praca fizyczna;
• praca umysłowa i stany emocjonalne;
• niska lub wysoka temperatura otoczenia;
• zwiększenie zawartości niektórych hormonów we krwi, zwłaszcza
hormonów gruczołu tarczowego i hormonów rdzenia nadnerczy;
• trawienie i przyswajanie pokarmów.
Praca fizyczna zwiększa zużycie energii w mięśniach szkieletowych.
U ludzi pracujących fizycznie przemiana materii jest 0,5—2 razy większa
od podstawowej przemiany materii.
Przemiana w organizmie
421
Zapotrzebowanie na energię u „standardowego" mężczyzny w wieku
od 20 do 39 lat, o masie ciała 70 kg, o odpowiednim wzroście w stosunku do
masy ciała i wykonującego umiarkowany wysiłek fizyczny w ciągu doby
wynosi 13,5 MJ. Umiarkowany wysiłek fizyczny w ciągu doby obejmuje:
8 godzin pracy fizycznej o umiarkowanej ciężkości, 6 godzin zajęć
siedzących, 2 godziny chodzenia i 8 godzin leżenia w łóżku. Dla tego
samego mężczyzny wykonującego mały wysiłek fizyczny zapotrzebowanie
na energię wynosi 12,3 MJ/d, duży wysiłek fizyczny — 15,8 MJ/d i bardzo
duży wysiłek — 18,2 MJ/d.
U „standardowej" kobiety w wieku od 20 do 39 lat, o masie ciała 55 kg
i o odpowiednim do masy ciała wzroście, umiarkowany wysiłek fizyczny
w ciągu doby wymaga energii w ilości 9,2 MJ, odpowiednio mały
wysiłek — 8,4 MJ/d, duży wysiłek — 10,9 MJ/d i bardzo duży wysiłek —
12,6 MJ/d. Ciąża wymaga dodatkowej energii w ilości około 335 MJ
w ciągu całego jej trwania. W pierwszym trymestrze ciąży dodatkowo —
0,6 MJ/d, w drugim i w trzecim trymestrze dodatkowo — 1,5 MJ/d.
W okresie laktacji zapotrzebowanie na energię jest większe niż w czasie
ciąży o dodatkowe 2,3 MJ/d.
Tabela 32
Dzienne zapotrzebowanie na energię
Płeć
JYICZczyzni
Kobiety
IM asa
ciulu
(kg)
Mała
Umiarkowana
Duża
Bardzo duża
aktywność
fizyczna
(kcal) (SI: M J)
aktywność
fizyczna
(kcal) (SI: M J)
aktywność
fizyczna
(kcal) (SI: MJ)
aktywność
fizyczna
(kcal) (SI: MJ)
50
55
60
65
70
75
80
2100
2310
2520
2700
2940
3150
3360
40
45
50
55
60
65
70
1440
1620
1800
2000
2160
2340
2520
8,8 2300
9,7 2530
10,5
11,3
12,3
13,2
14,1
6,0
6,8
7,5
8,4
9,0
9,8
10,5
2760
3000
3220
3450
3680
1600
1800
2000
2200
2400
2600
2800
9,6 2700
10,6
11,5
12,5
13,5
14,4
15,4
6,7
7,5
8,4
9,2
10,0
10,9
11,7
2970
3240
3500
3780
4050
4320
1880
2120
2350
2600
2820
3055
3290
11,3
12,4
13,6
14,6
15,8
16,9
18,1
3100
3410
3720
4000
4340
4650
4960
13,0
14,3
15,6
16,7
18,2
19,5
20,8
7,9
8,9
9,8
2200
2480
2750
3000
3300
3575
3850
9,2
10,9
11,8
12,8
13,8
10,4
11,5
12,6
13,8
15,0
16,1
Zapotrzebowanie energetyczne opracowane przez Światową Organizację do Spraw Żywienia i Żywności (FAO) i Światową Organizację Zdrowia (WHO).
422
Oddychanie
Poza ogólnym zapotrzebowaniem na energię konieczne jest białko
w pokarmach, na które zapotrzebowanie u „standardowego" mężczyzny
wynosi 0,57 g na 1 kg masy ciała na dobę, u „standardowej kobiety" zaś
0,52 g na 1 kg masy ciała na dobę.
Niska temperatura otoczenia, poniżej 20°C, zwiększa utratę ciepła
i przyspiesza metabolizm w organizmie w celu utrzymania stałej temperatury ciała. Temperatura otoczenia wyższa o kilkanaście lub kilkadziesiąt stopni od 20°C wymaga dodatkowej energii na odprowadzenie
ciepła z organizmu i zapobieżenie przegrzaniu.
Trawienie i przyswajanie składników pokarmowych zwiększają przemianę materii, co nosi nazwę swoistego dynamicznego działania pokarmów — SDA (specific dynamie action of foods). W warunkach pomiaru
zużycia tlenu, wykonanego tak jak przy oznaczaniu podstawowej przemiany materii (BMR), po spożyciu węglowodanów lub tłuszczów w ilości
równoważnej 419 kJ następuje zwiększenie przemiany o kilkanaście
kilodżulów. Po spożyciu natomiast białka w ilości równoważnej 419 kJ
przemiana zwiększa się o 126 kJ. Świadczy to o tym, że białka wymagają
znacznie większego zużycia energii do ich przyswojenia niż węglowodany
i tłuszcze.
Układ trawienny
I Odżywianie
I Równowaga energetyczna
Funkcje życiowe organizmu człowieka wymagają stałego dostarczania
energii i wody ze środowiska zewnętrznego. Spożywanie pokarmów,
a więc ilość energii wprowadzana do organizmu (energy intake) wyrażona
w dżulach (J) zależy przede wszystkim od metabolizmu i zużycia energii
(energy utilization) wewnątrz organizmu. Podobnie ilość wypijanej wody
ściśle wiąże się z ilością wody wydalanej z organizmu przez nerki, skórę
i płuca.
W ciągu doby, która stanowi jeden pełny cykl w życiu człowieka,
powinna być zachowana równowaga energetyczna (energy balance) pomiędzy energią wprowadzoną w postaci pokarmów a energią zużytą. U dorosłego człowieka bilans wprowadzonej i zużytej w ciągu doby energii
powinien równać się zeru. Ilości spożywanych pokarmów i wody są
uwarunkowane zapotrzebowaniem.
Odżywianie obejmuje:
•
•
•
•
przyjmowanie pokarmów;
trawienie pokarmów;
wchłanianie składników pokarmowych i wody;
przyswajanie składników pokarmowych.
424
Układ trawienny
Przyjmowanie pokarmów
Kontrolę nad ilością spożywanych pokarmów, czyli nad ilością wpn
wadzonej do organizmu energii, pełnią ośrodki pokarmowe (feedin
centers) w podwzgórzu, z których jeden — „ośrodek głodu" (hunge
center) — wyzwala mechanizm poszukiwania, zdobywania i przyjmowe
nia pokarmów, drugi zaś — „ośrodek sytości" (satiety center) — hamuj
apetyt.Układ limbiczny hamuje „ośrodek głodu", kora mózgu i inn
ośrodki podkorowe zaś wywierają wpływ pobudzający lub hamujący n
ten ośrodek. Występujący w podwzgórzu neuropeptyd Y (NPY) pobu
dza „ośrodek głodu" i może być uważany za „czynnik głodu".
Zasadniczym czynnikiem wpływającym na pobudliwość ośrodkóv
pokarmowych jest leptyna (leptin), hormon białkowy wydzielany prze;
komórki tkanki tłuszczowej (adipocyty) działająca jak „czynnik sytości"
Spożyte i wchłonięte w przewodzie pokarmowym składniki pokarmowi
pobudzają adipocyty do wydzielania leptyny bezpośrednio i pośrednie
poprzez insulinę z wysp trzustkowych. Leptyna na zasadzie ujemnego
Ryc. 225. Schemat zależności pomiędzy podwzgórzowymi ośrodkami głodu i sytości,
przyjmowaniem pokarmów i działaniem leptyny wydzielanej przez komórki tłuszczowe.
Leptyna hamuje przyjmowanie pokarmów bezpośrednio, pobudzając ośrodek sytości,
i pośrednio, hamując ośrodek głodu. Leptyna hamuje tworzenie się w podwzgórzu
neuropeptydu Y (NPY) pobudzającego ośrodek głodu.
Trawienie pokarmów
425
sprzężenia zwrotnego poprzez ośrodki pokarmowe w podwzgórzu hamuje
przyjmowanie pokarmów i tym samym hamuje magazynowanie energii
w tkance tłuszczowej.
U ludzi z nadwagą występuje duże stężenie leptyny we krwi, proporcjonalne do masy tkanki tłuszczowej, i jednoczesny brak hamowania
spożywania pokarmów. Świadczy to o niewrażliwości ośrodka sytości na
zwiększające się stężenie leptyny we krwi i na przewagę oddziaływania
podwzgórzowego neuropeptydu Y na „ośrodek głodu".
Impulsacja odbierana z interoreceptorów w ścianach przewodu pokarmowego zmienia pobudliwość „ośrodka sytości". Rozciągnięcie ścian
przewodu pokarmowego pobudza ten ośrodek. Apetyt jest hamowany
również przez zwiększone pragnienie wywołane wzrostem ciśnienia osmotycznego osocza krwi na skutek utraty wody z organizmu lub jej
wydzielaniem przez gruczoły przewodu pokarmowego wraz z innymi
składnikami soków trawiennych.
I Trawienie pokarmów
Pokarmy spożywane zostają w układzie trawiennym (systema digestorium) poddane działaniu czynników mechanicznych i chemicznych, co
doprowadza je do postaci, w której mogą być wchłonięte.
Pokarmy zawierają składniki odżywcze, jakimi są węglowodany, białka
i tłuszcze oraz witaminy, sole mineralne i woda. Węglowodany, białka
i tłuszcze spożywane są zazwyczaj w postaci wymagającej obróbki
mechanicznej pokarmów tak, aby enzymy soków trawiennych mogły
rozłożyć je na podstawowe wchłanialne składniki, którymi są cukry
proste, aminokwasy i kwasy tłuszczowe.
Witaminy i sole mineralne są uwalniane z pokarmów dzięki obróbce
mechanicznej, a tylko w mniejszym stopniu na skutek działania czynników
chemicznych.
I Jama ustna i przełyk
Pokarmy o stałej konsystencji wprowadzone do jamy ustnej (cavum oris)
są rozdrabniane i mieszane ze śliną w procesie żucia. Żucie pokarmów trwa
tak długo, aż zostaną one odpowiednio rozdrobnione i nasycone śliną,
aby uformowane w postaci kęsa mogły zostać połknięte.
426
Uk/ad trawienny
Wydzielanie śliny
Zetknięcie się pokarmu, zwłaszcza suchego, z powierzchnią błony
śluzowej jamy ustnej powoduje wydzielanie śliny na drodze odruchu
bezwarunkowego. Ślina i inne soki trawienne mogą wydzielać się na sam
widok lub zapach pokarmu. W tym przypadku wydzielanie soków
trawiennych następuje dzięki uprzednio wytworzonemu odruchowi nabytemu, czyli odruchowi warunkowemu.
Ślina w ilości około 1,5 L na dobę i pH około 7,0 (SI: 100 nmol/L H+)
jest wydzielana do jamy ustnej przez trzy parzyste gruczoły: śliniankę
podjęzykową (glandula sublingualis) , śliniankę podżuchwową {glandula
submandibularis) i śliniankę przyuszną {glandula parotis). Włókna nerwowe przywspółczulne unerwiają wydzielniczo slinianki. Włókna biegnące wraz z nerwem twarzowym (n. VII) unerwiają slinianki podjęzykową
i podżuchwową, a wraz z nerwem językowo-gardłowym (n. IX) biegną
włókna do slinianki przyusznej.
Tabela 33
Skład śliny mieszanej w jednym litrze
Stężenie
Składniki
Woda
Sucha pozostałość:
0,994
6—7
L
g/L
(994 mL)
(6—7 g)
(3000 mg)
(2000 mg)
(100 mg)
(20 mg)
Białko
Mucyna
Mocznik
Kwas moczowy
3
2
1,66
109
g/L
g/L
mmol/L
umol/L
Cholesterol
182
umol/L
Substancja nieorganiczna:
(70 mg)
2,8
g/L
(2800 mg)
Sód
Potas
26,09
10,23
mmol/L
mmol/L
(600 mg)
(400 mg)
Wapń
1,49
mmol/L
(60 mg)
Magnez
0,82
mmol/L
(20 mg)
Chlor
Fosfor
16,92
6,46
mmol/L
mmol/L
(600 mg)
(200 mg)
Wartości podane zostały w jednostkach zgodnych z układem SI i w tradycyjnych
(w nawiasach).
Trawienie pokarmów
427
Najwięcej śliny surowiczo-śluzowej wydzielają ślinianki podżuchwowe.
Ślinianki przyuszne wydzielają w mniejszych ilościach ślinę surowiczą,
najmniej śliny śluzowej wytwarzają zaś ślinianki podjęzykowe.
Ślina surowicza zawiera enzym trawiący wielucukry — alpha-amylazę
ślinową. Natomiast w ślinie śluzowej występują mucyny — glikoproteidy
ułatwiające połykanie uformowanego kęsa.
Połykanie
Połykanie dzieli się na trzy fazy, z których pierwsza ustno-gardłowa jest
fazą dowolną i polega na przesunięciu kęsa z jamy ustnej (cctvum oris) do
gardła (pharynx), w czym uczestniczą mięśnie języka i policzków.
Druga i trzecia faza są odruchowe. W fazie drugiej gardłowo-przełykowej mięsień zwieracz górny gardła (musculus constrictor pharyngis
Ryc. 226. Połykanie kęsa pokarmu oznaczonego kolorem czerwonym: a — początek fazy
ustno-gardłowej; b — faza ustno-gardlowa; c — faza gardłowo-przelykowa; d — faza
przełykowo-żołądkowa.
428
Układ trawienny
superior) początkowo rozkurcza się przepuszczając kęs. Następnie kurczy
się i rozpoczyna falę perystaltyczną przesuwającą kęs wzdłuż gardła
i przełyku.
W fazie trzeciej połykania, przełykowo-żołądkowej, kęs przesuwany
przez falę perystaltyczną zbliża się do wpustu {cardid), wywołuje jego
rozkurcz i dostaje się do jamy żołądka (cavum ventriculi).
I Żołądek
Żołądek (gaster) spełnia w organizmie człowieka trzy zasadnicze
funkcje:
• gromadzi i przechowuje spożywane pokarmy;
• trawi pokarmy;
• wyjaławia pokarmy.
Gromadzenie pokarmów w żołądku
Pokarmy płynne po dostaniu się do pustego żołądka spływają wzdłuż
krzywizny mniejszej (curvatura gastrica minor) do części odźwiernikowej
(parspylorica gastrica). Pokarmy o stałej konsystencji początkowo wypełniają trzon żołądka (corpus gastricum), a następnie w miarę ich przybywania zajmują dno żołądka (fundus gastricus). Kolejne spożywane porcje
pokarmów zajmują część środkową jamy żołądka, rozpychając w kierunku ścian żołądka znajdujący się w nim wcześniej przyjęty pokarm. Pierwsze
porcje pokarmu stykają się z błoną śluzową (tunica mucosa) żołądka,
ostatnie porcje wypełniają zaś środek jamy żołądka.
Trawienie pokarmów w żołądku
Pokarmy wypełniające część środkową żołądka, nie stykające się
z błoną śluzową, są trawione początkowo przez alpha-amylazę ślinową.
Zmieszanie się treści pokarmowej z sokiem żołądkowym rozpoczyna
właściwe trawienie w żołądku.
Sok żołądkowy jest wydzielany przez gruczoły błony śluzowej żołądka
średnio w ilości 3 L w ciągu całej doby, o pH około 1,0 (SI: 100 mmol/L
H+). Sok żołądkowy zawiera kwas solny, enzymy trawienne, śluz, sole
mineralne i wodę.
Trawienie pokarmów
429
Ryc. 227. Schemat budowy gruczołu błony
śluzowej trzonu żołądka. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie
PZWL, Warszawa 1997.)
Kwas solny (HC1) powstaje w kanalikach wewnątrz komórek okładzinowych gruczołów błony śluzowej pod wpływem histaminy działającej na
receptory H2. Jony wodorowe powstające w czasie dysocjacji kwasu
węglowego są aktywnie wydzielane do światła kanalików komórkowych.
W tym aktywnym transporcie jonów wodorowych do światła kanalików
biorą udział cyklaza adenylanowa i cykliczny AMP oraz prawdopodobnie również adenozynotrifosfataza. Jednocześnie z jonami wodorowymi
wydzielane są jony chlorowe i w świetle kanalików tworzy się kwas solny.
W komórkach głównych gruczołów błony śluzowej żołądka znajdują się
w cytoplazmie ziarnistości zawierające pepsynogen — nieczynny enzym
430
Układ trawienny
Ryc. 228. Komórka główna (a) i komórka okładzinowa (b) gruczołów błony śluzowej
żołądka. 1 — ziarno wydzielnicze, 2 — aparat Golgiego, 3 —jądro, 4 — ziarnista siateczka
śródplazmatyczna, 5 — kanalik wewnątrzkomórkowy, 6 — mitochondria, 7 — błona
podstawna. (Reprodukowano za zezwoleniem Michała Karaska.)
proteolityczny. Wydzielony do światła gruczołów błony śluzowej zamienia się pod wpływem kwasu solnego na aktywny enzym — pepsynę.
Komórki dodatkowe gruczołów błony śluzowej wydzielają śluz, który,
pokrywając ściany żołądka, zabezpiecza je przed działaniem kwasu
solnego i enzymów.
Pepsyna zawarta w soku żołądkowym rozkłada duże cząsteczki białek
na mniejsze cząsteczki polipeptydów. Przerywa wiązania peptydowe
w cząsteczkach białek i powstające z nich cząsteczki mają zarówno krótkie, jak i długie łańcuchy. Największa aktywność pepsyny występuje
w granicach pH od 1,6 do 3,2 (SI: 21,88 mmol/L H+ do 631 umol/L H+).
Motoryka żołądka
Pusty żołądek w kilka lub kilkanaście godzin po opróżnieniu wykazuje
okresowo występujące silne skurcze błony mięśniowej, tzw. skurcze
głodowe. Po dostaniu się pierwszych porcji pokarmu do żołądka następuje
chwilowy rozkurcz mięśni ścian żołądka. Następnie skurcze błony mię-
Trawienie pokarmów
431
Tabela 34
Skład soku żołądkowego w jednym litrze
Składniki
Stężenie
Woda
Sucha pozostałość
Związki organiczne:
L
g/L
g/L
Białko
0,990—0,995
5—10
4—6
4
g/L
(990—995 mL)
(5-10 g)
(4—6 g)
(4 g)
Azot całkowity
35,7—57,1
mmol/L
(500—800 mg)
Azot aminokwasów
2,14—7,14
mmol/L
(30—100 mg)
89,2
umol/L
Kwas moczowy
(15 mg)
Związki nieorganiczne:
1,3—1,4
Kwas solny wolny
109,5—136,9
mmol/L
(4,0—5,0 g)
Sód
Chlor
46,5
140,8—169,0
mmol/L
mmol/L
(1,07 g)
(5,0—6,0 g)
g/L
(1,3—1,4 g)
Wartości podane zostały w jednostkach zgodnych z układem SI i w tradycyjnych
(w nawiasach).
śniowej powracają i przyczyniają się zarówno do mieszania się treści
pokarmowej z sokiem żołądkowym, jak i do opróżnienia się żołądka.
Wyróżnia się dwa rodzaje aktywności ruchowej błony mięśniowej
żołądka:
• nasilenie i osłabienie napięcia całej błony mięśniowej żołądka wy
stępujące naprzemiennie, z czym wiążą się wahania ciśnienia w jamie
żołądka;
• skurcze perystaltyczne rozpoczynające się w okolicy części wpustowej
(pars cardiaca) i przesuwające się wzdłuż ścian żołądka aż do odźwiernika (pyloruś).
W części wpustowej żołądka średnio co 20 sekund występuje skurcz
warstwy okrężnej, który posuwa się wzdłuż ścian w postaci fali perystaltycznej i dochodząc do odźwiernika, powoduje jego otwarcie i przejście
z żołądka do dwunastnicy płynnej treści żołądkowej.
Fala perystaltyczna, przesuwając się wzdłuż ścian żołądka, po dotarciu
do części odźwiernikowej (pars pylorica) staje się znacznie silniejsza dzięki
skurczowi grubszej warstwy mięśni w tej części żołądka. Silny ucisk
wywierany przez skurczoną ścianę tej części żołądka przenosi się na treść
432
Układ trawienny
Ryc. 229. Motoryka pustego żołądka zarejestrowana przy użyciu balonika wprowadzonego
do jamy żołądka. (Reprodukowano za zezwoleniem firmy CIBA-GEIGY Ltd, Bazylea,
Szwajcaria z F. H. Netter The CIBA Collection of Medical Illustration 1959, vol. 3. Digestive
System. Wszystkie prawa zastrzeżone.)
wypełniającą jamę odźwiernikową (antrum pyloricum). Następuje wyciśnięcie treści płynnej do dwunastnicy przez otwierający się odźwiernik
i cofnięcie treści stałej do jamy żołądka. Powtarzające się w ciągu kilku
godzin trawienia w żołądku cofanie się treści stałej do jamy żołądka
powoduje jej mieszanie, rozdrobnienie i stopniową zamianę na treść
płynną. Wyciskanie płynnej treści do dwunastnicy i cofanie do żołądka
treści stałej nosi nazwę „pompy odźwiernikowej".
Regularne występowanie skurczów perystaltycznych wyzwalane jest
przez rozrusznik, który tworzą komórki mięśniowe gładkie warstwy
podłużnej błony mięśniowej żołądka znajdujące się w sąsiedztwie krzywizny większej. W komórkach rozrusznika regularnie i spontanicznie
występuje stan czynny, który rozprzestrzenia się na całą błonę mięśniową,
o czym świadczy podstawowy rytm bioelektryczny — BER (basie eleetrie
rhythm) — występujący w tej okolicy żołądka. Transmittery uwalniane na
drodze odruchowej z komórek nerwowych splotów autonomicznych
żołądka i hormony żołądkowo-jelitowe przyspieszają lub zwalniają częstotliwość występowania stanu czynnego w komórkach rozrusznika.
Pokarmy płynne szybko przechodzą przez żołądek. Pokarmy składające się z węglowodanów pozostają w żołądku kilka godzin. Dłużej
pozostają pokarmy o przewadze białek, a najdłużej bogate w tłuszcze.
Trawienie pokarmów
433
Ryc. 230. Wypełnianie się jamy żołądka kolejnymi porcjami pokarmu I, II i III (a)
i przechodzenie pokarmów płynnych (b). (Reprodukowano za zezwoleniem firmy CIBA-GEIGY Ltd, Bazylea, Szwajcaria z F. H. Netter The CIBA Collection of Medical Illustration 1959, vol. 3. Digestive System. Wszystkie prawa zastrzeżone.)
Kontrola motoryki żołądka
Na skurcze toniczne błony mięśniowej i perystaltykę żołądka mają
wpływ czynniki nerwowe i humoralne. Nerw błędny (n. X) poprzez
przywspołczulne neurony śródścienne nasila skurcze i przyspiesza perystaltykę żołądka. Przeciwnie działają nerwy współczulne.
Czynniki hamujące na drodze odruchowej i humoralnej perystaltykę
żołądka i zwalniające jego opróżnianie to:
• rozciągnięcie ścian dwunastnicy przez pokarm przechodzący z żołądka,
czemu towarzyszy zmniejszenie wartości pH w dwunastnicy;
434
Układ trawienny
• zawartość kwasów tłuszczowych, częściowo strawionych białek i węg
lowodanów w treści pokarmowej przechodzącej do dwunastnicy;
• sekretyna i cholecystokinina powstające w dwunastnicy;
• wysokie ciśnienie osmotyczne treści żołądkowej dostającej się do
dwunastnicy.
Ryc. 231. Działanie pompy odźwiernikowej w czasie dochodzenia fali perystaltycznej do
części odźwiernikowej żołądka (a i b), otwieranie się odźwiernika (c i e) i cofanie się
treści pokarmowej do jamy żołądka (d i f). (Reprodukowano za zezwoleniem firmy CIBAGEIGY Ltd, Bazylea, Szwajcaria z F. H. Netter The CIBA Collection of Medical
Illustration 1959, vol. 3. Digestive System. Wszystkie prawa zastrzeżone.)
Kontrola wydzielania soku żołądkowego
Wydzielanie soku żołądkowego dzieli się na trzy fazy, które w warunkach fizjologicznych występują często prawie jednocześnie i zachodzą na
siebie. Są to:
• faza głowowa;
• faza żołądkowa;
• faza jelitowa.
W fazie głowowej (cephalic phase), nazywanej dawniej fazą nerwową,
występuje wydzielanie soku żołądkowego pod wpływem impulsów bieg-
Trawienie pokarmów
FAZA GŁOWOWA
435
FAZA ŻOŁĄDKOWA
Ryc. 232. Kontrola wydzielania soku żołądkowego w fazie głowowej i w fazie żołądkowej.
436
Układ trawienny
nących przez nerwy błędne. W fazie tej uczestniczą zarówno odruchy
warunkowe, czyli nabyte, jak i odruchy bezwarunkowe, wrodzone, kiedy
pokarm znajduje się już w jamie ustnej i drażni receptory w błonie śluzowej. Acetylocholina uwolniona z zakończeń śródściennych neuronów
przywspółczulnych działa na:
• komórki główne błony śluzowej żołądka — bezpośrednio i pośrednio —
poprzez neurony zawierające peptyd uwalniający gastrynę (GRP)
i gastrynę wydzielaną przez komórki dokrewne G;
• komórki okładzinowe błony śluzowej żołądka za pośrednictwem his
taminy i receptora histaminowego H2 .
W fazie żołądkowej (gastric phase) na skutek podrażnienia błony
śluzowej przez pokarm wypełniający żołądek zachodzi odruchowe, jak
i bezpośrednie oddziaływanie na komórki dokrewne G znajdujące się
Ryc. 233 a — wydzielanie gastryny przez komórki G pod wpływem impulsacji eferentnej
n. X i śródściennego odruchu. Gastryna na drodze endokrynnej (linia przerywana czer wona) pobudza komórki główne i okładzinowe; b — hamowanie wydzielania
gastryny i soku żołądkowego na drodze parakrynnej (linia przerywana czarna) pod
wpływem somatostatyny (SRIF) uwalnianej przez komórki D. GRP — peptyd uwalniający
gastrynę.
r
Trawienie pokarmów
437
w błonie śluzowej żołądka. Wydzielają one do krwi gastrynę. Krążąc we
krwi gastryna na drodze humoralnej pobudza do wydzielania gruczoły
błony śluzowej żołądka.
W fazie jelitowej (intestinal phase) pod wpływem treści pokarmowej
przechodzącej z żołądka do dwunastnicy, na drodze nerwowej i humoralnej, zachodzi zarówno pobudzanie, jak i hamowanie czynności żołądka.
Wytwarzana w dwunastnicy gastryna i cholecystokinina (CCK) pobudzają
wydzielanie soku żołądkowego, natomiast sekretyna działa hamująco
(patrz Hormony żołądkowo-jelitowe, str. 450). Za pośrednictwem odruchu
jelitowo-żołądkowego następuje hamowanie opróżniania żołądka i wydzielania soku żołądkowego.
Hormony wytwarzane przez gruczoły dokrewne mają również wpływ na
wydzielanie soku żołądkowego. Hormony kory nadnerczy (glikokortykoidy) zwiększają wydzielanie soku żołądkowego, natomiast hormony
rdzenia nadnerczy (adrenalina i noradrenalina) hamują jego wydzielanie.
I Jelito cienkie
Jelito cienkie (intestinum tenue) stanowi niezbędny odcinek przewodu
pokarmowego. Treść pokarmowa jest tam trawiona do składników
prostych, które są w tym odcinku wchłaniane do krwi.
Motoryka jelita cienkiego
Błona mięśniowa jelita cienkiego wykazuje:
• okresowe zmiany napięcia;
• skurcze odcinkowe (segmentation contractions) zaznaczające się okre
sowym pojawianiem się i zanikaniem okrężnych przewężeń jelita;
• skurcze perystaltyczne (peristaltic contractions).
Skurcze perystaltyczne rozpoczynają się w dwunastnicy i przesuwają
w postaci fali perystaltycznej z różną prędkością od 2 do 25 cm/s przez
kolejne odcinki jelita cienkiego i jelita grubego aż do odbytnicy (rectum).
Płynna treść przechodząca z żołądka do dwunastnicy {duodenum) gromadzi się w części górnej dwunastnicy {pars superior duodeni). Po zgromadzeniu się w części górnej dwunastnicy trzech kolejnych porcji treści żołądkowej występuje przeciętnie jeden raz na minutę okrężny skurcz błony
mięśniowej rozpoczynający falę perystaltyczną.
438
Układ trawienny
Fala perystaltyczna w postaci przesuwającego się okrężnego skurczu
błony mięśniowej jest zasadniczym czynnikiem zapewniającym przesuwanie się treści jelitowej w kierunku od żołądka do odbytnicy. Powstaje ona
pod wpływem mechanicznego podrażnienia receptorów znajdujących się
w błonie śluzowej i pobudzenia komórek nerwowych splotu warstwy
mięśniowej za pośrednictwem komórek dwubiegunowych splotu podśluzowego. Komórki splotu warstwy mięśniowej, unerwiając mięśnie
gładkie jelita, powodują przesuwanie się skurczu perystaltycznego, za
którym postępuje rozkurcz.
Ryc. 234. Ruchy jelita cienkiego: a — okresowe zmiany napięcia błony mięśniowej;
b — ruchy wahadłowe; c — skurcze odcinkowe; d — skurcze perystaityczne.
Treść jelitowa przesuwana przez fale perystaityczne gromadzi się
w części jelita krętego (ileum) w pobliżu jego połączenia z jelitem ślepym
(cecum). Zastawka krętniczo-kątnicza (valva ileocecalis) jest zamknięta
i nie przepuszcza treści jelitowej do jelita ślepego. Po podniesieniu się
ciśnienia w jelicie krętym w pobliżu zastawki krętniczo-kątniczej treść
jelitowa zaczyna przechodzić do jelita ślepego.
Przechodzenie treści żołądkowej do dwunastnicy powoduje błyskawiczne ruchy perystaityczne jelita cienkiego, jednoczesne otwieranie się zastawki krętniczo-kątniczej i opróżnianie jelita krętego z treści jelitowej.
Otwieranie się zastawki krętniczo-kątniczej z chwilą rozpoczęcia opróżniania się żołądka nosi nazwę odruchu żołądkowo-krętniczego (gastroileal
reflex), który powstaje za pośrednictwem nerwów błędnych.
Trawienie pokarmów
439
Ryc. 235. Otwieranie się zastawki krętniczo-kątniczej i opróżnianie jelita krętego do kątnicy.
Błona śluzowa jelita cienkiego pokryta jest kosmkami (villi intestinales)
długości 0,5—1 mm. Występują one w liczbie 20—40 na 1 mm2 błony
śluzowej. Każdy kosmek ma kilka mikrokosmków. Łączna powierzchnia
jelita cienkiego po uwzględnieniu powierzchni kosmków i mikrokosmków
wynosi około 300 m2.
Kontrola motoryki jelita cienkiego
W czasie trawienia pokarmów występują zarówno skurcze błony
mięśniowej jelita, jak i ruchy kosmków. Ruchy kosmków polegają na ich
skracaniu się i wydłużaniu. Zmiany napięcia mięśni , ruchy wahadłowe,
skurcze odcinkowe, a zwłaszcza skurcze perystaltyczne, występują na
skutek pobudzenia komórek nerwowych w splotach jelitowych.
W splocie podśluzowym jelita (plexus submucosus) i w splocie warstwy
mięśniowej jelita {plexus myentericus) występują neurony o różnorodnej
funkcji. Są to neurony:
• przewodzące pobudzenie od receptorów w błonie śluzowej;
• pośredniczące;
440
Układ trawienny
unerwiające komórki gruczołowe błony śluzowej;
unerwiające błonę mięśniową jelita; unerwiające
naczynia krwionośne.
W neuronach tych transmitterami są: acetylocholina (ACh) oraz prawdopodobnie serotonina (5-HT) i kwas gamma-aminomasłowy (GABA).
W neuronach cholinergicznych wraz z acetylocholina koegzystuje jeden
lub kilka modulatorów. Są neurony, w których występuje Substancja P
(SP), w innych zaś cholecystokinina (CCK), dynorfina (DYN), enkefalina
(ENK), peptyd uwalniający gastrynę (GRP) i wazoaktywny peptyd
jelitowy (VIP), w jeszcze innych — cholecystokinina (CCK), peptyd
pochodny kalcytoninowego genu (CGRP), neuropeptyd Y (NPY) i somatostatyna (SRIF). Neurony o nie zidentyfikowanym transmitterze zawierają
dynorfinę (DYN), enkefalinę (ENK), neuropeptyd Y (NPY) i wazoaktywny peptyd jelitowy (VIP).
Impulsacja przewodzona przez nerwy błędne i nerwy współczulne
wywiera wpływ za pośrednictwem neuronów splotów jelitowych. Impulsacja nerwu błędnego nasila skurcze błony mięśniowej jelita i ruchy
kosmków, przewodzona przez nerwy współczulne zaś ma działanie przeciwne — hamujące.
I Trawienie w jelicie cienkim
W trawieniu treści wypełniającej jelito cienkie biorą udział:
• sok jelitowy;
• sok trzustkowy;
• żółć.
Wydzielanie soku jelitowego
W błonie śluzowej początkowego odcinka dwunastnicy występują
gruczoły dwunastnicze podśluzówkowe (Brunnera) wydzielające gęsty śluz.
Zabezpiecza on błonę śluzową dwunastnicy przed działaniem silnie
zakwaszonej treści żołądkowej. Znajdujące się w błonie śluzowej jelita
cienkiego gruczoły jelitowe (Lieberkuhna) wytwarzają sok jelitowy, zawierający enzymy trawienne rozkładające związki znajdujące się w treści
jelitowej do składników prostych, wchłanialnych.
Trawienie pokarmów
441
W soku jelitowym występują:
• aminopeptydazy rozkładające peptydy do aminokwasów;
• enzymy rozkładające kwasy nukleinowe do pentozy, zasad purynowych
i pirymidynowych oraz kwasu fosforowego;
• enzymy rozkładające wielocukry i dwucukry do jednocukrów;
• lipaza hydrolizująca tłuszcze obojętne do kwasów tłuszczowych i glicerolu.
Sok jelitowy ma odczyn słabo zasadowy lub zasadowy. Ilość jego
wydzielana w ciągu doby wynosi od 3 do 6 L, zmienia się również jego
skład. Sok jelitowy poza hydrolizą składników pokarmowych i alkalizacją
treści jelitowej doprowadza jej ciśnienie osmotyczne do ciśnienia izotonicznego. Sok jelitowy wydziela się pod wpływem bezpośredniego mechanicznego podrażnienia błony śluzowej przez przesuwającą się treść jelitową
i pośrednio pod wpływem pobudzającego działania hormonów żołądkowo-jelitowych.
I Wydzielanie soku trzustkowego
Sok trzustkowy jest wydzieliną zewnętrzną trzustki (pancreas), która
przez przewód trzustkowy {ductus pancreaticus) i bańkę wątrobowo-trzustkową {ampulla hepatopancreatica) dostaje się do dwunastnicy.
W ciągu doby wydziela się około 2 L soku trzustkowego o odczynie
zasadowym i pH w granicach 7,1—8,4 (SI: 79,43—3,98 nmol H+/L).
Ponieważ sok trzustkowy jest szczególnie bogaty w węglowodany, pod
jego wpływem kwaśna treść żołądka dostająca się do dwunastnicy zostaje
szybko zobojętniona.
Sok trzustkowy zawiera:
• trypsynogen i chymotrypsynogen — nieaktywne enzymy proteolityczne;
• rybonukleazę i deoksyrybonukleazę — enzymy trawiące kwasy rybonukleinowy i deoksyrybonukleinowy;
• alpha-amylazę — enzym rozkładający wielocukry do dwucukrów;
• lipazę — enzym hydrolizujący tłuszcze roślinne i zwierzęce (o punkcie
topnienia niższym od 46°C) do kwasów tłuszczowych i glicerolu.
Trypsynogen znajdujący się w soku trzustkowym po dostaniu się do
dwunastnicy zostaje zamieniany pod wpływem enzymu enteropeptydazy
(enterokinazy) na enzym aktywny. Enterokinaza wydzielana przez błonę
śluzową dwunastnicy aktywuje nieaktywny trypsynogen na aktywną
442
Układ trawienny
trypsynę, która z kolei aktywuje następne porcje trypsynogenu dostające
się do dwunastnicy, jak również zmienia nieaktywny chymotrypsynogen
na aktywną chymotrypsynę.
Tabela 35
Skład sodu trzustkowego w jednym litrze
Składniki
Stężenie
Sód
Potas
147,8—188,2
4,1—12,3
mmol/L
mmol/L
Wapń
0,12—4,6
mmol/L
Magnez
ślad—0,94
mmol/L
(ślad —23
mg)
Chlor
48,7—157,7
mmol/L
(1730—5600
mg)
Siarka
Fosfor
1,93—2,53
0,09—1,22
mmol/L
mmol/L
(62—81
(3—38
mg)
mg)
47,45—85,92
46,4—200
mmol/L
mmol/L
(2900—5250
(650—2800
mg)
mg)
(280—1300
mg)
Wodorowęglany
Azot ogólny
Azot resztkowy
20—92,8
mmol/L
Mocznik
Kwas moczowy
0,166—8,3
ślad—83,3
mmol/L
umol/L
(3400--4330
(161—480
(5—184
(10—500
(ślad —14
mg)
mg)
mg)
mg)
mg)
Wartości podane zostały w jednostkach zgodnych z układem SI i w tradycyjnych
(w nawiasach).
Przewód
wyprowadzający
Pęcherzyki
wydzielnicze
Iziarna zymogenu)
Komórki
iródpecherzykowe
Ryc. 236. Schemat budowy pęcherzyka
wydzielniczego części zewnątrzwydzielniczej trzustki. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd.
Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Trawienie pokarmów
443
Ryc. 237. Komórka zewnątrzwydzielnicza trzustki. 1 — ziarna zymogenu, 2 — aparat
Golgiego, 3 — mitochondrium, 4 — jądro, 5 — ziarnista siateczka śródplazmatyczna,
6 — błona podstawna. (Reprodukowano za zezwoleniem Michała Karaska.)
Ryc. 238. Aktywacja w dwunastnicy trypsynogenu zawartego w soku trzustkowym pod
wpływem enterokinazy dwunastniczej.
444
Układ trawienny
Kontrola wydzielania soku trzustkowego
Trzustka wydziela sok trzustkowy zarówno pod wpływem impulsów
nerwowych, jak i czynników humoralnych. Wyróżnia się trzy fazy
wydzielania:
• głowową;
• żołądkową;
• jelitową.
Sok trzustkowy zaczyna wydzielać się na drodze odruchowej już wtedy,
kiedy pokarm znajduje się w jamie ustnej. Nerwem wydzielniczym dla
trzustki jest nerw błędny.
Opróżnianie się żołądka i przechodzenie treści żołądkowej do dwunastnicy wywołuje wydzielanie do krwi przez błonę śluzową dwunastnicy
Ryc. 239. Działanie hormonów żołądkowo-jelitowych wytwarzanych przez komórki błony
śluzowej dwunastnicy. Pod wpływem cholecystokininy (CCK) wydziela się sok trzustkowy
o dużej zawartości enzymów i kurczy się błona mięśniowa pęcherzyka żółciowego, sekretyna
(S) zaś pobudza wydzielanie żółci i soku trzustkowego bogatego w wodorowęglany.
Trawienie pokarmów
445
cholecystokininy (CCK), która krążąc we krwi pobudza komórki trzustki
do wydzielania soku trzustkowego bogatego w enzymy.
Zwiększenie się odczynu kwaśnego w dwunastnicy poniżej pH 5 (SI:
10 umol H+/L) pod wpływem większej, silnie zakwaszonej porcji treści
żołądkowej, powoduje wydzielanie do krwi przez błonę śluzową drugiego
aktywnego hormonu żołądkowo-jelitowego — sekretyny. Polipeptyd ten
po dostaniu się za pośrednictwem krwi do trzustki pobudza ją do
wydzielania dużych ilości soku trzustkowego ubogiego w enzymy, ale
o znacznej zawartości wodorowęglanów. Receptory dla cholecystokininy
znajdują się w błonie komórkowej komórek zewnątrzwydzielniczych
pęcherzyków trzustki. Receptory dla sekretyny natomiast, występują
zarówno w błonie komórkowej komórek zewnątrzwydzielniczych pęcherzyków trzustki, jak i w błonie komórkowej komórek przewodów wyprowadzających trzustki.
Sekretyna zwiększa również wydzielanie żółci, hamuje skurcze błony
mięśniowej żołądka i hamuje wydzielanie kwasu solnego przez gruczoły
błony śluzowej żołądka. Sok trzustkowy powstający pod wpływem sekretyny ma przede wszystkim znaczenie zabezpieczające dwunastnicę przed
uszkadzającym działaniem silnie kwaśnej treści żołądkowej.
I Wydzielanie żółci
Żółć wytwarzana w wątrobie przez komórki wątrobowe (hepatocyty)
zawiera liczne katabolity, które przez drogi żółciowe i przewód pokarmowy wydalane są z organizmu. Jednocześnie te katabolity mają duże
znaczenie dla procesu trawienia i wchłaniania w jelicie cienkim.
W czasie trawienia pokarmów w jelicie cienkim żółć wytwarzana przez
komórki wątrobowe odpływa do dwunastnicy przez przewodziki żółciowe
{ductuli billiferi), przewód wątrobowy prawy i lewy {ductus hepaticus dexter et sinister), przewód wątrobowy wspólny {ductus hepaticus communis),
przewód żółciowy wspólny {ductus choledochus) i bańkę wątrobowo-trzustkową {ampulla hepatopancreatica). W okresach pomiędzy trawieniem pokarmów mięsień zwieracz bańki wątrobowo-trzustkowej (Oddiego — musculus sphincter ampullae hepatopancreaticae) jest skurczony
i żółć gromadzi się w pęcherzyku żółciowym {vesica fellea).
W czasie opróżniania się żołądka do dwunastnicy występuje odruchowy — za pośrednictwem nerwów błędnych — rozkurcz mięśnia
zwieracza bańki wątrobowo-trzustkowej i jednoczesny skurcz błony mięśniowej pęcherzyka żółciowego. Następnie tworząca się w błonie śluzowej
446
Układ trawienny
Tabela 36
Skład żółci w jednym litrze
Związki
Woda
Sucha pozostałość
Kwasy żółciowe
Mucyna i barwniki
żółciowe
Sole nieorganiczne
Tłuszcze i lipidy
Żółć wątrobowa
0,976
24
L
g/L
0,86C
140
g/L
g/L
6
g/L
80
g/L
5
g/L
41
g/L
7—8
3
g/L
g/L
8
19
g/L
g/L
9
1
g/L
g/L
Kwasy tłuszczowe
Tłuszcze obojętne
0,8
0,8
g/L
g/L
Fosfolipidy
0,2
g/L
Cholesterol
Żółć pęcherzykowa
2
SI: 1,56—4,14 mmol/L SI: 15,6
6
0,6 — 1,6
g/L
g/L
mmol/L
g/L
dwunastnicy i krążąca we krwi cholecystokinina (CCK) wywołuje również
skurcz błony mięśniowej pęcherzyka i jego opróżnianie się z żółci
odpływającej do dwunastnicy.
Żółć gromadząca się w pęcherzyku żółciowym ulega kilkakrotnemu
zagęszczeniu dzięki resorpcji wody oraz traci część zasad i jej pH zmniejsza
się. Żółć wytwarzana w wątrobie ma pH około 8,0 (SI: 10 nmol h +/L),
a pH żółci w pęcherzyku żółciowym waha się od 5,0 do 7,4 (SI: 10 39,81
nmol H+/L).
Udział żółci w trawieniu
W ciągu doby wydziela się około 0,5 L żółci. Najważniejszymi jej
składnikami są: sole kwasów żółciowych, barwniki żółciowe, cholesterol
i sole mineralne. W trawieniu w jelicie cienkim współdziałają sole kwasów
żółciowych. Są to sole sodowe i potasowe kwasu cholowego, deoksycholowego i chenodeoksycholowego.
Komórki wątrobowe wydzielają kwasy żółciowe w postaci związanej
z tauryną lub glicyną, jako np. kwasy taurocholowe i glikocholowe.
W przewodach żółciowych następuje tworzenie się odpowiednich soli po
związaniu się tych kwasów z sodem lub potasem.
Trawienie pokarmów
447
Sole kwasów żółciowych wydzielone do światła jelita:
• zmniejszają napięcie powierzchniowe;
• łączą się z produktami lipolizy: kwasami tłuszczowymi o długich
łańcuchach i monoglicerydami;
• aktywują lipazę — enzym hydrolizujący tłuszcze.
Niektóre związki wydzielane do żółci przez komórki wątrobowe są
wchłaniane do krwi w jelicie cienkim. Wchłonięte do krwi powracają przez
żyłę wrotną do wątroby, gdzie są ponownie wychwytywane przez komórki
wątrobowe i wydzielane do żółci.
Do związków kilkakrotnie krążących pomiędzy jelitem cienkim i wątrobą należą sole kwasów żółciowych, cholesterol, bilirubina i urobilinogen. Dzięki temu krążeniu jelitowo-wątrobowemu (entero-hepatic circulation) cząsteczki kwasów żółciowych do 8 razy na dobę krążą pomiędzy
jelitem i wątrobą, biorąc udział w transporcie i wchłanianiu tłuszczów
w jelicie cienkim.
I Jelito grube
Treść jelita cienkiego przedostaje się do jelita ślepego (cecum), części
początkowej jelita grubego {intestinum crassum) w momencie otwierania
się zastawki krętniczo-kątniczej (valva ileocecalis). Zastawka otwiera się
wtedy, kiedy przechodzi przez nią fala perystaltyczna. Występuje to
zwłaszcza w okresie opróżniania się żołądka (odruch żołądkowo-krętniczy). W warunkach fizjologicznych zastawka krętniczo-kątnicza uniemożliwia cofanie się treści z jelita grubego do jelita cienkiego.
W jelicie grubym zachodzą cztery zasadnicze procesy:
• woda zawarta w treści jelita grubego jest zwrotnie wchłaniana;
• wchłaniane są elektrolity, witaminy i aminokwasy;
• formowany jest kał i nie strawione produkty są tam czasowo magazyno
wane;
• drobnoustroje stale się mnożą, wytwarzając zarówno związki dla
organizmu niezbędne (niektóre witaminy), jak i toksyczne (np. tyraminę, amoniak itp.).
Gruczoły błony śluzowej jelita grubego nie wytwarzają enzymów
trawiennych. Jedyną ich wydzieliną jest śluz.
448
Układ trawienny
Ryc. 240. Schemat budowy komórki kubkowej
wydzielającej śluz w błonie śluzowej jelita grubego. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie
PZWL, Warszawa 1997.)
Aparat Golgiego
Jądro
Motoryka jelita grubego
Podobnie jak w jelicie cienkim błona mięśniowa jelita grubego wykazuje okresowe zmiany napięcia, skurcze odcinkowe i skurcze perystaltyczne. Od 2 do 3 razy na dobę, na skutek rozciągnięcia się jelita grubego
przez wypełniającą go treść, dochodzi do silnych skurczów perystaltycznych, czyli ruchów masowych (mass movements). Treść wypełniająca jelito
grube zostaje przesunięta w kierunku odbytnicy (rectum). Ruchy masowe
występują w pierwszej godzinie po spożyciu pokarmów na drodze odruchu
żołądko wo-okrężniczego.
Oddawanie kału
Treść jelita grubego przesuwana do odbytnicy zawiera: wodę, błonnik,
ciała bakterii, sole mineralne, tłuszcze, złuszczone komórki nabłonka jelita cienkiego i jelita grubego oraz śluz. Kał wypełniający odbytnicę
rozciąga jej ściany i może dojść na drodze odruchu do mimowolnego
Trawienie pokarmów
449
oddawania kału, jak to występuje u dzieci w pierwszym roku życia.
W późniejszym okresie życia wykształca się w obrębie wyższych pięter
ośrodkowego układu nerwowego mechanizm kontrolujący oddawanie
kału.
Podrażnienie receptorów w ścianie odbytnicy odbierane jest przez
ośrodek w części krzyżowej rdzenia kręgowego, który z kolei za pośrednictwem przywspółczulnych nerwów trzewnych miednic/nych zmniejsza
napięcie mięśnia gładkiego — mięśnia zwieracza wewnętrznego odbytu
{musculus sphincter ani internus) — i rozkurcza mięsień poprzecznie prążkowany — zwieracz zewnętrzny odbytu {musculus sphincter ani externus) — unerwiany przez nerw sromowy {nervus pudendus).
Jednocześnie z rozluźnieniem obu zwieraczy występują skurcze przepony i mięśni brzucha, ciśnienie w jamie brzusznej wzrasta i kał usuwany
jest na zewnątrz.
Ryc. 241. Jelito grube: s.p. — skurcze perystaltyczne, r.m. — ruchy masowe w czasie
przesuwania uformowanego kału — k.
15 — Fizjologia człowieka
450
Układ trawienny
Ryc. 242. Łuki odruchowe związane z oddawaniem kału. Włókna aferentne nerwu
sromowego zaznaczone są linią ciągłą. Włókna eferentne przedzwojowe i zazwojowe
współczulne oznaczone są linią przerywaną, włókna eferentne przedzwojowe i zazwojowe
przywspołczulne przedstawiono za pomocą linii ciągłej.
I Hormony żołądkowo-jelitowe
Hormony żołądkowo-jelitowe (gastrointestinal hormones) są wydzielane przez komórki rozsianego układu wydzielania wewnętrznego występujące w błonie śluzowej żołądka i jelit. Mają one budowę peptydową
Hormony żo/ądkowo-jelitowe
451
i krążą we krwi, tak jak hormony wydzielane przez gruczoły dokrewne, lub
też działają miejscowo. Wydzielone do światła gruczołów błony śluzowej
żołądka i jelita cienkiego pobudzają lub hamują czynność zewnątrzwydzielniczą tych gruczołów.
Hormony żołądkowo-jelitowe można podzielić na dwie grupy, mając
na uwadze ich budowę chemiczną i właściwości. Jedną grupę stanowi
gastryna i cholecystokinina (CCK), drugą zaś — sekretyna, glukagon,
peptyd glukagonopodobny (czyli glicentyna — GLI), wazoaktywny
peptyd jelitowy (VIP) i peptyd hamujący czynność żołądka (GIP).
Gastryna
Gastryna jest zasadniczym hormonem żołądkowo-jelitowym kontrolującym motorykę żołądka i jelit oraz wydzielanie soku żołądkowego
i jelitowego. Komórki G wewnątrzwydzielnicze błony śluzowej żołądka
i jelit wydzielają trzy peptydy: „dużą" gastrynę — G34 — zbudowaną z 34
aminokwasów, „małą" gastrynę — G17 — zbudowanąz 17 aminokwasów
i „mini" gastrynę — G14 — zbudowaną z 14 aminokwasów. Mają one
identyczną sekwencję aminokwasów karboksylowego końca łańcucha
peptydowego.
Gastryna G34 ma najdłuższy okres połowicznego rozpadu we krwi —
T 1/2 = 15 minut. Gastryna G17 i G14 mają znacznie krótszy okres
połowicznego rozpadu we krwi — TV2 = 2—3 minuty. Z tego względu
ich działanie jest słabsze.
Gastryna jest hormonem pobudzającym najsilniej wydzielanie soku
żołądkowego. W mniejszym stopniu pobudza gruczoły w błonie śluzowej
jelita cienkiego do wydzielania soku jelitowego, komórki zewnątrzwydzielnicze trzustki do wydzielania soku trzustkowego i komórki wątrobowe do
wydzielania żółci. Nasila również motorykę żołądka i jelit oraz skurcze
ścian pęcherzyka żółciowego.
Cholecystokinina
Cholecystokinina (CCK) wytwarzana jest przez komórki I wewnątrzwydzielnicze błony śluzowej dwunastnicy i początkowych odcinków jelita
czczego. Wydzielana jest do krwi w postaci cząsteczek o identycznym
końcu karboksylowym, ale o różnej długości łańcucha peptydowego —
o 8, 12, 33 i 39 aminokwasach. Ostatnie pięć aminokwasów końca
452
Układ trawienny
karboksylowego cholecystokininy jest identyczne z końcem karboksylowym gastryny. Cholecystokinina pobudza wydzielanie soku trzustkowego,
w niniejszym stopniu soku żołądkowego, soku jelitowego i żółci. Silnie
kurczy ścianę pęcherzyka żółciowego i rozkurcza mięsień zwieracz bańki
wątrobowo-trzustkowej, wzmaga perystaltykę jelit, hamuje zaś perystaltykę żołądka.
Tabela 37
Oddziaływanie hormonów żołądkowo-jelitowych na czynność motoryczną i wydzielniczą
żołądka i jelit
Skrót
Liczba
Działanie na
lub
aminożołądek
jelita
Peptyd
symbol
kwasów
motoryka wydzielanie motoryka wydzielanie
Gastryna „mini"
G14
14
+
+++
++
+
Gastryna „mała"
hG17
17
Gastryna „duża"
hG34
34
CCK- 8
CCK-12
CCK-33
CCK-39
8 12
33
39
-
+
GRP
27
+
++
Sekretyna
27
—
pepsynogen + +
HC1 --
Glukagon
29
-
-
+
GLP-1
37
_
—
+
Wazoaktywny
peptyd jelitowy
VIP
28
------
--------
—
++
Peptyd hamujący
czynności żołądka
GIP
42
------
---------
—
++
22
++
+
++
+
14
--------
---------
--------
-
Cholecystokinina
Peptyd uwalniający
gastrynę
Peptyd
glukagonopodobny
Motylina
Somatostatyna
SRIF
+++
+
—
+
Działanie: pobudzające silne 4- 4- +, umiarkowane + 4-, nieznaczne +.
hamujące silne — ---- , umiarkowane ------ , nieznaczne —.
Hormony iolądkowo-jelitowe
453
Tabela 38
Oddziaływanie hormonów żołądkowo-jelitowych na czynność trzustki, wątroby i na
pęcherzyk żółciowy
Peptyd
Skrót
lub
symbol
Działanie na
Liczba
aminokwasów
trzustkę
wydzielanie
zewnętrzne
dokrewne na
komórki
A
Gastryna
„mini"
Gastryna
„mała"
Gastryna
„duża"
+
+
+
+
+
+
+++
G14
14
hG17
17
hG34
34
CCK- 8
CCK-12
CCK-33
CCK-39
8 12
33
39
+++
GRP
27
++
Sekretyna
27
+++
Glukagon
29
Cholecystokinina
Peptyd
uwalniający
gastrynę
Peptyd glukagonopodobny
GLP-1
37
Wazoaktywny
peptyd jelitowy
VIP
28
Peptyd hamujący czynności
żołądka
GIP
42
Motylina
Somatostatyna
B
wątrobę
błonę
wydziela- mięśnionie żółci wą pęcherzyka
żółciowego
+
+
++
+
+
++
+
+
--------
--------
22
SRIF
14
--------
Działanie: pobudzające silne + + +, umiarkowane + +, nieznaczne+
hamujące silne- - -, umiarkowane- -, nieznaczne-
-
-
454
Układ trawienny
Sekretyna
Sekretyna, o łańcuchu peptydowym z 27 aminokwasów, wydzielana jest
do krwi przez komórki S wewnątrzwydzielnicze błony śluzowej dwunastnicy i jelita cienkiego pobudzone przez bardzo kwaśną treść żołądkową
dostającą się do dwunastnicy. Jest hormonem silnie pobudzającym
trzustkę do wydzielania dużych ilości zasadowego soku trzustkowego oraz
wątrobę do wydzielania żółci. Hamuje perystaltykę żołądka i jelit.
Działanie hormonów i przekaźników chemicznych
w przewodzie pokarmowym
Komórki wewnątrzwydzielnicze błony śluzowej dwunastnicy i jelita
cienkiego wydzielają hormony o pewnym podobieństwie w budowie
chemicznej do sekretyny. Są to: glukagon, peptyd glukagonopodobny —
glicentyna (GLI — glucagon-like peptide), wazoaktywny peptyd jelitowy
(VIP — vasoactive intestinal peptide) i peptyd hamujący czynność żołądka
(GIP — gastric inhibitory peptide). Glukagon i glicentyna uczestniczą
w metabolizmie węglowodanów. Wazoaktywny peptyd jelitowy (VIP)
rozszerza naczynia krwionośne w przewodzie pokarmowym, hamuje
motorykę żołądka i wydzialanie soku żołądkowego oraz zwiększa wydzielanie soku jelitowego, trzustkowego i żółci. Peptyd hamujący czynność
żołądka (GIP) hamuje perystaltykę i wydzielanie soku żołądkowego.
W błonie śluzowej przewodu pokarmowego wytwarzane są również:
motylina wzmagająca motorykę i wydzielanie soku żołądkowego oraz
somatostatyna (SRIF) o działaniu przeciwnym — hamującym motorykę
i wydzielanie soków trawiennych. Hormony te działają przede wszystkim
miejscowo — parakrynnie — na komórki sąsiadujące z komórkami
wewnątrzwydzielniczymi.
Motoryka i wydzielanie żołądka i jelit przebiega przy stałym, często
antagonistycznym, oddziaływaniu hormonów żołądkowo-jelitowych wydzielanych
przez
komórki
rozsianego
układu
wydzielania
wewnętrznego i przekaźników chemicznych, czyli transmitterów i
modulatorów, uwalnianych przez neurony przywspółczulne, współczulne
oraz neurony splotów podśluzowego i warstwy mięśniowej jelita.
Transmitterem uwalnianym przez neurony zwojowe przywspółczulne
jest acetylocholina (ACh), przez neurony zwojowe współczulne — noradrenaiina (NA), przez neurony splotów jelita zaś są acetylocholina (ACh),
serotonina (5-HT) i kwas gamma-aminomasłowy (GABA). Neurony te
Wchłanianie w przewodzie pokarmowym
455
uwalniają również modulatory: Substancję P (SP), cholecystokininę
(CCK), dynorfinę (DYN), enkefaliny (ENK), peptyd uwalniający gastrynę
(GRP), wazoaktywny peptyd jelitowy (VIP), peptyd pochodny kalcytoninowego genu (CGRP) i neuropeptyd Y (NPY).
I
Wchłanianie w przewodzie
pokarmowym
Substancje odżywcze, po ich rozłożeniu do związków prostych przez
enzymy soków trawiennych (digestive juices), oraz woda, witaminy i sole
mineralne są wchłaniane do krwi lub chłonki. Mechanizm wchłaniania
polega na dyfuzji lub aktywnym transporcie.
Odcinkiem przewodu pokarmowego, w którym najwięcej wchłania się
składników pokarmowych, jest jelito czcze. Komórki nabłonkowe jelita
czczego z rąbkiem prążkowanym, czyli enterocyty, zachowują się w stosunku do związków wchłanianych tak, jakby miały pory o średnicy
750 pm. Natomiast błona śluzowa jelita krętego zachowuje się tak, jak
gdyby miała mniejsze pory o średnicy 350 pm.
Związki dobrze rozpuszczalne w tłuszczach mogą być wchłaniane
z każdego odcinka przewodu pokarmowego. Cząsteczki o średnicy
mniejszej lub równej średnicy porów i ładunku elektrycznym obojętnym
Ryc. 243. Komórka wchłaniająca jelita
cienkiego. 1 — rąbek prążkowany, 2 — obwódka przylegania, 3 — aparat Golgiego,
4 — ziarnista siateczka śródplazmatyczna,
5 —jądro, 6 — mitochondrium, 7 — błona
podstawna. (Reprodukowano za zezwole
niem Michała Karaska.)
456
Układ trawienny
lub ujemnym dyfundują zgodnie z gradientem stężeń. Do tych związków
należy woda, która wchłania się zgodnie z gradientem ciśnienia osmotycznego, dyfundując przez błonę śluzową w kierunku od niższego do wyższego ciśnienia osmotycznego.
Związki o średnicy większej od średnicy porów wchłaniają się na
zasadzie dyfuzji ułatwionej (facilitated diffusion) lub transportu aktywnego
(active transport) dzięki przejściowemu wiązaniu się z nośnikiem (carrier
substance) znajdującym się w błonie komórkowej enterocytów. Transport
aktywny wymaga zużycia energii i zazwyczaj zachodzi wbrew gradientowi
stężenia. Enterocyty mogą również wchłaniać niektóre duże cząsteczki na
zasadzie pinocytozy.
I Węglowodany
Węglowodany wchłaniają się w postaci monosacharydów, heksoz i pentoz w dwunastnicy i w jelicie czczym. Większość monosacharydów,
a zwłaszcza glukoza i galaktoza, jest wchłaniana na zasadzie transportu
aktywnego. Niektóre z monosacharydów, do których należy fruktoza,
dostają się do krwi dzięki dyfuzji ułatwionej.
Aktywny transport glukozy przyspiesza się w obecności jonów sodowych. Istnieje wzajemna zależność pomiędzy transportem glukozy i jonów sodowych. Wchłanianie jonów sodowych również ulega przyspieszeniu (symport) w obecności glukozy.
Jednorazowe spożycie dużych ilości bisacharydów powoduje ich wchłanianie bez uprzedniej hydrolizy do monosacharydów. Bisacharydy wnikają
do rąbka prążkowanego enterocytów, gdzie zostają zhydrolizowane przez
enzymy tam występujące do monosacharydów. Większość powstających
monosacharydów z enterocytów dyfunduje do krwi, część zaś powraca
do światła jelita.
I Tłuszcze
Tłuszcze spożywane w przeważającej części jako triacyloglicerole są
hydrolizowane w przewodzie pokarmowym do monoacylogliceroli i wolnych kwasów tłuszczowych. Sole kwasów żółciowych tworzą wraz z monoacyloglicerolami i wolnymi kwasami tłuszczowymi agregaty umożliwiające utrzymanie ich w wodzie w postaci roztworu micelarnego.
Wchłanianie w przewodzie pokarmowym
457
Ryc. 244. Schemat budowy błony śluzowej jelita cienkiego. Pomiędzy kosmkami występują
krypty jelitowe, w których zachodzą podziały komórek nabłonka przemieszczających się
w kierunku wierzchołka kosmków, gdzie następuje ich złuszczenie się. (Reprodukowano za
zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa
1997.)
Monoacyloglicerole i wolne kwasy tłuszczowe wnikają do enterocytów.
Monoacyloglicerole i kwasy tłuszczowe o dużej cząsteczce podlegają
w enterocytach estryfikacji do triacylogliceroli. W tych komórkach tworzą się chylomikrony, wraz z którymi triacyloglicerole wydzielane są
następnie do naczyń chłonnych.
Wolne kwasy tłuszczowe o krótkich łańcuchach, nie zawierających
więcej niż 10—12 atomów węgla, dyfundują przez enterocyty i w postaci
nie zmienionej dostają się do krwi. Z krwią przez żyłę wrotną przepływają
do wątroby.
Cholesterol spożywany w pokarmach zostaje w obecności soli kwasów żółciowych zemulgowany i zestryfikowany z wolnymi kwasami
tłuszczowymi. Następnie dostaje się do enterocytów, gdzie tworzy chylomikrony i w tej postaci trafia do naczyń chłonnych.
458
Układ trawienny
TRIACYLOGLICEROLE -
KWASY TłlJSZCIOM
ŚWIATŁO JELITA
Ryc. 245. Wchłanianie tłuszczów przez komórki nabłonka błony śluzowej jelita cienkiego.
FFA — wolne kwasy tłuszczowe.
Triacyloglicerole, cholesterol i fosfolipidy są transportowane za pośrednictwem chłonki do krwi, w postaci kompleksów z białkami, tworząc
kropelki chylomikronu. Krążące we krwi chylomikrony osadzają się na
powierzchni śródbłonka naczyniowego. Występująca tam lipa/a lipoproteinowa hydrolizuje triacyloglicerole do wolnych kwasów tłuszczowych
(FFA) i glicerolu. Związki te wnikają do komórek tłuszczowych, gdzie
zachodzi resynteza triacylogliceroli. Pozostałość chylomikronu jest internalizowana w komórkach wątrobowych.
Wchłanianie w przewodzie pokarmowym
459
Pomiędzy komórkami wątrobowymi i komórkami innych tkanek
zachodzi stała wymiana cholesterolu i triacylogliceroli. Krew transportuje
te związki w postaci lipoprotein pomiędzy wątrobą i tkankami.
Występujące we krwi lipoproteiny o bardzo małej gęstości VLDL (very
Iow density lipoproteins) pochodzą z wątroby. Osadzają się na powierzchni
śródbłonka naczyń krwionośnych, stopniowo tracą triacyloglicerole i zamieniają się w lipoproteiny o pośredniej gęstości: — IDL (intermediate-density lipoproteins), i o małej gęstości — LDL (low-density lipoproteins). W miarę ubywania triacylogliceroli zwiększa się w kompleksach
procentowa zawartość cholesterolu, który zostaje zatrzymywany w komórkach wszystkich tkanek. Natomiast cholesterol pochodzący z komórek wątrobowych występuje w lipoproteinach o dużej gęstości (HDL —
high-density lipoproteins), o małej zawartości triacylogliceroli i cholesterolu oraz o dużej zawartości białek.
Ryc. 246. Wchłanianie z dwunastnicy i jelita cienkiego: cholesterolu, wolnych kwasów
tłuszczowych (FFA) i monoacylogliceroli; ich krążenie w organizmie, osadzanie się na
komórkach śródbłonka, hydroliza pod wpływem lipazy lipoproteinowej i resynteza w komórkach tkanki tłuszczowej — adipocytach. Wytwarzanie przez komórki wątrobowe
(hepatocyty) lipoprotein o bardzo małej gęstości (VLDL), ich krążenie i przekształcanie się
w lipoproteiny o małej gęstości <LDL) i lipoproteiny o dużej gęstości (HDL).
460
Układ trawienny
Tabela 39
Miejsce powstawania i skład procentowy lipoprotein osocza
Średnica Białka
Choleste- TriacyloLipoproteiny
(nm)
rol
glicerole
(%)
(%)
(%)
Chylomikrony
Reszty chylomikronów
2
Fosfolipidy
(%)
Miejsce
powstawania
jelita
śródbłonek
naczyń
włosowatych
5
90
3
8 10
20
55
17
20
30
40 6
20
50
53
5
21
75—1000
30—80
Lipoproteiny o bardzo
małej gęstości (VLDL) 30—80
Lipoproteiny o pośredniej gęstości (IDL)
25-^40
Lipoproteiny o małej
gęstości (LDL)
20 7,5—
Lipoproteiny o dużej
gęstości (HDL)
10
20
wątroba i
jelita
zVLDL
z IDL
wątroba i
jelita
25
Lipoproteiny o małej gęstości (LDL) zawierają dużo cholesterolu
i mało białek. Są utleniane w czasie przechodzenia przez śródbłonek
naczyń tętniczych. Utlenione LDL uszkadzają komórki śródbłonka,
jednocześnie wywołując dodatnią chemotaksję monocytów i makrofagów,
które skupiają się pod komórkami śródbłonka i pożerają utlenione
lipoproteiny LDL, same zamieniając się w komórki piankowate (foam
cells). W miejscu uszkodzenia śródbłonka naczyń tętniczych trombocyty
krążące we krwi przylepiają się tworząc agregaty.
I Białka i kwasy nukleinowe
W jelicie cienkim podlegają wchłanianiu produkty trawienia białek egzogennych, znajdujących się w spożytych pokarmach, oraz białek endogennych, występujących w sokach trawiennych i w złuszczonych komórkach
nabłonka przewodu pokarmowego.
Wchłaniane są tam przede wszystkim za pomocą transportu aktywnego
L-izomery aminokwasów oraz w niewielkich ilościach dwupeptydy i w jeszcze mniejszych oligopeptydy w obecności jonów sodowych, co nosi nazwę
Wchłanianie w przewodzie pokarmowym
461
„pompy aminokwasowej". Wewnątrz komórek nabłonka jelita cienkiego dwupeptydy i oligopeptydy rozkładane są do aminokwasów i w tej
postaci dyfundują do krwi. D-izomery aminokwasów są znacznie wolniej wchłaniane na zasadzie biernej dyfuzji.
Kwasy nukleinowe są rozkładane przez wiele enzymów trawiennych do
pentoz, zasad purynowych i pirymidynowych oraz fosforanów. Zasady
purynowe i pirymidynowe są aktywnie transportowane do komórek jelita
cienkiego.
U osesków część białka jest wchłaniana z przewodu pokarmowego na
drodze pinocytozy. Następnie nie rozłożone cząsteczki białka przenoszone
są w wakuolach przez komórki nabłonka i wydzielane do krwi.
I
Witaminy, sole mineralne
i woda
Witaminy rozpuszczalne w tłuszczach — A, D, E i K — są wchłaniane
z przewodu pokarmowego w obecności tłuszczów oraz żółci. Witaminy
rozpuszczalne w wodzie nie wymagają szczególnych warunków i są szybko wchłaniane do krwi.
Witamina B12 wchłania się wyłącznie w jelicie krętym, tworząc kompleks z czynnikiem wewnętrznym.
Cząsteczki wody dyfundują bez przeszkód przez nabłonek żołądka
i jelita w obu kierunkach zgodnie z gradientem ciśnienia osmotycznego.
W przypadkach treści hipertonicznej woda dyfunduje z krwi do światła
przewodu pokarmowego, aż do wyrównania ciśnienia osmotycznego.
W jelicie grubym jony sodowe są aktywnie wchłaniane i tam też woda
dyfunduje do krwi.
Jony wapniowe, magnezowe i żelazawe (Fe2+) są aktywnie transportowane przez nabłonek górnego odcinka jelita cienkiego. Jony żelazawe
w komórkach błony śluzowej dwunastnicy i jelita czczego wiążą się
z białkiem apoferrytyną i są tam magazynowane w postaci ferrytyny.
W czasie zwiększonej syntezy hemoglobiny jony żelazawe są uwalniane
z nabłonka jelita do krwi, w której krążą związane z białkiem osocza jako
transferryna.
462
Układ trawienny
Ryc. 247. Wchłanianie składników pokarmowych w dwunastnicy, w jelicie czczym, w jelicie
krętym i w jelicie grubym.
Czynność wątroby
463
I Czynność wątroby
Wątroba {hepar) jest największym gruczołem w organizmie o złożonej
czynności. Zbudowana jest z komórek miąższowych, czyli hepatocytów,
stanowiących 80% komórek, z 16% komórek siateczkowo-śródbłonkowych gwiaździstych {reticuloendotheliocytus stellatus), czyli Browicza-Kupffera, i z 4% komórek nabłonka dróg żółciowych.
Hepatocyty są komórkami gruczołowymi zewnątrz- i wewnątrzwydzielniczymi o złożonym metabolizmie. Komórki siateczkowo-śródbłonkowe
gwiaździste wyścielające zatoki żylne w zrazikach wątroby są komórkami
żernymi. Wychwytują z krwi fragmenty komórek, rozpadające się krwinki
czerwone oraz pośredniczą w wymianie składników pomiędzy krwią
przepływającą przez zatoki i hepatocytami.
Ryc. 248. Komórka wątrobowa. S — światło naczynia włosowatego, K — kanalik żółciowy,
P — przestrzeń okołozatokowa, 1 — jądro, 2 — ziarnista siateczka śródplazmatyczna,
3 — aparat Golgiego, 4 — lizosomy, 5 — mikrociałko, 6 — mitochondrium, 7 — ziarna
glikogenu, 8 — gładka siateczka śródplazmatyczna. (Reprodukowano za zezwoleniem
Michała Karaska.)
464
Układ trawienny
Wątroba pełni w organizmie funkcje:
• filtru zarówno dla związków wchłoniętych z przewodu pokarmowego
do krwi, jak i dla związków uwolnionych do krwi w innych układach
i narządach;
• zewnątrzwydzielniczą związaną z powstawaniem żółci i jej wydziela
niem do dwunastnicy;
• wewnątrzwydzielniczą polegającą na wydzielaniu do krwi i chłonki ich
składników;
• magazynu związków niezbędnych do prawidłowego funkcjonowania
organizmu.
Filtr wątrobowy
Krew dopływająca do wątroby w 80% stanowi krew z żyły wrotnej
(yena portae) i w 20% krew tętniczą z tętnicy wątrobowej {arteria hepatica). Krew dopływająca przez żyłę wrotną zawiera składniki wchłonięte
w przewodzie pokarmowym oraz wydzielone do krwi przez narządy, od
których krew odpływa do żyły wrotnej.
Do składników zatrzymywanych przez wątrobę należą monosacharydy, wolne kwasy tłuszczowe, aminokwasy i pozostałości chylomikronów (chylomicron remnants).
Monosacharydy, przede wszystkim glukoza, zatrzymywane są przez
komórki wątrobowe, w obrębie których następuje polimeryzacja cząsteczek glukozy i powstawanie glikogenu wątrobowego.
Wolne kwasy tłuszczowe (FFA) dostają się do komórek wątrobowych,
w których częściowo zamieniane są na tłuszcze obojętne — triacyloglicerole — i częściowo są przekształcane na acetylo-koenzym A. Następnie cząsteczki acetylo-koenzymu A podlegają kondensacji i powstaje
aceto-acetylo-koenzym A. Z kolei jest on rozkładany do zredukowanego
koenzymu A i uwalnianych do krwi ciał ketonowych: kwasu acetylooctowego, beta-hydroksymasłowego i acetonu.
Komórki wątrobowe odznaczają się zdolnością wychwytywania aminokwasów. Stężenie aminokwasów w komórkach wątrobowych może być do
dziesięciu razy większe w porównaniu z ich stężeniem w osoczu.
Hepatocyty wychwytują z krwi dopływającej przez żyłę wrotną bilirubinę, wydzieloną z żółcią do dwunastnicy i zwrotnie wchłoniętą w jelicie
cienkim, oraz inne związki powstające w jelitach, jak skatol, indol itp.
Wątroba stanowi także filtr dla związków wytwarzanych w innych
tkankach, poza przewodem pokarmowym. Stale wychwytywane są z krwi
Czynność wątroby
465
amoniak, glutamina i kwas glutaminowy powstające we wszystkich tkankach w procesie deaminacji białek. Z amoniaku lub z glutaminy i kwasu
glutaminowego syntetyzowany jest w wątrobie mocznik uwalniany następnie do krwi.
Bilirubina krążąca we krwi jako katabolit hemu, czyli bilirubina nie
związana, pośrednio reagująca z odczynnikiem dwuazowym, jest zatrzymywana w wątrobie, wiązana z kwasem glukuronowym lub kwasem
siarkowym i wydzielana z żółcią. Część związanej z kwasami bilirubiny
trafia z wątroby do krwi i krąży w niej jako tzw. bilirubina bezpośrednio
reagująca z odczynnikiem dwuazowym.
Wątroba stanowi również filtr zatrzymujący większość krążących we
krwi hormonów i w ten sposób pośrednio reguluje ich stężenie we krwi.
Ryc. 249. Krążenie bilirubiny pomiędzy komórkami wątrobowymi, kanalikami żółciowymi
(k.ż.), przewodami żółciowymi (p.ż.), dwunastnicą i jelitem cienkim oraz krwią żyły wrotnej.
466
Układ trawienny
Hormony polipeptydowe są rozkładane do aminokwasów, a hormony
pochodne cholesterolu metabolizowane i wiązane z kwasem glukuronowym. Glikokortykoidy i testosteron metabolizowane są w wątrobie do
17-ketosteroidów i w tej postaci wydalane z moczem. Mineralokortykoidy,
estrogeny, progesteron i ich pochodne ulegają związaniu z kwasem
glukuronowym i w postaci glukuronianów są wydalane z moczem.
Estrogeny, glikokortykoidy i bilirubina (bezpośrednio reagująca) związane z kwasem glukuronowym są również wydzielane wraz z żółcią
i w jelitach zwrotnie wchłaniane, krążąc pomiędzy wątrobą i jelitem
cienkim w obrębie tzw. krążenia jelitowo-wątrobowego (enterohepatic
circulation).
I Czynność zewnątrzwydzielnicza
Komórki wątrobowe tworzą ściany kanalików żółciowych i wydzielają do ich światła wszystkie składniki żółci, w tym również związane
z kwasem glukuronowym: bilirubinę, biliwerdynę, hormony kory nadnerczy i gruczołów płciowych oraz ich pochodne. Pozostałymi składnikami
żółci są: sole kwasów żółciowych, cholesterol, kwasy tłuszczowe, tłuszcze
obojętne, sole nieorganiczne i woda (patrz tab. 36, str. 446)
I Czynność wewnątrzwydzielnicza
Białka osocza krwi, a więc albuminy, alpha- i beta-globuliny oraz
fibrynogen, syntetyzowane są w wątrobie. Jedynie gamma-globuliny nie są
syntetyzowane w wątrobie, lecz w układzie limfoidalnym, przede wszystkim w węzłach chłonnych. Komórki wątrobowe wydzielają również do
krwi enzymy osocza oraz czynniki krzepnięcia krwi.
I Magazynowanie
Składnikiem magazynowanym w największych ilościach w komórkach
wątrobowych jest glikogen. Powstaje on w wyniku kondensacji wchłoniętej
w jelitach glukozy oraz w znacznie mniejszym stopniu z glukozy resyntetyzowanej z krążącego we krwi kwasu mlekowego.
Czynność wątroby
467
Zwiększenie stężenia glukozy we krwi sprzyja magazynowaniu się
glikogenu w wątrobie. Przeciwnie — zmniejszenie stężenia glukozy we
krwi powoduje glikolizę i przechodzenie glukozy z wątroby do krwi. Na tej
zasadzie wątroba utrzymuje stałe stężenie glukozy we krwi, co jest
nazywane „glukostatem wątrobowym" (hepatic glucostate).
Duża zawartość glikogenu w wątrobie hamuje deaminację aminokwasów. Aminokwasy wchłonięte z przewodu pokarmowego nie podlegają deaminacji i są zużywane do syntezy białek w wątrobie. W wątrobie
magazynowane są witaminy rozpuszczalne w tłuszczach: A, D, E i K.
Ryc. 250. Biosynteza w komórkach wątrobowych białek osocza, tłuszczów obojętnych
i mocznika, k.ż. — kanaliki żółciowe, p.ż. — przewodziki żółciowe.
468
Układ trawienny
Żelazo magazynowane w organizmie w postaci ferrytyny znajduje się
w komórkach błony śluzowej jelita cienkiego i w wątrobie. Zmniejszenie
stężenia żelaza krążącego w osoczu w postaci transferryny powoduje
przechodzenie jonów żelaza z obu magazynów do osocza.
I Regulacja czynności
Czynność wątroby uzależniona jest od przepływu krwi oraz od
związków zmieniających metabolizm komórek wątrobowych. Przepływ
krwi i metabolizm są ze sobą ściśle związane. Przepływ krwi przez wątrobę
oznaczany jest u ludzi metodą pośrednią na podstawie obliczonego
„wskaźnika oczyszczania" krwi przepływającej przez wątrobę lub zatrzymywania się w niej gazów obojętnych (np. ksenonu) na skutek ich
dyfuzji w wątrobie. Do tych oznaczeń niezbędna jest kateteryzacja żył
wątrobowych.
Z różnicy tętniczo-żylnej w zawartości składnika zatrzymywanego
przez wątrobę uzyskuje się tzw. obliczony przepływ krwi przez wątrobę —
EHBF (estimated hepatic blood flow). U ludzi zdrowych EHBF wynosi
około 1500 mL krwi na minutę.
Unerwienie naczynioruchowe ma niewielki wpływ na objętość krwi
przepływającej przez wątrobę w ciągu minuty. Nerw błędny nie ma wpływu na przepływ krwi, natomiast pod wpływem impulsów z nerwów
trzewnych ulegają zwężeniu naczynia krwionośne w wątrobie, wzrasta
ciśnienie w żyle wrotnej i zmniejsza się przepływ krwi.
Zasadniczy wpływ na przepływ krwi ma pozycja ciała i praca fizyczna.
W pozycji siedzącej EHBF zmniejsza się o 20%, w stojącej o 40%,
a w czasie pracy fizycznej o 80—85%. Wraz ze zmniejszeniem przepływu
krwi przez wątrobę zmniejsza się prężność tlenu P 02 w komórkach
wątrobowych i zwalniają się wszystkie procesy wymagające stałego
dopływu tlenu.
Przepływ krwi przez wątrobę znacznie się zwiększa pod wpływem
sekretyny i wchłoniętych w jelitach kwasów żółciowych, a w mniejszym
stopniu pod wpływem adrenaliny, glukagonu i insuliny, która wtórnie
wywołuje wydzielanie się adrenaliny. Przepływ krwi przez wątrobę zmniejsza wazopresyna i w stopniu nieznacznym — noradrenalina.
Hormony krążące we krwi i wpływające na metabolizm węglowodanów, białek i tłuszczów zmieniają odpowiednio funkcję filtracyjną, zewnątrzwydzielniczą i wewnątrzwydzielniczą wątroby.
Płyny ustrojowe i
czynność nerek
I Płyny i elektrolity ustrojowe I
Woda
Woda jest składnikiem ustrojowym stanowiącym ponad połowę masy
ciała. U kobiet na wodę przypada przeciętnie 52%, a u mężczyzn 63%
masy ciała. Odchylenie od przeciętnego procentowego udziału wody
w masie ciała zależy od stopnia rozwoju tkanki tłuszczowej. U ludzi otyłych
procent masy ciała przypadający na wodę jest mniejszy. Tkanka tłuszczowa zawiera tylko około 10% wody, tkanka kostna około 22%,
a pozostałe tkanki miękkie od 68 do 82%.
Całkowita woda organizmu — TBW (total body water) — występuje
w trzech zasadniczych przestrzeniach (body fluid compartments) wypełnionych:
• płynem wewnątrzkomórkowym — ICF (intracellular fluid);
• płynem zewnątrzkomórkowym — ECF (extracellular fluid);
• płynem transkomórkowym — TCF (transcellular fluid).
Płyn w przestrzeni wewnątrzkomórkowej (ICF) stanowi ponad połowę
wody znajdującej się w organizmie, co odpowiada 30 — 40% masy ciała.
Pozostałe 23% przypada na wodę wchodzącą w skład płynu przestrzeni
zewnątrzkomórkowej (ECF) i transkomórkowej (TCF).
Płyn w przestrzeni zewnątrzkomórkowej (ECF) stanowi osocze krwi,
płyn tkankowy i chłonka.
Płyn w przestrzeni transkomórkowej (TCF) obejmuje płyn mózgowo-rdzeniowy, płyn w komorach oka, płyn w jamach ciała (opłucnej,
470
Płyny ustrojowe i czynność nerek
osierdziu, otrzewnej), płyn w torebkach stawowych i soki trawienne
znajdujące się w przewodzie pokarmowym (ślina, sok żołądkowy, sok
trzustkowy, żółć i sok jelitowy).
Stosunek objętości płynu w przestrzeni wewnątrzkomórkowej do
objętości płynu w przestrzeni zewnątrzkomórkowej i przestrzeni transkomórkowej podlega zmianom w zależności od ilości wypijanej wody i od
ilości soli mineralnych wprowadzonych do organizmu wraz z pokarmami
oraz od utraty wody przez organizm. Ze względów praktycznych płyn
w przestrzeni zewnątrzkomórkowej i transkomórkowej określa się łącznie
jako płyn zewnątrzkomórkowy — ECF (extracellular fluid).
Po wypiciu płynu hipotonicznego zwiększa się objętość płynu zewnątrzkomórkowego i wewnątrzkomórkowego. Towarzyszy temu obniżenie się
ciśnienia osmotycznego obu płynów.
Wypicie płynu hipertonicznego lub jego dożylna infuzja zwiększa
objętość płynu zewnątrzkomórkowego, zmniejszając jednocześnie objętość
płynu wewnątrzkomórkowego. Wzrasta ciśnienie osmotyczne obu płynów.
Płyny izotoniczne wypijane lub wprowadzane bezpośrednio dożylnie,
np. jako krew konserwowana, zwiększają tylko objętość ECF.
Ryc. 251. Stosunek objętości płynu i ciśnienia osmotycznego elektrolitów w przestrzeni
wewnątrzkomórkowej (I) do objętości płynu i ciśnienia osmotycznego elektrolitów w przestrzeni zewnątrzkomórkowej (E): a — po wypiciu płynu hipotonicznego; b — po wypiciu
płynu hipertonicznego; c — po wypiciu płynu izotonicznego; d — w wyniku utraty chlorku
sodowego.
Utrata jonów nieorganicznych w czasie długotrwałych wymiotów lub
w czasie biegunki zmniejsza objętość płynu zewnątrzkomórkowego i zwiększa objętość płynu wewnątrzkomórkowego. Ciśnienie osmotyczne obu
płynów obniża się.
Płyny i elektrolity ustrojowe
471
I Elektrolity
Sód
W organizmie dorosłego człowieka znajduje się około 60 mmol sodu
w przeliczeniu na 1 kg masy ciała. Ponad połowa sodu w organizmie
znajduje się w płynie zewnątrzkomórkowym. Sód występujący w płynach
ustrojowych zewnątrz- i wewnątrzkomórkowym oraz częściowo w kościach stanowi tak zwany sód wymienialny (exchangeable), w ilości około 42
mmol/kg masy ciała. Pozostałe 18 mmol/kg masy ciała stanowi sód
niewymienialny (nonexchangeable) związany w tkance kostnej.
Potas
Potas występuje prawie wyłącznie w płynie wewnątrzkomórkowym
w ilości około 42 mmol/kg masy ciała. Prawie cały potas występuje
w organizmie w postaci wymienialnej. W osoczu znajduje się on w ilości
około 4 mmol/L. Podwojenie się stężenia potasu w osoczu może być
przyczyną śmierci. Zwiększenie zawartości jonów potasu w płynie zewnątrzkomórkowym powoduje utratę pobudliwości komórek mięśniowych, zwłaszcza mięśnia sercowego i komórek nerwowych.
Wapń i magnez
W organizmie dorosłego człowieka znajduje się około 27,5 mol wapnia,
czyli około 400 mmol wapnia w przeliczeniu na 1 kg masy ciała. W 99%
jest to wapń zmagazynowany w tkance kostnej. Pozostałe 1% wapnia
znajduje się w płynie zewnątrzkomórkowym (ECF) i w komórkach.
Całkowita zawartość wapnia, wolnego i związanego z białkami osocza,
wynosi około 2,5 mmol/L osocza. Z albuminami i globulinami osocza
związane jest 1,16 mmol/L wapnia. Wapń zjonizowany występuje w osoczu
w stężeniu 1,18 mmol/L, związany z anionami zaś — 0,16 mmol/L.
Prawidłowe stężenie wapnia zjonizowanego w osoczu ma istotne znaczenie dla czynności wszystkich komórek, szczególnie dla komórek tkanek
pobudliwych i komórek gruczołowych. Jony Ca 2+, przechodząc do
cytoplazmy z płynu zewnątrzkomórkowego lub ze zbiorników wewnątrzkomórkowych, są drugim przekaźnikiem informacji wewnątrzkomórkowej dzięki aktywacji wielu enzymów. Ruch komórek i egzocytoza
472
Płyny ustrojowe i czynność nerek
pęcherzyków cytoplazmatycznych, w tym również pęcherzyków synaptycznych, zależą od stężenia jonów Ca 2+.
Zmagazynowane w tkance kostnej 99% wapnia organizmu tworzy dwie
pule:
• pulę wapnia łatwo wymienialnego (readily exchangeable pool);
• pulę wapnia niewymienialnego (stable pool).
Wapń łatwo wymienialny w ilości około 500 mmol wymienia się
w ciągu doby pomiędzy tkanką kostną i płynem zewnątrzkomórkowym
(ECF). Wapń z puli niewymienialnej w tkance kostnej również podlega
wymianie, ale znacznie wolniej. U ludzi dorosłych połowa wapnia z tej puli
wymienia się w ciągu 3—5 lat.
U dorosłego człowieka w całym organizmie znajduje się około 1 mol
magnezu. Z tego połowa występuje w tkance kostnej, druga połowa zaś
w płynach ustrojowych: zewnątrzkomórkowym (ECF) i wewnątrzkomórkowym (ICF). Magnez występuje w osoczu w stężeniu o połowę mniejszym
(0,8—1,3 mmol/L) w stosunku do wapnia. W płynie wewnątrzkomórkowym (ICF) zaś stężenie magnezu jest około dziesięć razy większe niż
stężenie w osoczu.
Tabela 40
Czynniki działające na komórki tkanki kostnej
Czynniki aktywujące osteoblasty
Czynniki aktywujące osteoklasty
Parathormon (PTH) 1,25dihydroksycholekalcyferol (1,25-DHD3)
Trijodotyronina (T 3), tyroksyna (T 4)
Hormon wzrostu (hGH) i czynnik
wzrostowy insulinopodobny I (IGF-I)
Prostaglandyny grupy E2 (PGE2) Czynnik
wzrostowy transformujący beta (TGFp)
Estrogeny
Parathormon (PTH) 1,25dihydroksycholekalcyferol (1,25-DHD3)
Interleukina 1 (IL-1) i interleukina 6 (IL-6)
Czynnik martwicy nowotworów (TNF)
Czynnik wzrostowy transformujący alpha
(TGFa)
Czynniki hamujące osteoblasty
Czynniki hamujące osteoklasty
Hormony kory nadnerczy
Kalcytonina Estrogeny zmniejszające
tworzenie się interleukiny 6 (IL-6)
Czynnik wzrostowy transformujący beta
(TGFP) Interferon alpha (IFNa)
Prostaglandyny grupy E 2 (PGE2)
Płyny i elektrolity ustrojowe
473
Jony magnezowe współzawodniczą z jonami wapniowymi, działając
antagonistycznie w stosunku do nich.
Jony magnezowe hamują:
• aktywność ATP-azową miozyny;
• uwalnianie jonów wapniowych ze zbiorników siateczki sarkoplaz matycznej;
• otwieranie kanałów potasowych w błonie komórkowej i otwieranie
kanałów wapniowych poprzez receptory NMDA dla aminokwasów
pobudzających.
Chlor i wodorowęglany
W organizmie człowieka znajduje się średnio około 33 mmol jonów
chlorkowych na 1 kg masy ciała. W 70% znajdują się w osoczu i w płynie
tkankowym. Pozostałe 30% występuje w tkance łącznej we włóknach
kolagenowych oraz w komórkach. Spośród wszystkich komórek w organizmie najwięcej jonów chlorkowych zawierają erytrocyty.
Anionów wodoroweglanowych jest w organizmie średnio 10—12 mmol
na 1 kg masy ciała. Ich zawartość w płynach ustrojowych waha się.
Zwiększenie zawartości jonów wodorowych w organizmie powoduje
tworzenie się z wodorowęglanów kwasu węglowego, który następnie
rozpada się na usuwany przez płuca dwutlenek węgla i na wodę. Przeciwnie — alkalizacja płynów ustrojowych wywołuje zwiększenie zawartości
anionów wodorowęglanowych w organizmie.
I Utrata wody i elektrolitów
Organizm człowieka traci wodę:
•
•
•
•
•
wraz z moczem wydalanym przez nerki;
wraz z potem wydzielanym przez gruczoły potowe;
z powierzchni skóry w wyniku parowania;
wraz z kałem przez przewód pokarmowy;
przez płuca z powietrzem wydychanym.
Najwięcej wody i elektrolitów, około 1,5 L na dobę, organizm traci wraz
z moczem. Utrata wody na skutek parowania powierzchni skóry i wydzielania gruczołów potowych zależy od temperatury własnej ciała i otoczę-
474
Płyny ustrojowe i czynność nerek
nia oraz wilgotności powietrza. W temperaturze nie wyższej niż +28°C
organizm traci w ciągu doby około 500 mL wody wydzielanej z potem
przez parowanie z powierzchni skóry.
Utrata wody przez płuca zależy również od temperatury własnej ciała
i otoczenia oraz od wilgotności powietrza. W warunkach przeciętnych
człowiek traci w ten sposób około 300 mL wody na dobę.
Z kałem wydalane jest z organizmu 100 — 200 mL wody na dobę.
I Czynność nerek
Jednostką funkcjonalną nerki (ren) jest nefron składający się z ciałka
nerkowego (corpusculum renale) i kanalików nerkowych (tubuli renales).
W ciałku nerkowym zachodzi filtracja kłębuszkowa, czyli przenikanie
części osocza przepływającego przez kłębuszek naczyniowy ciałka nerkowego {glomerulus corpusculi renalis) do światła torebki kłębuszka (lumen
capsulae glomeruli). Ultrafiltrat gromadzący się w świetle torebki kłębuszka kolejno przepływa przez część bliższą kanalika nerkowego (pars
proximalis tubuli nephroni), składającą się z kanalika nerkowego głównego
(tubulus renalis cardinalis) i z części zstępującej pętli nefronu (pars
descendens ansae nephroni) i część dalszą kanalika nerkowego (pars distalis
tubuli nephroni) utworzoną z części wstępującej pętli nefronu (pars
ascendens ansae nephroni) i z wstawki (pars intercalata). Mocz tworzący się
w nefronie odpływa przez kanalik nerkowy zbiorczy (tubulus renalis
colligens) i przewód brodawkowy (ductus papillaris) do miedniczki nerkowej (pehis renalis).
W każdej nerce człowieka znajduje się około 1 miliona nefronów, czyli
w obu nerkach około 2 milionów. Prawidłowa czynność wydalnicza nerek
jest zachowana wtedy, kiedy nie mniej niż 30% nefronów, czyli 600 tysięcy nefronów, jest w pełni sprawna.
Czynność nerek wiąże się z:
• tworzeniem moczu, z którym usuwane są z organizmu metabolity
i nadmiar płynów i elektrolitów, czyli z czynnością zewnątrzwydzielniczą;
• wydzielaniem do krwi związków biologicznie czynnych, co jest czynnoś
cią wewnątrzwydzielniczą.
W tworzeniu się moczu udział biorą wszystkie odcinki nefronu oraz
kanaliki nerkowe zbiorcze.
Czynność nerek
475
Ciałko nerkowe korowe
Tętniczka
doprowadzająca
Przewód brodawkowy
Tętniczka odprowadzająca
Włosowate naczynia Plamka
gęsta
Ciałko nerkowe przyrdzeniowe
Kanalik U rzędu Kanalik I rządu
Kanalik zbiorczy
Pętla Henlego:
/ramię zstępujące
,ramię wstępujące
Naczynia proste
Ryc. 252. Schemat budowy nefronów: a — korowych o
krótkiej pętli nefronu (Henlego); b — o długiej pętli
nefronu. (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL,
Warszawa 1997.)
476
Płyny ustrojowe i czynność nerek
Ryc. 253. Schemat budowy ciała nerkowego. (Reprodukowano za zezwoleniem z W. Sawicki
Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Tworzenie się moczu związane jest z trzema procesami:
filtracją kłębuszkową;
resorpcją kanalikową;
sekrecją kanalikową.
I Przepływ krwi
Przez nerki przepływa w spoczynku 1,08 L/min (SI: 18 mL/s) krwi,
co stanowi 20% pojemności minutowej serca. Do kłębuszków krew
dopływa przez tętniczki doprowadzające (arterioli glomerulares afferenś)
i odpływa przez tętniczki odprowadzające (arterioli glomerulares efferens).
Czynność nerek
Ali
Ciśnienie w naczyniach kłębuszka jest wyższe w porównaniu do ciśnienia
w naczyniach włosowatych w innych narządach i wynosi 6 kPa przy
średnim ciśnieniu w zbiorniku tętniczym dużym (Ra s) równym 13,3 kPa.
Komórki błony mięśniowej tętnic nerkowych kurczą się pod wpływem
impulsacji przewodzonej przez współczulne nerwy nerkowe i uwalnianą
noradrenalinę z zakończeń tych nerwów. Skurcz błony mięśniowej tętnic
zmniejsza przepływ krwi przez nerki i przez naczynia kłębuszków nerkowych.
Przepływ krwi przez naczynia kłębuszków zależy również od przekaźników chemicznych i hormonów dopływających z krwią tętniczą do
kłębuszków. Hormony i przekaźniki działają na komórki mezangium
{mesangiocytus) występujące w kłębuszkach nerkowych i otaczające od
zewnątrz naczynia włosowate kłębuszków. Skurcz komórek mezangium
zmniejsza przepływ krwi przez naczynia włosowate kłębuszków.
Komórki mezangium kurczą się pod wpływem dopływających z krwią
tętniczą: noradrenaliny. hormonów peptydowych — endoteliny, wazopresyny (AVP), angiotensyny II (Ang. II), eikozanoidów — tromboksanu
A2 (TxA2), prostaglandyny F 2 (PGF ą), leukotrienów C 4 (LTC 4) i D4 (LTD
). Na komórki mezangium działają rozkurczająco, zwiększając
Ryc. 254. Schemat budowy ściany naczynia krwionośnego włosowatego kłębuszka nerkowego. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo
Lekarskie PZWL, Warszawa 1997.)
478
Płyny ustrojowe i czynność nerek
przepływ kłębuszkowy krwi: dopamina, przedsionkowy peptyd natriuretyczny (ANP) i prostaglandyny E 2 (PGE2).
Niektóre związki wprowadzone do osocza krwi, np. kwas para-aminohipurowy (PAH), przepływając przez nerki zostają w nich prawie całkowicie wychwytywane i wydalane z moczem. Osocze krwi przepływającej
przez nerki prawie całkowicie „oczyszcza się" z tych związków.
Klirens nerkowy PAH (renal clearance of PAH) oblicza się wg wzoru:
CpAH —
uP
,xV
gdzie:
CPAH — klirens dla kwasu para-aminohipurowego (PAH),
UPAH — stężenie PAH w moczu w mg/mL moczu,
V
— objętość moczu wydalana przez nerki w ciągu 1 minuty,
PPAH — stężenie PAH w osoczu krwi w mg/mL osocza.
Ryc. 255. Budowa kanalika nerkowego: a — przekrój poprzeczny; b — fragment powiększony. 1 — część bliższa kanalika nerkowego, 2 — ramię zstępującej pętli nefronu,
3 — część dalsza kanalika nerkowego. (Reprodukowano za zezwoleniem Michała Karaska.)
Czynność nerek
479
Na podstawie klirensu nerkowego PAH można obliczyć efektywny
przepływ osocza przez czynny miąższ nerkowy. Tym samym klirens
nerkowy PAH równy jest efektywnemu przepływowi osocza przez nerki —
ERPF (effective renal plasma flow).
Efektywny przepływ osocza przez nerki (ERPF), obliczany na podstawie klirensu dla kwasu para-aminohipurowego, wynosi około 625
mL/min (SI: 10,41 mL/s). Znając wartość ERPF oraz wskaźnik hematokrytu (Het), łatwo obliczyć efektywny przepływ krwi przez nerki — ERBF
(effective renal blood flow).
ERBF = C
100-Het
Część krwi (około 10%) przepływającej przez nerki pomija nefrony,
przez co nie podlega oczyszczeniu np. z PAH. Dlatego też całkowity
przepływ krwi przez nerki — TRBF (total renal blood flow) jest średnio
o 10% większy od wartości ERBF.
Całkowity przepływ krwi przez nerki (TRBF) można obliczyć, znając
wartość ERBF oraz stężenie PAH we krwi tętniczej (a) i żylnej (v) nerek,
posługując się wzorem:
TRBF = ERBF:
apAH VpAH
~
=
Filtracja kłębuszkowa
Część osocza krwi przepływającej przez naczynia włosowate kłębuszków nerkowych ulega przefiltrowaniu do światła torebki kłębuszka.
W naczyniach włosowatych kłębuszka panuje ciśnienie filtracyjne około
1,4 kPa (6,0 kPa ciśnienie hydrostatyczne — 3,3 kPa ciśnienie onkotyczne — 1,3 kPa ciśnienie w świetle torebki kłębuszka) i dzięki temu ciśnieniu
filtracyjnemu 1/5 część osocza przepływająca przez nerki zostaje przefiltrowana.
Wielkość filtracji kłębuszkowej — GFR (glomerular filtration ratę) —
można oznaczyć u człowieka, wprowadzając do krwi substancje nie
podlegające resorpcji lub sekrecji kanalikowej. Taką substancją egzogenną, nie wchłaniającą się i nie wydzielaną w kanalikach, jest wielocukier — inulina, która ma masę cząsteczkową 5,5 kDa.
Nerkowy klirens dla inuliny — C in (clearance of inulin) — jest
odzwierciedleniem wielkości filtracji kłębuszkowej (GFR).
480
Płyny ustrojowe i czynność nerek
.
U,„xV
K~
gdzie:
Cin — klirens dla inuliny (in), w mL/min,
Ujn — stężenie inuliny w moczu w mg/mL moczu,
V — objętość moczu wydalona przez nerki w ciągu 1 minuty,
Pin — stężenie inuliny w osoczu w mg/mL osocza.
W ciągu 1 minuty powstaje u mężczyzn o przeciętniej budowie ciała
125 mL/min ultrafiltratu, u kobiet zaś o 10% mniej.
GFR u mężczyzn=125±15 mL/min/1,75 m2 powierzchni ciała (SI:
2,08 + 0,25 mL/s/1,75 m2).
GFR u kobiet=110±15 mL/min/1,75 m 2 powierzchni ciała (SI:
1,83±0,25 mL/s/1,75 m2).
W ciągu doby tworzy się w kłębuszkach nerkowych u mężczyzn 180 L
przesączu kłębuszkowego, czyli ultrafiltratu.
Ryc. 256. Rozkład ciśnień w naczyniach krwionośnych włosowatych kłębuszka naczyniowego
ciałka nerkowego. Ciśnienie hydrostatyczne w naczyniu krwionośnym włosowatym (1)
i w świetle torebki kłębuszka (3) nie zmienia się w czasie przepływu krwi przez naczynie
krwionośne włosowate kłębuszka. W wyniku filtracji kłębuszkowej wzrasta ciśnienie
onkotyczne osocza (2) i zmniejsza się ciśnienie filtracyjne (4).
Przepuszczalność naczyń kłębuszka
Filtracja w kłębuszkach nerkowych przebiega tak, jak gdyby pory
w ścianie naczyń włosowatych kłębuszka miały średnicę 10 nm. W prze-
Czynność nerek
481
Ryc. 257. Schemat budowy dwóch nefronów. 1 — tętniczka doprowadzająca, 2 — tętniczka
odprowadzająca: a — przez nefron przepływa krew zawierająca egzogenny polisacharyd inulinę, która filtruje się w kłębuszkach nerkowych i nie jest resorbowana ani wydalana
w kanalikach nerkowych; b — przez nefron przepływa krew zawierająca glukozę, która
jest filtrowana w kłębuszkach nerkowych i jest całkowicie wchłaniana w kanalikach
nerkowych aż do osiągnięcia progu maksymalnej wchłanialnosci.
16 — Fizjologia człowieka
482
Pfyny ustrojowe i czynność nerek
sączu kłębuszkowym występują związki o masie cząsteczkowej poniżej
70 kDa. Nieznaczne ilości albumin filtrują się do przesączu kłębuszkowego
i w częściach bliższych kanalików nerkowych są wchłaniane na zasadzie
pinocytozy.
Hemoglobina uwolniona z erytrocytów w czasie zwiększonej hemolizy
również filtruje się w kłębuszkach nerkowych i występuje w moczu.
Frakcja filtracyjna
Frakcja filtracyjna (filtration fraction) jest to stosunek wielkości filtracji
kłębuszkowej (GFR) do efektywnego przepływu przez nerki osocza
(ERPF) i wynosi u mężczyzn od 0,12 do 0,20 (SI: volfr. 0,12—0,20
mL/mL), średnio około 0,18.
GFR 125 mL/min
Frakcja filtracyjna = ————— ' , =0,18
ERPF 625 mL/min
I Resorpcja i sekrecja kanalikowa
Resorpcja kanalikowa (tubular resorption) i sekrecja kanalikowa
(tubular secretion) są to zasadnicze funkcje części bliższych i dalszych
kanalików nerkowych.
Związki chemiczne krążące we krwi i przefiltrowane w kłębuszkach
nerkowych do przesączu kłębuszkowego następnie przepływają przez
kanaliki nerkowe, gdzie:
• nie są wchłaniane w kanalikach ani wydzielane przez nabłonek kanali
ków, np. inulina;
• zostają wchłaniane całkowicie lub częściowo, np. glukoza;
• zostają dodatkowo wydzielane przez nabłonek kanalików, np. kwas
para-aminohipurowy;
• są jednocześnie wchłaniane i wydzielane, np. jony potasowe.
Związki należące do pierwszej grupy występują w moczu ostatecznym
w tej samej ilości, w jakiej zostały przefiltrowane w kłębuszkach. Dzięki
temu klirens dla tych związków wskazuje na wielkość filtracji kłębuszkowej.
Czynność nerek
483
Resorpcja kanalikowa
Resorpcja kanalikowa ma charakter:
• resorpcji biernej, zgodnej z gradientem stężenia i potencjału elektrycz
nego;
• resorpcji czynnej zachodzącej wbrew gradientom.
Do związków, które są w kanalikach wchłaniane biernie wraz z wodą,
należą mocznik i jony chlorkowe.
Związki resorbowane czynnie dzielą się na związki, których maksymalna wchłanialność ograniczona jest:
• stężeniem;
• stężeniem i czasem wchłaniania.
Związki pierwszej grupy są aktywnie transportowane przez nabłonek
kanalików nerkowych proporcjonalnie aż do określonego ich stężenia
w przesączu kłębuszkowym. Największe stężenie tych związków w przesączu kłębuszkowym, przy którym ulegają one jeszcze całkowitej resorpcji
w kanalikach nerkowych, określa wartość Tm (transport maximum).
Wartość Tm określa tę maksymalną ilość danej substancji, jaka może być
zresorbowana w ciągu jednej minuty.
Glukoza jest związkiem transportowanym aktywnie przez błonę komórkową nabłonka kanalika. Przekroczenie stężenia glukozy w osoczu ponad
10,0 mmol/L, z czym wiąże się zwiększenie jej stężenia w przesączu
kłębuszkowym, jest równoznaczne z przekroczeniem Tm dla niektórych
nefronów. Wówczas w moczu pojawia się glukoza. Progowe stężenie
glukozy w osoczu, przy którym pojawia się ona w moczu, nosi nazwę progu
nerkowego dla glukozy (renal threshold).
Poza glukozą wchłaniane są aktywnie w kanalikach nerkowych z udziałem nośnika: kationy potasowe, aniony fosforanowe i siarczanowe,
aminokwasy, kreatyna, kwas moczowy, kwas askorbinowy, kwas acetylooctowy i ciała ketonowe.
Do związków resorbowanych czynnie o największej wartości Tm
(proporcjonalnej wchłanialności ograniczonej stężeniem i czasem wchłaniania) zalicza się jony sodowe.
Sekrecja kanalikowa
Przez komórki nabłonka wydzielanych jest do światła kanalików wiele
związków endogennych i egzogennych. Wydzielanie każdego z tych
związków odbywa się na zasadzie jednego z trzech mechanizmów:
484
Płyny ustrojowe i czynność nerek
Błona podstawną
------ Dyfuzja
^—^— Transport aktywny
O mV
Na+
SWIATkO
KANALIKA
BLIŻSZEGO
KOMÓRKA NABkONKA
KANALIKA BLIŻSZEGO
PkYN OKOkOKANALIKOWY
Ryc. 258. Wchłanianie jonów przez nabłonek części bliższych kanalików nerkowych: P.D. —
różnica ładunku elektrycznego pomiędzy wnętrzem komórki nabłonka części bliższej
kanalika nerkowego, światłem kanalika i płynem ókołokanalikowym. Linia ciągła oznacza
transport aktywny, linia przerywana — bierną dyfuzję, zaś strzałka (NaCl + H2O) oznacza
filtrację z bocznej przestrzeni międzykomórkowej przez błonę podstawną do płynu okołokanalikowego. (Zmodyfikowano i reprodukowano za opłaconym zezwoleniem z R. F. Pitts
Physiology of the Kidney and Body Fluids, III wyd. Year Book Medical Publishers Inc.,
Chicago 1980.)
• biernego wydzielania (passive secretion), czyli dyfuzji, zgodnej z gradien
tem stężenia;
• aktywnego wydzielania o bezwzględnie ograniczonej najwyższej pojem
ności wydzielniczej (transport maximum limited mechanism);
• aktywnego wydzielania, którego pojemność zależna jest od gradientu
stężenia i czynnika czasu (grandient-time limitation).
Czynność nerek
485
Bierne wydzielanie na zasadzie dyfuzji dotyczy wydzielania słabych
zasad, np. soli amonowych, i słabych kwasów, np. kwasu salicylowego.
W przypadku, jeżeli mocz przepływający przez kanaliki osiąga pH 5
(SI: 10 umol H+/L), wówczas słabe zasady dyfundują z przestrzeni
okołokanalikowych przez nabłonek do światła kanalika. Alkalizacja
moczu przepływającego przez kanaliki do pH 8,0 (SI: 10 nmol H +/L)
powoduje dyfuzję słabych kwasów z przestrzeni okołokanalikowych do
światła kanalików.
---- Dyfu2Ja
Transport akiywny
Ryc. 259. Wchłanianie i wydzielanie jonów przez komórki nabłonka części dalszych
kanalików nerkowych i kanalików nerkowych zbiorczych. P.D. — różnica ładunku
elektrycznego pomiędzy wnętrzem komórki nabłonka części dalszej kanalika nerkowego,
światłem kanalika i płynem okołokanałikowym. Linia ciągła oznacza transport aktywny, linia
przerywana — dyfuzję. (Zmodyfikowano i reprodukowano za opłaconym zezwoleniem
z R. F. Pitts Physiology of the Kidney and Body Fluids, III wyd. Year Book Medical
Publishers Inc., Chicago 1980.)
486
Pfyny ustrojowe i czynność nerek
Kanalik
zbiorczy
Część wewnętrzna
rdzenia
15
Pętla nefronu
Ryc. 260. Wchłanianie wody w częściach kanalików nerkowych bliższych, w kanalikach
nerkowych dalszych i w kanalikach nerkowych zbiorczych. 100 L ultrafiltratu przefiltrowanego w kłębuszkach nerkowych ulega zmniejszeniu do 0,5 L moczu odpływającego do
miedniczki nerkowej.
Na zasadzie mechanizmu aktywnego wydzielania o bezwzględnie
ograniczonej najwyższej pojemności wydzielniczej wydzielane są przez
kanaliki związki egzogenne, takie jak kwas para-aminohipurowy, penicylina, sulfonamidy, preparaty Diodrast i Uroselectan, oraz endogenne, do
których należą: kreatynina i hormony steroidowe w postaci glukuronianów i siarczanów.
Czynność nerek
487
Drugi mechanizm aktywnego wydzielania o pojemności wydzielniczej
ograniczonej przez gradient stężenia i czynnik czasu odnosi się do
wydzielania jonów potasowych i wodorowych przez części dalsze kanalików
nerkowych i kanaliki nerkowe zbiorcze. Jony K+ i H+ są wydzielane do
światła kanalików nerkowych, a jednocześnie jony Na + zostają wchłaniane do komórek nabłonka.
Wchłanianie wody
Przesącz kłębuszkowy przepływający ze światła torebki kłębuszka do
kanalika krętego bliższego jest izoosmotyczny w stosunku do osocza i jego
ciśnienie osmotyczne jest równe 300 mOsm/L H 2O.
W części bliższej kanalika nerkowego wiele związków, przede wszystkim
jony sodowe, jest aktywnie wchłanianych przez nabłonek i woda biernie
podąża wraz z nimi do przestrzeni okołokanalikowej. W ramieniu
zstępującym pętli nefronu (Henlego) ciśnienie osmotyczne wzrasta, osiągając w miejscu jej zagięcia 1200 mOsm.
Wzrost ciśnienia osmotycznego jest spowodowany wzrostem ciśnienia
osmotycznego w przestrzeni okołokanalikowej w okolicy szczytów piramid nerkowych (pyramides renales).
Nabłonek części wstępującej pętli nefronu jest nieprzenikliwy dla wody.
W tym odcinku nefronu są aktywnie wchłaniane jony sodowe, które po
przejściu przez komórki nabłonka części wstępującej pętli nefronu (Henlego) gromadzą się w przestrzeni okołokanalikowej, wytwarzając w niej
wysokie ciśnienie osmotyczne.
W dalszych odcinkach kanalików nerkowych, jeżeli nie występuje
wazopresyna, nabłonek jest również nieprzenikliwy dla wody i ciśnienie
osmotyczne moczu obniża się do 100 mOsm.
W kanalikach nerkowych zbiorczych w miarę zbliżania się do ich ujścia
do miedniczki nerkowej ciśnienie osmotyczne moczu ponownie wzrasta do
1200 mOsm.
Zagęszczanie się moczu u dołu pętli nefronu i w kanalikach nerkowych
zbiorczych dzięki nagromadzonym jonom sodowym w przestrzeni okołokanalikowej przy szczytach piramid nosi nazwę przeciwprądowych wzmacniaczy (countercurrent multipliers).
Naczynia krwionośne włosowate sieci wtórnej, otaczające pętlę nefronu w postaci naczyń prostych (vasa recta), zachowują się tak, jak
przeciwprądowy wymiennik (countercurrent exchanger). Jony sodowe
wchłonięte do krwi w miarę jej przepływu przez naczynia proste w kie-
488
Płyny ustrojowe i czynność nerek
Kora
Część
zewnętrzna
rdzenia
Częsc
wewnętrzna^
rdzenia
Naczynie krwionośne
włosowate nroste
/i
PCf
Kanalik
zbiorczy
włosowat
e proste
Ryc. 261. Schemat nefronu i kanalika nerkowego zbiorczego oraz naczyń krwionośnych
włosowatych pierwotnej (kłębuszkowej) i wtórnej sieci (naczynia proste). Jony sodowe,
mocznik i woda przechodzą przez nabłonek części bliższej kanalików nerkowych i kanalika
zbiorczego do przestrzeni okołokanalikowej, a stąd do naczyń włosowatych prostych wtórnej
sieci naczyń krwionośnych.
runku do szczytu piramid wychodzą następnie z krwi do przestrzeni
okołokanalikowych w miejscu zawracania naczyń w kierunku kory nerki.
Naczynia proste, jako wymiennik przeciwprądowy, odprowadzają wodę
i inne związki z przestrzeni okołokanalikowych, jednocześnie pozostawiając w nich duże stężenie jonów sodowych.
Czynność nerek
Naczynie krwionośne
włosowate proste
p„ f/a
489
ne f ronu
Ryc. 262. Schemat nefronu i kanalika nerkowego zbiorczego oraz naczyń krwionośnych
włosowatych pierwotnej (kłębuszkowej) i wtórnej sieci (naczynia proste). Ciśnienie osmotyczne podane w miliosmolach, zaznaczono w części bliższej i w części dalszej kanalika nerkowego, w kanaliku zbiorczym oraz w płynie okołokanalikowym i w naczyniach włosowatych
prostych. Pętla nefronu (Henlego) działa jak wzmacniacz przeciwprądowy, naczynia proste
zaś jak wymiennik przeciwprądowy.
Czynniki wpływające na zagęszczenie moczu
Wazopresyna krążąca we krwi aktywuje enzym cyklazę adenylanową
w komórkach nabłonka części dalszej kanalików nerkowych. Pod wpływem tworzącego się w tych komórkach cyklicznego AMP woda zostaje
490
Płyny ustrojowe i czynność nerek
również resorbowana w części dalszej kanalików nerkowych. W następstwie postępującej resorpcji wody w kanalikach nerkowych zbiorczych
tylko 1 % wody przefiltrowanej w kłębuszkach nerkowych podlega ostatecznie wydaleniu z moczem.
Po wypiciu dużych ilości płynów hipotonicznych ciśnienie osmotyczne
osocza obniża się, uwalnianie wazopresyny z części nerwowej przysadki
zostaje zahamowane i następuje diureza wodna, charakteryzująca się
wydalaniem dużych ilości hipotonicznego moczu.
Wprowadzenie do organizmu związków nie wchłaniających się w kanalikach nerkowych powoduje, że więcej wody jest zatrzymywane w ich
świetle. W wyniku tego dochodzi do wydalania zwiększonej objętości
moczu, co nosi nazwę diurezy osmotycznej.
I Regulacja tworzenia się moczu
Tworzenie się moczu w nerkach zależy od optymalnego średniego
ciśnienia tętniczego w naczyniach kłębuszków. Czynniki obniżające znacznie średnie ciśnienie w zbiorniku tętniczym dużym lub kurczące tętnice
nerkowe, jak np. impulsacja biegnąca przez nerwy współczulne, zmniejszają filtrację kłębuszkową. Wzrost do pewnych granic ciśnienia w zbiorniku tętniczym dużym nie wpływa na ciśnienie w kłębuszkach dzięki
samoregulacji przepływu krwi przez nerki.
Na filtrację kłębuszkową wpływają pośrednio liczne hormony krążące we
krwi, zmieniając średnie ciśnienie w zbiorniku tętniczym dużym. Natomiast
wazopresyna (AVP), mineralokortykoidy, hormon gruczołów przytarczycznych (PTH) i przedsionkowy peptyd natriuretyczny (ANP — atrial natriuretic peptide) bezpośrednio wpływają na czynność kanalików nerkowych.
Wazopresyna aktywuje cyklazę adenylanową w nabłonku części dalszych kanalików nerkowych. Pod wpływem tworzącego się cyklicznego
AMP nabłonek staje się bardziej przenikliwy dla wody i w kanalikach
zwiększa się wchłanianie wody.
Mineralokortykoidy, a szczególnie aldosteron, zwiększają wchłanianie
jonów sodowych i wydzielanie jonów potasowych w częściach dalszych
kanalików nerkowych. Hormon gruczołów przytarczycznych (PTH) zwiększa wydalanie fosforanów z moczem dzięki zmniejszeniu ich wchłaniania
w kanalikach.
Prekursor przedsionkowych peptydów natriuretycznych jest polipeptydem o 151 aminokwasach wytwarzanym przez komórki mięśniowe
Czynność nerek
Tabela 41
Składniki moczu wydalane z organizmu w ciągu doby
stosowania diety
podczas
mieszanej Woda
1200 mL/d
Składniki nieorganiczne:
Chlorki
Sód
282
173
mmol/d
mmol/d
Fosfor
Potas
71
51
mmol/d
mmol/d
Siarka (jako SO3)
25
mmol/d
5
mmol/d
6
mmol/d
Wapń
Magnez
Jod
0,4—2, 0
urnol/d
210—500
12
mmol/d
mmol/d
41
4
mmol/d
mmol/d
0,01—2,
10 mmol/d
Składniki organiczne:
Mocznik
Kreatynina
Amoniak
Kwas moczowy
Kreatyna
Ryc. 263. Cząsteczka przedsionkowego peptydu
natriuretycznego (ANP).
491
492
Płyny ustrojowe i czynność nerek
przedsionków serca. Aktywne fragmenty prekursora, tworzące pierścień
z jedenastu aminokwasów i dwóch krótkich łańcuchów peptydowych
(zbudowane z 22, 23, 24 lub 28 aminokwasów), uwalniane są do krwi po
rozciągnięciu ścian przedsionków serca, przede wszystkim pod wpływem
zwiększonej całkowitej objętości krwi krążącej (TBV). Przedsionkowe
peptydy natriuretyczne (ANP) zwiększają wydalanie przez nerki jonów
sodowych i wody i jednocześnie obniżają ciśnienie tętnicze krwi. Ich
działanie rozkurczające mięśnie gładkie naczyń krwionośnych jest antagonistyczne w stosunku do wazopresyny i układu hormonów renina-angiotensyna-aldosteron. Hamują również wydzielanie tych hormonów
do krwi.
I Czynności wewnątrzwydzielnicze
Obniżenie się ciśnienia tętniczego krwi lub osłabienie prężności tlenu
w nerkach powoduje wydzielanie przez nerkę związków podwyższających
ciśnienie tętnicze lub zwiększających erytropoezę w szpiku kostnym.
Renina
Mięśnie gładkie tętniczki doprowadzającej w miejscu przylegania do
plamki gęstej (macula densa) kanalika krętego dalszego tworzą aparat
przykłębuszkowy {apparatus iuxtaglomerularis). W czasie niedokrwienia
nerek aparat przykłębuszkowy wydziela do krwi reninę, która jest
enzymem proteolitycznym działającym na alpha 2-globulinę osocza (angiotensynogen), odczepiając od niej nieaktywny dekapeptyd — angiotensynę I. Pod wpływem enzymu konwertującego nieaktywna angiotensyna I
ulega przekształceniu na aktywny oktapeptyd — angiotensynę II.
Zasadniczymi czynnikami wywołującymi wydzielanie reniny przez nerki są:
• obniżenie się ciśnienia w zbiorniku tętniczym dużym;
• zmniejszenie się przepływu nerkowego krwi wskutek skurczu błony
mięśniowej tętnic nerkowych;
• zmniejszenie się stężenia NaCl w moczu pierwotnym przepływającym
przez kanalik kręty dalszy koło plamki gęstej;
• napływające z krwią tętniczą eikozanoidy takie, jak prostaglandyny
E2 (PGE ) i prostacyklina (PGI 2 ) oraz kininy.
Czynność nerek
493
Ryc. 264. Powstawanie angiotensyny II.
Erytropoetyna
Miejscem wytwarzania erytropoetyny, która jest jednym z czynników wzrostowych, są komórki śródbłonka naczyń włosowatych otaczających kanaliki nerkowe w korze nerek. Erytropoetyna jest glikoproteiną o masie cząsteczkowej około 19 kDa i okresie połowicznego rozpadu we krwi (T 1 /^ około 5 godzin. Pobudza erytropoezę
w szpiku kostnym. W wyniku utraty krwi lub przewlekłej hipoksji,
spowodowanej na przykład przebywaniem na dużej wysokości, zwiększa
się wydzielanie erytropoetyny. Również aminy katecholowe, działając
poprzez beta-receptory adrenergiczne, zwiększają wydzielanie erytropoetyny.
Metabolizm cholekalcyferolu
W skórze pod wpływem światła ultrafioletowego prowitamina D,
czyli 7-dehydrocholesterol, zamieniana jest na witaminę D3 — cholekalcyferol. Witamina D 3 wchłonięta do krwi zostaje przekształcona w wątrobie na 25-hydroksycholekalcyferol (HD 3 ) i następnie
w nerkach na dwa związki: 1,25-dihydroksycholekalcyferol (1,25-DHD3)
i 24,25-dihydroksycholekalcyferol (24,25-DHD 3 ). Pod wpływem
1,25-DHD , działającego na nabłonek jelita i na
się stężenie Ca 2+ i tkankę kostną, zwiększa
w osoczu krwi. Zwiększenie stężenia tych
494
Płyny ustrojowe i czynność nerek
jonów w osoczu krwi powoduje, że w nerkach tworzy się mało aktywny
24,25-DHD3 kosztem zmniejszonego wytwarzania aktywnego związku
1,25-DHD3.
Metabolizując pochodne cholekalcyferolu, nerki uczestniczą w regulacji
przemiany wapniowo-fosforanowej w organizmie.
I Wydalanie moczu
Mocz wydostający się z kanalików nerkowych zbiorczych gromadzi się
w miedniczce nerkowej (pehis renalis). Na skutek ruchów perystaltycznych
moczowodu (ureter) porcje moczu przemieszczane są z miedniczki nerkowej do pęcherza moczowego (vesica urinarid).
Wypełnianie się pęcherza moczowego powoduje rozciąganie jego
ścian i podrażnienie receptorów. Na drodze odruchowej poprzez ośrodek oddawania moczu w części krzyżowej rdzenia kręgowego występuje
jednoczesny skurcz mięśnia gładkiego w ścianie pęcherza moczowego,
zwanego mięśniem wypierającym {musculus detrusor), rozkurcz mięśnia gładkiego zwieracza wewnętrznego cewki moczowej {musculus sphincter urethrae internus) i rozkurcz mięśnia poprzecznie prążkowanego
stanowiącego zwieracz zewnętrzny cewki moczowej {musculus sphincter
urethrae externus). Na skutek zwiększonego ciśnienia w pęcherzu moczowym i rozkurczu zwieraczy mocz zostaje wydalony przez cewkę moczową
na zewnątrz.
Odruchowy mechanizm oddawania moczu na skutek pobudzenia
ośrodka w części krzyżowej rdzenia kręgowego występuje u niemowląt
i u ludzi dorosłych po przerwaniu ciągłości rdzenia kręgowego. U dzieci
w obrębie pnia mózgowia wykształcają się ośrodki nadrzędne, kontrolujące ośrodek oddawania moczu.
U ludzi dorosłych wydalanie moczu z pęcherza moczowego jest
czynnością dowolną. Wypełnienie pęcherza moczowego 250—300 mL
moczu pobudza receptory w ścianie pęcherza moczowego, wywołuje
impulsację aferentną przewodzoną przez drogi rdzeniowo-wzgórzowe
i następnie ze wzgórza do pól czuciowych w korze mózgu. Czucie to nasila
się po dalszym zwiększeniu objętości moczu w pęcherzu do 400—500 mL.
Eferentna impulsacja biegnąca z kory płatów czołowych do ośrodków
mikcji w moście powoduje:
• pobudzenie neuronów prz\ środkowego ośrodka mikcji w moście (medial
pontine micturition center) i zależnych od tego ośrodka neuronów
Wydalanie moczu
495
Ryc. 265. Ośrodki nerwowe kontrolujące mikcję. Impulsy z receptorów wrażliwych na
rozciąganie w ścianie pęcherza moczowego są przewodzone przez włókna aferentne w splocie
krzyżowym (linia przerywana) do rdzenia kręgowego, części krzyżowej (S2, S3, S4) i do
ośrodka mikcji w części przysrodkowej mostu. Z ośrodka hamującego mikcję w części bocznej
mostu biegną impulsy eferentne do neuronów przedzwojowych w części lędźwiowej rdzenia
kręgowego, przez zwój krezkowy dolny i nerw podbrzuszny do mięśnia wypierającego
pęcherza moczowego i do mięśnia zwieracza wewnętrznego cewki moczowej oraz do
neuronów ruchowych i przez nerw sromowy do mięśnia zwieracza zewnętrznego cewki
moczowej.
496
Płyny ustrojowe i czynność nerek
Początek
przepływu
cewkowego
Maksymalny
przepływ
cewkowy
Koniec
przepływu
cewkowego
I
Ciśnienie w
jamie
brzusznej
Ciśnienie w
pęcherzu
moczowym
Przepływ
cewkowy
moczu w
mL/s
Ryc. 266. Ciśnienie w
jamie brzusznej, ciśnienie
w pęcherzu moczowym i
przepływ cewkowy moczu. 1 — czas uzyskania maksymalnego przepływu cewkowego
moczu, 2 — czas trwania mikcji.
przywspółczulnych, w rogach bocznych w części krzyżowej rdzenia
kręgowego, unerwiających mięsień wypierający w ścianie pęcherza
moczowego;
hamowanie neuronów bocznego ośrodka mikcji w moście (lateral pontine
micturition center) i kontrolowanych przez ten ośrodek neuronów
ruchowych w części krzyżowej rdzenia kręgowego, unerwiających
mięsień zwieracz zewnętrzny cewki moczowej. Skurcz mięśnia wypierającego i rozkurcz mięśnia zwieracza zewnętrznego cewki moczowej
rozpoczyna opróżnianie się pęcherza moczowego.
Mi kej a
Wypływ moczu z pęcherza moczowego przez cewkę, w czasie dowolnej
mikcji, poprzedza wzrost ciśnienia w jamie brzusznej spowodowany
Wydalanie moczu
497
odruchowym skurczem mięśni w ścianach jamy brzusznej i wzrost ciśnienia śródpęcherzowego. Dynamika procesu mikcji jest badana nieinwazyjnie metodą pomiaru przepływu cewkowego. Polega ona na automatycznym
pomiarze i wykreśleniu krzywej objętości wydalonego przez cewkę moczu
w każdej sekundzie mikcji.
Zapisana krzywa przepływu cewkowego wykazuje maksymalną i średnią szybkość przepływu i czas przepływu moczu przez cewkę. Przy 400 mL
wydalanej objętości moczu u kobiet maksymalna szybkość przepływu
wynosi od 20 do 60 mL/s, średnia szybkość przepływu zaś 10—30 mL/s.
U mężczyzn przy tej samej objętości wydalanego moczu maksymalna
szybkość przepływu jest od 18 do 46 mL/s, średnia zaś od 11 do 28 mL/s.
Czas potrzebny do osiągnięcia maksymalnej szybkości przepływu zarówno u kobiet, jak i u mężczyzn wynosi l / 3 czasu mikcji.
Rozród
I Determinacja płci
Połączenie się dwóch odmiennych komórek płciowych, czyli gamet
(gametes), powstanie zygoty i rozwój nowego ustroju potomnego są
podstawą rozrodu (reproduction). Komórka płciowa męska, czyli plemnik (spermium), wytwarzana jest w jądrze (testis), a komórka płciowa
żeńska, czyli jajo (ovum), powstaje w jajniku (ovarium).
Zarówno komórki płciowe męskie, jak i żeńskie w czasie dojrzewania
wykazują podział redukcyjny (meiotic division), w czasie którego liczba
chromosomów zmniejsza się o połowę. W organizmie mężczyzny i kobiety
w jądrze komórkowym komórek somatycznych jest 46 chromosomów.
U mężczyzny są to 22 pary autosomów i dwa chromosomy p łciowe:
X i Y. U kobiety występują również 22 pary autosomów i dwa chromosomy płciowe: X i X.
W czasie podziału redukcyjnego w cewkach nasiennych krętych jąder
powstają komórki płciowe męskie zawierające 22 chromosomy (autosomy) i jeden chromosom płciowy X lub Y. W jajnikach dojrzałe komórki
płciowe żeńskie mają 22 chromosomy i 1 chromosom płciowy X. Od
informacji genetycznej zawartej w chromosomach płciowych zależy płeć
osobnika. Po połączeniu się dwóch dojrzałych komórek płciowych, czyli po
zapłodnieniu (fertilization), i powstaniu zygoty determinuje się płeć genetyczna (genetic sex) przyszłego organizmu. Jeżeli zygota zawiera chromosomy XY, będzie to genetycznie organizm męski, jeśli zaś zygota ma
2 chromosomy XX — genetycznie organizm żeński.
W 9 tygodniu życia płodowego pod wpływem hormonów wydzielanych
przez gruczoły płciowe — testosteronu u płodów męskich i estrogenów
u płodów żeńskich — tworzy się płeć gonadalna (gonadal sex). Garnitur
Rozród u pici męskiej
499
chromosomalny i hormony płciowe warunkują płeć somatyczną (somatic
sex), na którą składa się rozwój narządów płciowych wewnętrznych
i zewnętrznych oraz budowa ciała i zachowanie typowe dla płci męskiej
lub żeńskiej.
Gruczoły płciowe nie wykazują większej aktywności w organizmach
wzrastających aż do czasu osiągnięcia dojrzałości płciowej, która występuje u chłopców średnio w 14 roku życia i u dziewcząt w 12 roku życia.
W okresie pierwszych kilkunastu lat życia wytwarzanie i wydzielanie do
przysadkowych naczyń wrotnych hormonu uwalniającego gonadotropiny
z przysadki (GnRH) jest hamowane m.in. przez melatoninę powstającą
w szyszynce. Osiągnięcie dojrzałości płciowej jest uwarunkowane zwiększeniem wydzielania gonadotropin przez część gruczołową przysadki.
! Rozród u płci męskiej
I Spermatogeneza
Po osiągnięciu dojrzałości płciowej w cewkach nasiennych krętych
(tubuli seminiferi contorti) jąder zachodzi spermatogeneza. Komórki
płciowe w cewkach nasiennych tworzą kilka warstw. Pomiędzy nimi
występują komórki podporowe. Komórki płciowe dojrzewając zbliżają się
do światła cewek nasiennych. Najdalej od światła w cewkach nasiennych są
rozmieszczone spermatogonie. następnie występują spermatocyty I i II
rzędu, spermatydy i w samym świetle cewek znajdują się dojrzałe komórki
płciowe męskie, czyli plemniki. Dojrzewanie męskiej komórki płciowej
trwa około 74 dni.
Plemniki są transportowane wraz z innymi składnikami nasienia przez
cewki nasienne proste i przewodziki odprowadzające jądra do najądrza
(epididymis), w którego przewodzie są magazynowane.
Hormon folikulotropowy (FSH) pobudza czynność komórek podporowych w cewkach nasiennych. Spermatogeneza jest kontrolowana
przez hormon folikulotropowy, hormon luteinizujący i testosteron wytwarzany przez komórki śródmiąższowe jądra.
500
Rozród
Ryc. 267. Spermatogeneza w cewkach nasiennych krętych jąder. Dojrzewanie męskich
komórek płciowych przebiega kolejno od spermatogonii (1), przez podział mitotyczny (2),
spermatocyty I rzędu (3), podział mejotyczny I (4), spermatocyty II rzędu przed podziałem
mejotycznym II (5), do spermatyd (6) i plemników (7). 8 — komórka podporowa (Sertolego)
z jądrem komórkowym i jąderkiem, której cytoplazma (9) otacza męskie komórki płciowe,
10 — błona podstawna cewek nasiennych krętych jądra.
Wzwód prącia i wytrysk nasienia
Wzwód prącia (erection) zachodzi w następstwie pobudzenia neuronów
przywspółczulnych w rogach bocznych segmentów krzyżowych (S2—S4)
rdzenia kręgowego. Pod wpływem impulsacji przywspółczulnej rozkurczają się tętniczki doprowadzające krew do ciał jamistych prącia {corpus
cavernosum penis) i ciała gąbczastego prącia {corpus spongiosum penis)
i następuje wzwód prącia.
Unerwienie współczulne ma działanie przeciwne, jeśli chodzi o wypełnienie się ciał jamistych krwią, natomiast wywołuje skurcz najądrzy
{epididymis), nasieniowodów {ductus deferens), pęcherzyków nasiennych
{vesicula seminalis) i gruczołu krokowego {prostata). Dzięki tym skurczom
nasienie wypełniające przewód najądrza i nasieniowód jest wydalane do
Rozród u pici żeńskiej
Gruczoły błony
podśluzowej
Zrąb
501
Gruczoły^ błony
śluzowej
Przewody
Cewka moczowa
Ryc. 268. Schematyczna budowa gruczołu krokowego na przekroju poprzecznym. (Reprodukowano za zezwoleniem z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie
PZWL, Warszawa 1997.)
cewki moczowej wraz z wydzieliną pęcherzyków nasiennych i gruczołu
krokowego. Skurcz zwieracza wewnętrznego cewki moczowej zapobiega
przepływowi nasienia do pęcherza moczowego.
Po emisji nasienia do cewki moczowej następuje wytrysk nasienia, czyli
ejakulacja. Pobudzenie receptorów w cewce moczowej i w gruczole
krokowym powoduje na drodze odruchowej skurcz toniczno-kloniczny
mięśni opuszkowo-jamistego {musculus bulbocavernosus) i kulszowo-jamistego {musculus ischiocavernosus). Rytmiczne skurcze tych mięśni powodują wyrzucanie nasienia z cewki moczowej na zewnątrz. Ośrodki odruchowe w rdzeniu kręgowym związane ze wzwodem prącia i wytryskiem
nasienia są kontrolowane przez ośrodki w mózgowiu.
| Rozród u płci żeńskiej
I Cykl płciowy żeński
Cykle płciowe (sexual cycles) zaczynają występować u dziewcząt
w okresie pokwitania (puberty) w wieku od 11 do 14 lat. Przekwitanie
(menopause) objawia się wydłużeniem się i zanikaniem cykli płciowych, co
502
Rozród
zachodzi u kobiet w wieku od 45 do 55 roku życia. Zasadniczym objawem
wskazującym na występowanie cykli płciowych są krwawienia miesiączkowe. W czasie krwawienia dochodzi do utraty łącznie około 70 mL krwi.
Dzień, w którym pojawia się krwawienie, rozpoczyna cykl miesiączkowy
trwający przeciętnie 28 dni.
W cyklu płciowym żeńskim wyróżnia się:
•
•
•
•
krwawienie miesiączkowe (menstruation), które trwa przeciętnie 4 dni;
fazę folikularną (follicular phase), która trwa około 9 dni;
owulację (ovulation), która trwa 1 dzień;
fazę lutealną (luteal phase), która trwa 14 dni.
Poszczególne dni cyklu miesiączkowego charakteryzują się określoną
czynnością wewnątrzwydzielniczą podwzgórza, przysadki i jajników oraz
towarzyszącymi im zmianami morfologicznymi i czynnościowymi w obrębie jajników, macicy, pochwy i w sutkach.
Hormonalne sprzężenie zwrotne
Regularne występowanie cykli płciowych u dojrzałych płciowo kobiet
zależy od fizjologicznego mechanizmu kontrolowanego przez żeński
ośrodek rozrodczy w podwzgórzu. Ośrodek ten pozostaje pod wpływem
różnorodnych oddziaływań. Przed osiągnięciem dojrzałości płciowej
szyszynka i wydzielana przez nią melatonina hamują czynność ośrodka
rozrodczego w podwzgórzu, opóźniają pokwitanie i wystąpienie
pierwszych cykli miesiączkowych. Układ limbiczny, a zwłaszcza należące
do niego ciało migdałowate, działają przeciwnie przyspieszając pokwitanie.
Regularna owulacja i występowanie krwawienia miesiączkowego u dojrzałych płciowo kobiet zależy od kolejnego oddziaływania wielu hormonów na ośrodek rozrodczy w podwzgórzu. Ośrodek ten stanowi
zasadnicze ogniwo w humoralnym sprzężeniu zwrotnym dla pętli zarówno
zewnętrznego, jak i wewnętrznego sprzężenia zwrotnego.
Ogniwami pętli zewnętrznego sprzężenia zwrotnego (external feedback
loop) są podwzgórze, płat gruczołowy przysadki i jajniki. W sprzężeniu
tym hormony wytwarzane w jajniku oddziałują na podwgórze, hamując
wydzielanie hormonu uwalniającego gonadotropiny z płata gruczołowego
przysadki (GnRH). Prawidłowe, czyli pulsacyjne, wydzielanie GnRH
przez podwzgórze do przysadkowych naczyń wrotnych w fazie folikularnej
cyklu miesiączkowego powoduje również prawidłowe, czyli pulsacyjne,
Rozród u pici żeńskiej
503
wydzielanie przez płat gruczołowy przysadki hormonów luteinizującego
(LH) i folikulotropowego (FSH) oraz przez jajnik — estrogenów. W ostatnich dniach fazy folikularnej zawartość estrogenów we krwi zmniejsza
się. Jest to prawdopodobnie spowodowane zwrotnym, hamującym działaniem estrogenów na komórki płata gruczołowego przysadki wytwarzające hormony gonadotropowe. Również zwrotnie hamuje wydzielanie FSH
czynnik zwany inhibiną wytwarzany przez komórki warstwy ziarnistej
pęcherzyka jajnikowego dojrzałego.
W wyniku zmniejszenia się zawartości estrogenów we krwi komórki
podwzgórza przestają być zwrotnie hamowane przez estrogeny i zaczynają
uwalniać od przysadkowych naczyń wrotnych większe ilości GnRH. W
następstwie zwiększonego uwalniania przez podwzgórze tego neurohormonu przysadka wydziela do krwi w ciągu kilku godzin większą ilość
hormonu luteinizującego (LH), co powoduje owulację.
Wydzielanie pulsacyjne GnRH przez podwzgórze i gonadotropin przez
przedni płat przysadki występuje w odstępach 90—120 minut. Częstotliwość występowania i amplituda stężenia we krwi hormonów gonadotropowych są modulowane przez kilka czynników. Największa amplituda
wydzielanych pulsacyjnie gonadotropin występuje we krwi w pierwszych
dniach fazy folikularnej w porze dziennej.
Systematyczne wprowadzanie do organizmu egzogennych estrogenów
w fazie folikularnej cyklu miesiączkowego hamuje czynność podwzgórza.
Nie dochodzi do znacznego zwiększenia wydzielania LH i pęcherzyki
jajnikowe dojrzałe nie pękają — nie ma więc owulacji.
Ryc. 269. Wydzielanie hormonów w fazie owulacyjnej i lutealnej cyklu miesiączkowego.
504
Rozród
Wprowadzane egzogenne estrogeny hamują wydzielanie GnRH przez
podwzgórze i powodują występowanie bezowulacyjnych cykli miesiączkowych (unovulatory menstrual cycles).
W pętli wewnętrznego sprzężenia zwrotnego (internal feedback loop)
uczestniczą: podwzgórze, podwzgórzowy hormon uwalniający gonadotropiny — GnRH, płat gruczołowy przysadki i wydzialane przez przysadkę gonadotropiny — FSH i LH. Znaczenie wewnętrznego sprzężenia
zwrotnego w regulacji żeńskiego cyklu płciowego nie zostało dotychczas
dostatecznie poznane.
Cykl jajnikowy i maciczny
W czasie krwawienia miesiączkowego dochodzi do złuszczania się
warstw powierzchniowych błony śluzowej macicy i ich wydalania na
zewnątrz wraz z krwią.
W fazie folikularnej zwanej również proliferacyjną, następuje odnowa
błony śluzowej macicy. W końcu tej fazy, trwającej przeciętnie około 9 dni,
Ryc. 270. Pęcherzyk jajnikowy dojrzewający w końcu fazy folikularnej cyklu miesiączkowego.
Komórka jajowa (1) otoczona osłonką przejrzystą (2), wieńcem promienistym (3) i war stwą ziarnistą (4) tworzy wzgórek jajonośny wewnątrz jamy pęcherzyka (6) wypełnionej
płynem. Osłonka pęcherzyka (5) składa się z warstwy zewnętrznej
(łącznotkankowej) i z warstwy wewnętrznej zawierającej komórki gruczołowe
wydzielające estrogeny. (Reprodukowano za zezwoleniem Michała Karaska.)
Rozród u płci żeńskiej
505
błona śluzowa macicy osiąga taką grubość, jaką miała bezpośrednio przed
złuszczaniem się. Odnowa dotyczy również gruczołów błony śluzowej
macicy, które w fazie folikularnej mają przebieg prosty: od powierzchni
błony aż do mięśnia macicznego. Na początku tej fazy kilka pęcherzyków
jajnikowych pierwotnych {folliculi ovarici primarii) zaczyna w obu jajnikach przekształcać się w pęcherzyki jajnikowe wzrastające (folliculi ovarici
crescendi). Następnie zazwyczaj jeden, a czasami dwa pęcherzyki dojrzewają, osiągając średnicę co najmniej 10 mm. Pęcherzyk jajnikowy
dojrzały (folliculus ovaricus maturus) zajmuje całą szerokość kory jajnika i uwypukla się na jego powierzchni. Pozostałe pęcherzyki zmniejszają
swoje rozmiary i uwsteczniają się. Proces ten nosi nazwę ątręzjf.
W cyklu 28-dniowym zazwyczaj czternastego dnia cyklu miesiączkowego następuje pękanie pęcherzyka jajnikowego dojrzałego pod wpływem hormonu luteinizującego (LH) i komórka jajowa, owocyt II rzędu,
zostaje wydalona przez jajnik. W czasie jednego cyklu miesiączkowego
stadium pęcherzyka jajnikowego dojrzałego osiąga w obu jajnikach jeden
lub dwa pęcherzyki jajnikowe wzrastające. Tym samym z obu jajników
zostaje wydalona jedna lub rzadziej dwie komórki jajowe. Po owulacji
zmniejsza się w mięśniu macicy liczba receptorów błonowych wiążących
oksytocynę i zmniejsza się jego wrażliwość na oksytocynę.
Komórka jajowa w postaci owocytu II rzędu trafia do bańki jajowodu.
W jajowodzie nie zapłodniona komórka jajowa pozostaje około 4 dni,
a następnie zostaje wydalona do jamy macicy, gdzie podlega cytolizie.
Po owulacji rozpoczyna się faza lutealna, zwana również fazą sekrecyjną. W końcu pierwszej połowy tej fazy błona śluzowa macicy osiąga swoją
maksymalną grubość około 6 mm. W tej fazie gruczoły błony śluzowej
macicy mają przebieg kręty i wydzielają duże ilości śluzu. Błona śluzowa
macicy jest przygotowana do zagnieżdżenia zapłodnionej komórki jajowej.
Jama pęcherzyka jajnikowego dojrzałego po wydaleniu komórki jajowej początkowo jest wypełniona krwią. Następnie zapełniają ją komórki
luteinowe {luteocyti) i pęcherzyk zamienia się w ciałko żółte miesiączkowe
(corpus luteum menstruationis), które wytwarza żeńskie hormony płciowe:
/Sstrogeny i progesteron oraz oksytocyncJW końcu fazy lutealnej, jeżeli
nie nastąpiło zapłodnienie, ciałko żółte miesiączkowe przestaje wydzielać
progesteron, natomiast w dalszym ciągu wydziela oksytocynę. Zwiększa
się jednocześnie liczba receptorów wiążących oksytocynę i mięsień macicy
staje się na nią bardziej wrażliwy. Pod jej wpływem tworzą się w macicy
substancje luteolityczne, głównie prostaglandyny PGF2a i ciałko żółte
miesiączkowe przekształca się w ciałko białawe miesiączkowe {corpus
albicans menstruationis). Po upływie 14 dni od dnia owulacji pojawia się
krwawienie miesiączkowe i rozpoczyna się nowy cykl płciowy.
506
0
Rozród
2 4
6
8
10 12 14 16 18 20 22 24 26 28 128 dzień cyklu miesiącikowego
Temperatura ciała
Ryc.
271.
Procesy
zachodzące
w
podwzgórzu,
w przysadce,
w jajnikach i
w błonie śluzowej macicy
w
poszczególnych dniach cyklu miesiączkowego. W fazie lutealnej cyklu miesiączkowego
zwiększa się we krwi zawartość hormonu luteinizującego (LH) i następuje pękanie
pęcherzyka jajnikowego dojrzałego. W poszczególnych dniach cyklu zmienia się
temperatura ciała, zawartość we krwi hormonów gonadotropowych (FSH i LH), estrogenów i progesteronu oraz zachodzą zmiany w błonie śluzowej macicy. GnRH — hormon
uwalniający gonadotropiny.
Rozród u pici żeńskiej
507
Pod wpływem hormonów płciowych wydzielanych przez jajnik w różnych dniach cyklu miesiączkowego dochodzi do zmian w ilości i konsystencji śluzu wypełniającego szyjkę macicy. W fazie lutealnej powiększają się
sutki, w końcu tej fazy zaś, przed wystąpieniem krwawienia miesiączkowego, następuje zatrzymanie wody w organizmie.
W czasie cyklu miesiączkowego stwierdza się zmiany temperatury ciała
mierzonej w godzinach porannych. W fazie folikularnej temperatura ciała
mierzona w godzinach porannych nie przekracza 37°C. W dniu owulacji
temperatura ciała obniża się, a następnego dnia, już w fazie lutealnej, jest
podwyższona i przekracza 37°C. Do końca fazy lutealnej temperatura
w godzinach rannych jest stale podwyższona, a przed wystąpieniem
miesiączki obniża się.
I Ciąża
Zapłodnienie (fertilization), czyli połączenie się męskiej komórki płciowej z żeńską komórką płciową, następuje w jajowodzie. Zapłodniona
komórka jajowa przemieszcza się przez jajowód do jamy macicy w czasie
3—4 dni. W tym okresie podlega podziałom i przemianom.
Zapłodnione jajo zagnieżdża się (implantation) w błonę śluzową macicy około 7 dnia od dnia owulacji, czyli w połowie fazy lutealnej. [Prawdopodobnie implantowane jajo oddziałuje hamująco na tworzenie się
i uwalnianie z macicy czynnika luteolitycznego — prostaglandyn. Dzięki
temu ciałko żółte miesiączkowe nie zanika, lecz przeciwnie, powiększa
swoje wymiary i przekształca się w ciałko żółte ciążowej (corpus luteum
graviditatis). Wydzielanie progesteronu przez ciałko żółte ciążowe zwiększa
się kilkakrotnie w czasie pierwszych kilkunastu tygodni ciąży (pregnancy).
Największe wydzielanie progesteronu przez jajniki stwierdzono w 12
tygodniu ciąży, licząc czas trwania ciąży od dnia owulacji. Następnie
wydzielanie progesteronu przez jajniki zmniejsza się w miarę zbliżania się
porodu. Rozwijające się łożysko przejmuje funkcję wydzielniczą jajników,
wydzielając do krwi znaczne ilości progesteronu i estrogenów.
Czynność łożyska
Łożysko (placenta), tworzące się w pierwszych tygodniach ciąży, w 10
tygodniu ciąży, licząc od dnia owulacji, waży około 10 g, w 20 tygodniu —
około 170 g, w 30 tygodniu — około 430 g i największą swą masę osiąga
w 40 tygodniu ciąży — około 500—600 g.
508
Rozród
Zasadniczą funkcją łożyska jest wymiana gazów (O2 i CO2), produktów
energetycznych i budulcowych oraz ich metabolitów pomiędzy krwią matki
a krwią płodu. Przez łożysko w ciągu jednej sekundy przepływa 8,3 raL
krwi matki i 6,6 mL krwi płodu. Wymiana pomiędzy krwią matki a krwią
płodu zachodzi dzięki dyfuzji, aktywnemu transportowi i pinocytozie.
W łożysku dyfundują zgodnie z gradientem koncentracji lub prężności:
tlen, dwutlenek węgla i większość soli mineralnych. Na zasadzie aktywnego transportu, wbrew gradientowi koncentracji, przechodzą przez
łożysko: glukoza, aminokwasy oraz jony wapniowe i żelazowe. W wyniku
pinocytozy w łożysku są transportowane z krwi matki do krwi płodu
nieznaczne ilości białek.
Drugą ważną funkcją łożyska jest wytwarzanie hormonów. Poczynając od 8 tygodnia ciąży, łożysko stopniowo przejmuje wytwarzanie
progesteronu, zastępując w ten sposób czynność ciałka żółtego ciążowego.
0 dobowym wytwarzaniu progesteronu pośrednio można wnioskować^
na podstawie oznaczania zawartości pregnanendiolu — metabolitu
progesteronu wydalanego z moczem. Zawartość pregnanendiolu w do
bowej porcji moczu zwiększa się od około 2 mg w 8 tygodniu ciąży do
około 40 mg w 36—40 tygodniu ciąży. Ilość dobowa pregnanendiolu
wydalana z moczem stanowi około 15% wytwarzanego progesteronu. Tym
samym w końcu ciąży łożysko wytwarza dziennie około 300 mg proges
teronu.
We krwi tylko około 10% progesteronu znajduje się w postaci wolnej
1 wykazującej aktywność fizjologiczną. Pozostałe ponad 90% proges
teronu jest związane z białkami osocza i nie wykazuje aktywności
fizjologicznej. Progesteron osłabia pobudliwość mięśnia macicy oraz
obkurcza ujście wewnętrzne szyjki macicy. Dzięki temu zapobiega wy
stąpieniu poronienia lub porodu przedwczesnego. Działa synergicznie
wraz z estrogenami na wzrost sutków w czasie ciąży.
W czasie ciąży również znacznie zwiększa się wydzielanie estrogenów,
które powstają głównie w łożysku. Spośród trzech estrogenów: estriolu.
estronu i estradiolu w dobowej porcji moczu najbardziej zwiększa się
zawartość estriolu, znacznie mniej zaś estronu i estriadolu. Zawartość
estriolu w dobowej porcji moczu i we krwi stale zwiększa się, poczynając
od początku ciąży aż do porodu. Również zwiększeniu ulega jego zawartość w wodach płodowych. W końcu ciąży koncentracja jego we krwi
jest około 1000 razy większa w porównaniu z koncentracją we krwi w czasie cyklu miesiączkowego.
Pod wpływem estrogenów następują podziały komórek mięśnia macicznego i zwiększenie jego masy w czasie ciąży. Jednocześnie estrogeny
zwiększają pobudliwość mięśnia macicznego i tym samym mają działanie
Rozród u pici żeńskiej
509
antagonistyczne w stosunku do progesteronu, tak że mięsień ten zaczyna
kurczyć się pod wpływem krążącej we krwi oksytocyny.
Poza hormonami steroidowymi łożysko wytwarza również hormony
białkowe. Są to: ludzka gonadotropina łożyskowa (hCG) i ludzki łożyskowy laktogen (hPL) oraz w nieznacznych ilościach hormony stale
wytwarzane przez płat przedni przysadki.
Największe wydzielanie ludzkiej gonadotropiny łożyskowej (hCG) przypada na 8 tydzień ciąży i w następnych tygodniach zaczyna się zmniejszać.
Poczynając od 12 tygodnia ciąży aż do dnia porodu, zawartość hCG we
krwi matki stopniowo zmniejsza się w porównaniu z zawartością w 8 tygodniu ciąży. Pod wpływem hCG ciałko żółte ciążowe wzrasta i wydziela
progesteron. Zmniejszeniu się wydzielania hCG towarzyszy stopniowe
zanikanie ciałka żółtego ciążowego w jajnikach.
Działanie ludzkiego łożyskowego laktogenu (hPL) jest podobne do
działania hormonu wzrostu. Pod jego wpływem następuje wzrost sutków
w czasie ciąży.
I Poród
Po upływie 280 dni ciąży, licząc od pierwszego dnia ostatniej miesiączki,
następuje poród (parturition). Tylko 5—10% kobiet rodzi w tym czasie. Znaczny procent kobiet (66%) rodzi w czasie do 11 dni krótszym lub dłuższym.
Mechanizm rozpoczynający poród nie jest w pełni poznany. Przypuszcza
się, że podwzgórze płodu po osiągnięciu określonego stadium rozwoju
zaczyna wydzielać podwzgórzowe hormony uwalniane do przysadkowych naczyń wrotnych. Pod wpływem tych hormonów płat gruczołowy
przysadki zwiększa wydzielanie hormonów tropowych, wśród nich również hormonu kortykotropowego. Hormon ten pobudza korę gruczołu
nadnerczowego płodu do znacznego wydzielania kortyzolu, który — krążąc we krwi płodu — przechodzi do wód płodowych i działa na mięsień
maciczny antagonistycznie w stosunku do progesteronu.
Inny przypuszczalny mechanizm rozpoczynający poród wiąże się ze
zwiększeniem koncentracji estrogenów w wodach płodowych. Zwiększenie
koncentracji estrogenów, zwłaszcza estriolu, w wodach płodowych powoduje ich większe przenikanie do mięśnia macicznego, zwiększenie jego
pobudliwości, wystąpienie pierwszych skurczów i rozpoczęcie porodu.
W czasie porodu następuje wydalanie płodu wraz z błonami płodowymi i łożyskiem. W większości fizjologicznych porodów częścią przodującą
jest główka płodu ustawiona potylicowo. Poród fizjologiczny dzieli się na
510
Rozród
Ryc. 272. Początek I okresu porodu — rozwieranie się szyjki macicy.
trzy okresy. W pierwszym okresie rozpoczyna się czynność porodowa,
która kończy się pełnym rozwarciem szyjki macicy. W drugim okresie,
następującym bezpośrednio po pierwszym, płód zostaje wydalony z macicy
poza drogi rodne. Okres trzeci kończy się porodem łożyska i błon
płodowych.
Na początku pierwszego okresu porodu odpływają wody płodowe
i rozpoczyna się rozwieranie szyjki macicy. Pełne rozwarcie szyjki macicy
następuje po upływie 8—15 godzin. Okres ten jest krótszy u wieloródek,
a dłuższy u pierworódek. W czasie jego trwania występują regularnie
silne skurcze mięśnia macicznego subiektywnie odczuwane jako bóle porodowe.
W drugim okresie porodu, pod wpływem skurczów mięśnia macicznego
i jednoczesnych skurczów mięśni ściany jamy brzusznej wzmagających
ciśnienie w jamie brzusznej, następuje przesuwanie się płodu przez kanał
rodny i wydalenie go na zewnątrz poza drogi rodne. Płód urodzony
Rozród u płci żeńskiej
511
*~Kość krzyżowa
Ryc. 273. III okres porodu — wydalanie łożyska z jamy macicy.
pozostaje w łączności z organizmem matki za pośrednictwem sznura
pępowinowego. Drugi okres porodu trwa od kilku minut do kilku godzin
i kończy się w momencie odpępnienia płodu. Następuje to wtedy, kiedy
tętnice pępowinowe przestają tętnić. Sznur pępowinowy zostaje przecięty
i łączność pomiędzy noworodkiem i organizmem matki zostaje całkowicie
przerwana.
W trzecim okresie porodu łożysko i błony płodowe zostają wydalone
z jamy macicy poza drogi rodne. Po urodzeniu się płodu w dalszym ciągu
występują skurcze macicy. Powoduje to oddzielenie się łożyska od ściany
macicy. Pomiędzy ścianą macicy a łożyskiem zaczyna się gromadzić krew.
Następujące po sobie kolejne skurcze mięśnia macicznego powodują
wydalenie łożyska z macicy. Zazwyczaj częścią przodującą łożyska w porodach fizjologicznych jest powierzchnia płodowa łożyska. Po urodzeniu
się łożyska poza drogi rodne zostają również wydalone błony płodowe.
Trzeci okres porodu trwa średnio około pół godziny.
512
Rozród
I Połóg i laktacja
Po porodzie następuje okres połogu i laktacji. Okres połogu trwa
6 tygodni od dnia porodu i charakteryzuje się przede wszystkim zwijaniem się macicy. Pod koniec tego okresu macica osiąga zbliżoną wielkość, jaką miała przed ciążą i jej zwijanie zostaje zakończone.
Ryc. 274. Schemat budowy komórki gruczołu sutkowego w czasie wydzielania mleka. (Reprodukowano za zezwoleniem
z W. Sawicki Histologia, II wyd. Wydawnictwo Lekarskie PZWL, Warszawa 1997.)
Laktacja, czyli wydzielanie mleka, rozpoczyna się po upływie 48 godzin od zakończenia porodu i może trwać kilka miesięcy. W końcowym
okresie ciąży i bezpośrednio po porodzie sutki wydzielają siarę. Dopiero
po dwóch dniach rozpoczyna się wydzielanie mleka, w czym bierze udział
prolaktyna (PRL). Hormon ten, wydzielony przez przedni płat przysadki, pobudza komórki gruczołowe gruczołów sutkowych. Natomiast
w wydalaniu mleka bierze udział oksytocyna uwalniana do krwi z tylnego
płata przysadki.
Rozród u płci żeńskiej
51 3
Osesek ssąc brodawkę (papilla mammae) drażni receptory i za pośrednictwem impulsów nerwowych biegnących przez rdzeń kręgowy, rdzeń
przedłużony oraz śródmózgowie zostają pobudzone neurony oksytocynoergiczne podwzgórza. Z ich zakończeń w tylnym płacie przysadki uwalnia
się oksytocyna, która krąży we krwi i dostając się do gruczołów sutkowych, działa kurcząco na ściany przewodów mlecznych. Ten odruch
neurohormonalny sprzyja opróżnianiu się gruczołów sutkowych z nagromadzonego w nich mleka.
17 — Fizjologia człowieka
Skróty i symbole powszechnie
używane w fizjologii
Aa
ABP
ACE
Acetyl-CoA
Ach
AChE
ACTH
ADH
ADP
AEP
AHG
AMP
AMPA
Angll
ANP
APC
APUD komórki
AQRS ARAS
ATP ATP-ase
avD różnica
AVP
aVR, aVF, aVL
adrenalina
krew tętnicza
białka osocza wiążące androgeny enzym
hydrolizujący angiotensynę I do angiotensy-n yl l
acetylo-koenzym A acetylocholina
esteraza cholinowa swoista hormon
adrenokortykotropowy hormon
antydiuretyczny = wazopresyna
adenozynodifosforan słuchowy potencjał
wywołany globulina przeciwhemofilowa
adenozyno-5'-monofosforan
kwas a-amino-3-hydroksy-5-metylo-4-izoksazolopropionowy angiotensyna II
przedsionkowy peptyd natriuretyczny komórka
prezentująca antygen układ komórek endokrynnych
rozsianych wychwytujących prekursory amin
dekarboksylujących je i wydzielających oś
elektryczna serca
układ siatkowaty aktywujący wstępujący
adenozynotrifosforan adenozynotrifosfataza różnica
tętniczo-żylna w stężeniu dowolnej substanwazopresyna argininowa
elektrokardiograficzne odprowadzenia kończynowe
jednobiegunowe, nasilone
Skróty i symbole używane w fizjologii
AV węzeł
AV zastawki
AWR
B
BAEP
BAO
BDNF
BER
BFU-E
BFU-Mk
BMR
BNP
BSP
BUN
(z następującymi
symbolami)
C19 steroidy C21 steroidy
cAMP
51 5
węzeł przedsionkowo-komorowy
zastawki przedsionkowo-komorowe serca
opór dróg oddechowych
— krew
słuchowy potencjał wywołany pnia mózgowia
zawartość kwasu w wydzielinie podstawowej żołądka
czynnik neurotropowy pochodzenia mózgowego
podstawowy rytm elektryczny przewodu pokarmowego
komórka ukierunkowana linii erytrocytów = komórka tworząca kolonie rozsadzające erytroidalne
komórka ukierunkowana linii megakariocytów =
= komórka tworząca kolonie rozsadzające megakariocytarne
podstawowa przemiana materii
mózgowy peptyd natriuretyczny
bromsulftaleina azot mocznika
we krwi
C
CAM
CBF
CBG
CCK
CCK-8
CD
CFC
CFF
CFU-Baso
CFU-E
CFU-Eos
CFU-G
CFU-GEMM
CFU-GM
CFU-M
CFU-Mk
klirens, np. Cin = klirens nerkowy dla inuliny
steroidy zawierające w cząsteczce 19 atomów węgla
steroidy zawierające w cząsteczce 21 atomów węgla
cykliczny adenozyno-3', 5'-monofosforan = cykliczny AM P
cząsteczki białek adhezujących komórki przepływ
krwi przez mózgowie globulina wiążąca
kortykosteroidy (kortyzol), transkortyna
cholecystokinina cholecystokinina oktapeptyd
cząsteczka różnicująca współczynnik filtracji
włośniczkowej częstotliwość krytyczna fuzji
obrazów komórka macierzysta linii bazofilów
komórka macierzysta linii erytrocytów komórka
macierzysta linii eozynofilów komórka macierzysta
linii neutrofilów komórka szpiku macierzysta
nieukierunkowana komórka ukierunkowana
granulocytów i makrofa-gów
komórka macierzysta linii makrofagów/monocytów
komórka macierzysta linii megakariocytów
516
Skróty i symbole używane w fizjologii
cGMP
CGP
CGRP
CH2O CLIP
CMO
CMRO2
CNP
CNS CoA
CO-Hb
COMT
*-PAH
CR
Cr
CRH, CRF
CrP CS
CSF CSF-1
CSF-G
CSF-GM
CSF-M
CT CTM
CTP CV
CVR CZI
d
D
Da
DA
DAG
DBP
DDAVP
DEA
DFP
cykliczny 3', 5'-guanozynomonofosforan = cykliczny GMP
hormon wzrostu kosmówkowy (to samo co hCS)
peptyd pochodny od genu kalcytoninowego
„klirens wolnej wody"
peptyd kortykotropowopodobny części pośredniej
przysadki
pojemność minutowa serca mózgowa
przemiana materii, tlenowa pula neutrofilów
swobodnie krążących we krwi ośrodkowy
układ nerwowy koenzym A
karboksyhemoglobina O-metylotransferaza
katecholowa klirens nerkowy kwasu
paraaminohipurowego odruch warunkowy
kreatynina
hormon uwalniający hormon adrenokortykotropowy = kortykoliberyna = CRF-41 fosforan
kreatyny = fosfokreatyna bodziec warunkowy płyn
mózgowo-rdzeniowy
czynnik wzrostowy pierwszy = czynnik wzrostowy
makrofagów
czynnik wzrostowy granulocytów czynnik
wzrostowy granulocytów i makrofagów czynnik
wzrostowy makrofagów = czynnik wzrostowy
pierwszy kalcytonina
tomografia komputerowa
cytydynotrifosforan
objętość krwi w komorach serca po zamknięciu
zastawek
opór naczyń mózgowych
insulina krystaliczna cynkowa
doba
enancjomer D (zazwyczaj prawoskrętny)
cholekalcyferol
dopamina
diacyloglicerol
białko wiążące witaminę D
l-deamino-8-D-argininowazopresyna
dehydroepiandrosteron = DHEA
diizopropylofluorofosforan = fluostygmina
Skróty i symbole używane w fizjologii
DHD3
DHEA
DHT
DIT
DNA
D/N stosunek
DOC
DOCA
DOMA
DOPA
DOPAC
DPG
DPN
DPNH
DPPC
Dyn
E
(następującymi
cyframi)
EACA
ECF
ECG
Ech
ECoG
EDCF
EDRF
EDTA
EEG
EFP
EGF
EHBF
EJP
517
— 1,25-dihydroksycholekalcyferol
— dehydroepiandrosteron
— dihydrotestosteron
— dijodotyrozyna
— kwas deoksyrybonukleinowy
— stosunek wydalanej z moczem glukozy do azotu
całkowitego moczu
— deoksykortykosteron
— octan deoksykortykosteronu
— kwas 3, 4-dihydroksymigdałowy
— 3, 4-dihydroksyfenyloalanina = L-DOPA
— kwas 3, 4-dihydroksyfenylooctowy
— 2, 3-difosfoglicerynian
— difosfopirydynowy nukleotyd = dinukleotyd nikotynamidoadeninowy (forma utleniona)
— difosfopirydynowy nukleotyd zredukowany = dinukleotyd nikotynamidoadeninowy (forma zredukowana)
— dipalmitoilofosfatydylocholina
— dynorfina
odprowadzenie przełykowe w elektrokardiografii, następujące cyfry określają głębokość w centymetrach,
na jaką elektroda została wprowadzona do przełyku
estron estradiol estriol estetrol
kwas epsilon-aminokapronowy
płyn zewnątrzkomórkowy
elektrokardiogram esteraza
cholinowa elektrokortykogram
śródbłonkowy czynnik zwężający naczynia krwionośne
śródbłonkowy czynnik rozluźniający mięśnie gładkie
naczyń krwionośnych
kwas etylenodiaminotetraoctowy = kwas wersenowy = wersenian elektroencefalografii efektywne
ciśnienie filtracyjne czynnik wzrostu naskórka
obliczony przepływ krwi przez wątrobę potencjał
złącza pobudzający
51 8
Skróty i symbole używane w fizjologii
EKG
ELAM-1
EMG
Enk
Enk-Leu
Enk-Met
EN,
EK
EOG
EP
EPO
EPSP
ERBF
ERG
ERPF
Erys
Eq
ERV
ET-1
ET-2
ET-3
ETP
EV
F
FDP
FEF
FEV
FEV10
FF
FFA
fPSP
FGF
FGFS
FIVC
FIV
FRC
FSH
FVC
G-14
G-17
G-34
GABA
elektrokardiogram
cząsteczki adhezyjne srodbłonkowe leukocytarne
elektromiogram
enkefalina
enkefalina leucynowa
enkefalina metioninowa
potencjał równowagi dla jonów sodowych
potencjał równowagi dla jonów potasowych
elektrookulogram
pirogen endogenny
erytropoetyna
postsynaptyczny potencjał pobudzający
efektywny przepływ krwi przez nerki
elektroretinogram
efektywny przepływ osocza przez nerki
erytrocyty
ekwiwalent
objętość zapasowa wydechowa
endotelina pierwsza
endotelina druga
endotelina trzecia
cząstki transportujące elektrony
objętość wydechowa
— kał
produkty degradacji fibrynogenu natężony
przepływ wydechowy natężona objętość
wydechowa natężona objętość wydechowa w
pierwszej sekundzie frakcja filtracyjna
wolne kwasy tłuszczowe (występują również w skrótach NEFA, UFA)
postsynaptyczny potencjał pobudzający, szybki
czynnik wzrostu fibroblastów czynniki
wzrostu fibroblastów pojemność życiowa płuc
natężona wdechowa objętość wdechowa
natężona pojemność zalegająca czynnościowa
płuc hormon folikulotropowy pojemność
życiowa płuc natężona
gastryna „mała"
gastryna
gastryna „duża"
kwas gamma-aminomasłowy
.
Skróty i symbole używane w fizjologii
GABAA
GABAB
GAD
GBG
GDP
GFR
GH
GIP
GLI
GLP-1
GMP
GnRH
GRH
GRP
GTP
h
Hb
Hb4O2
HBE
25-HCC
hCG
hCS
Het
HDL
HFOV
hGH
fflOMT
HLA
hPL
HR
HS-CoA
5-HT
HVA
IC ICF
519
receptor A kwasu gamma-aminomasłowego
receptor B kwasu gamma-aminomasłowego
dekarboksylaza glutaminowa
globulina wiążąca steroidowe hormony płciowe
guanozynodifosforan
wielkość filtracji kłębuszkowej
hormon wzrostu
peptyd hamujący czynność żołądka
czynnik immunologicznie glukagonopodobny (glicentyna) wyizolowany z błony śluzowej przewodu
pokarmowego
peptyd glukagonopodobny
guanozyno-5'-monofosforan
hormon uwalniający gonadotropiny = gonadoliberyna
hormon uwalniający hormon wzrostu = somatoliberyna, somatokrynina
peptyd uwalniający gastrynę
guanozyno-5'-trifosforan
godzina(y)
hemoglobina odtlenowana
oksyhemoglobina
elektrokardiogram pęczka przedsionkowo-komorowego
25-hydroksycholekalcyferol, aktywny metabolit witaminy D 3 = HD 3
gonadotropina kosmówkowa ludzka
somatomammotropina kosmówkowa ludzka
hematokryt = wskaźnik hematokrytu lipoproteiny
osocza o dużej gęstości sztuczna wentylacja płuc
oscylacjami powietrza o dużej częstotliwości hormon
wzrostu ludzki hydroksyindolowa-Ometylotransferaza antygen leukocytarny ludzki
laktogen łożyskowy ludzki = somatomammotropina
kosmówkowa ludzka częstotliwość skurczów serca
na minutę koenzym A zredukowany 5hydroksytryptamina kwas homowanilinowy
pojemność wdechowa płuc
płyn przestrzeni wewnątrzkomórkowej
520
Skróty i symbole używane w fizjologii
ICV
IDL
IF
IFN
IgA
IgD
IgE
IGF-I, IGF-II
IgG
IgM
IJP
IL
IL-1 — IL-13
In
IP3
IPS
P
IRV
ITP
IVC
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
17-KS
KE
— 17-ketosteroidy
— potas organizmu wymienialny
— wskaźnik filtracji kłębuszkowej
Kr
LATS
LCAT
LDL
LH
LPH
LSD
LT
LTA4
LTB4
LTC4
LTD4
LTE4
LTF4
LTP
LVET
MAO
MBC
MCD
MCH
objętość przestrzeni wodnej wewnątrzkomórkowej
lipoproteidy osocza o pośredniej gęstości
czynnik wewnętrzny żołądkowy
interferon
immunoglobuliny A
immunoglobuliny D
immunoglobuliny E
czynnik wzrostowy insulinopodobny I i II
immunoglobuliny G
immunoglobuliny M
potencjał złącza hamujący
interleukina
interleukina pierwsza — interleukina trzynasta
inulina
inozytolotrifosforan
postsynaptyczny potencjał hamujący
objętość zapasowa wdechowa
inozyno-5'-trifosforan
pojemność życiowa płuc
— długo działający czynnik pobudzający gruczoł tar
czowy
— acylotransferaza lecytynowo-cholesterolowa
— lipoproteiny osocza o małej gęstości
— hormon luteinizujący
— hormon lipotropowy
— dietyloamid kwasu lizerginowego
— leukotrien
— leukotrien A4
— leukotrien B4
— leukotrien C4
— leukotrien D4
— leukotrien E4
— leukotrien F4
— potencjacja długotrwała
— czas wyrzutu lewej komory
—
—
—
—
monoaminooksydaza = oksydaza monoaminowa
maksymalna wentylacja płuc
średnia średnica erytrocytów
średnia masa hemoglobiny w erytrocycie
Skróty i symbole używane w fizjologii
MCHC
MCV
MDLVP
MF
MGP
MHC
MHPG
MIF
MIT
MNP
MRI
ITIRNA
MSH
MSH-R-IH
MT
MV
MW
NA NAD+
NADH
NADP+
NADPH
NaE
Na-K-ATP-aza
NEFA
NGF
NMDA
NO
NPN
NPY
NREM sen
NREM-1
NREM-2
NREM-3
NREM-4
NT
521
średnie stężenie hemoglobiny w erytrocycie
średnia objętość erytrocytu
ciśnienie średnie w lewej komorze serca rozkurczowe
czynnik mitogenny
pula granulocytów przyściennych
główny układ zgodności tkankowej
3-metoksy-4-hydroksyfenyloglikol
czynnik hamujący migrację makrofagów
monojodotyrozyna
pula neutrofilów przyściennych
obrazowanie rezonansu magnetycznego
matrycowy kwas rybonukleinowy (messenger RNA)
= informacyjny RNA
hormon melanotropowy = hormon alpha-, betai gamma-melanotropowy
hormon hamujący uwalnianie MSH = melanostatyna
3-metoksytyramina
maksymalna wentylacja płuc na minutę
maksymalna dowolna wentylacja płuc na minutę
noradrenalina
dinukleotyd nikotynoamido-adeninowy (forma utleniona)
dinukleotyd dihydronikotynoamido-adeninowy (forma zredukowana)
fosforan dinukleotydu nikotynoamido-adeninowego
(forma utleniona)
fosforan dinukleotydu dihydronikotynoamido-adeninowego (forma zredukowana) sód organizmu
wymienialny
adenozyno-trifosfataza aktywowana przez Na + i K+
wolne kwasy tłuszczowe, niezestryfikowane czynnik
wzrostu nerwów N-metylo-D-asparaginian tlenek
azotu azot pozabiałkowy neuropeptyd Y
sen o wolnych ruchach gałek ocznych sen o
wolnych ruchach gałek ocznych, stadium 1 sen o
wolnych ruchach gałek ocznych, stadium 2 sen o
wolnych ruchach gałek ocznych, stadium 3 sen o
wolnych ruchach gałek ocznych, stadium 4
neurotensyna
522
Skróty i symbole używane w fizjologii
OGF
17-OHCS
OMI
Osm
OXY
P P50
PA
PAF
PAH
PAI-1
PAI-2
PAM
PBI
Pc
PDECGF
PDGF
PEEP
PEP
PET
PG
PGA
PGE
PGF
PGI2
PGO
pH
PHI
PHM-27
PIH
P» PIP
PIP2 PMN
czynnik wzrostu jajników
17-hydroksykortykoidy
inhibitor dojrzewania owocytu
osmól = 1 mol związku chemicznego występujący
w 1 L wody
oksytocyna
osocze
prężność tlenu, przy której hemoglobina jest w połowie wysycona tlenem powietrze pęcherzykowe w
płucach czynnik aktywujący trombocyty kwas
paraaminohipurowy inhibitor aktywacji
plazminogenu pierwszy inhibitor aktywacji
plazminogenu drugi makrofagi pęcherzyków
płucnych jod związany z białkiem osocza ciśnienie
w naczyniach włosowatych czynnik wzrostu
śródbłonka pochodzenia z trombo-cytów
czynnik wzrostu z trombocytów ciśnienie końcowe
wydechowe dodatnie w czasie oddychania
okres poprzedzający wyrzut krwi komputerowa
tomografia emisyjna pozytronowa prostaglandyna
prostaglandyny typu A prostaglandyny typu E
prostaglandyny typu F prostacyklina
iglice mostowo-kolankowato-potyliczne w czasie snu
REM
ujemny logarytm stężenia jonów wodorowych w roztworze
peptyd histydylo-izoleucyna
peptyd histydylo-metionina-27 wytwarzany wraz
z VIP z prekursora pre-pro-VIP u ludzi hormon
hamujący uwalnianie prolaktyny = prolaktostatyna
stężenie inuliny w osoczu fosfatydyloinozytol 4,5difosforan fosfatydyloinozytolu granulocyt o jądrze
segmentowanym, ziarnistoś-ciach
obojętnochłonnych, kwasochłonnych lub zasadochłonnych
Skróty i symbole używane w fizjologii
PNMA
PNMT
POMC
PP
PRA
PRC
pre-POMC
pre-pro-AVP-NP II
pre-pro-OXY-NP I
PRF
PRL
PRU
5-PRPP
PTA
PTC
PTH PV
Rap
RAR
Ra,
RAS
RBF
RDS
REM
RES
Rh czynnik
RLP
RMV
RNA
RPF
RQ
rRNA
RV
Rvp
Ry,
s
SA SAR
SCF
523
pierwszorzędowe pole korowe hamujące ruchy
N-metylotransferaza fenyloetanoloaminowa
prężność cząsteczek tlenu
pro-opiomelanokortyna
polipeptyd trzustkowy
reninowa aktywność osocza
stężenie reniny osoczowej
pre-pro-opiomelanokortyna
pre-pro-wazopresyno-neurofizyna druga
pre-pro-oksytocyno-neurofizyna pierwsza
czynnik uwalniający prolaktynę
prolaktyna
jednostka obwodowego oporu naczyniowego
5-fosfororybozylopirofosforan
prekursor tromboplastyny osoczowej = czynnik XI
krzepnięcia krwi
składnik tromboplastyn y os oczowej = czynnik IX
krzepnięcia krwi; również: fenylotiokarbamid
parathormon
objętość osocza krwi
zbiornik tętniczy płucny
mechanoreceptory płuc szybko adaptujące się
zbiornik tętniczy duży
układ siatkowaty wstępujący pobudzający
przepływ krwi przez nerki
zespół oddechowego wyczerpania
faza snu o szybkich ruchach gałek ocznych
układ siateczkowo-śródbłonkowy
aglutynogeny grupowe rhesus erytrocytów
pula limfocytów recyrkulujących
objętość oddechowa minutowa
k was y r yb on uklei n owe = mRNA, tRNA i r RNA
przepływ osocza przez nerki
współczynnik oddechowy
kwas rybonukleinowy rybosomalny
rewers-3, 3', 5'-trijodotyronina = izomer T 3
objętość zalegająca płuc
zbiornik żylny płucny
zbiornik żylny duży
surowica
aktywność właściwa
receptory płuc wolno adaptujące się
czynnik komórek pnia
524
Skróty i symbole używane w fizjologii
SDA
SEP
sEPSP
SGOT
SI
SIDS
SIF neurony
SNMA
SP
SPCA
SPECT
SRIF
SRIF14 SRIF
28 SRIF 28 (1—
12)
sRNA
SSEP
SSMA
SV
3 T4
TBG
TBNP
TBPA
TBV
TBW
TCF
TGF
TGF-a
TLC
Tm
TNF
t-PA
TPN
TPNH
TPO
TPR
TRBF
swoiste dynamiczne działanie pokarmów
dotykowe potencjały wywołane
postsynaptyczny potencjał pobudzający powolny
glutaminianowo-szczawiowo-octanowa transaminaza surowicy krwi
Międzynarodowy Układ Jednostek Miar
zespół nagłej śmierci noworodków
małe neurony zwojów współczulnych o intensywnej
fluorescencji (pośredniczące, dopaminergiczne)
dodatkowe pole korowe hamujące ruchy
Substancja P= tachykinina 1
prokonwertyna = czynnik VII krzepnięcia krwi
tomografia komputerowa emisyjna pojedynczego
protonu
hormon hamujący uwalnianie hormonu wzrostu =
= somatostatyna
somatostatyna 14-peptyd
somatostatyna 28-peptyd
polipeptyd pochodny somatostatyny występujący
w tkankach
rozpuszczalny lub transportujący RNA
dotykowy potencjał wywołany
dodatkowe pole korowe czuciowo-ruchowe
objętość wyrzutowa serca
3, 5, 3'-trijodotyronina
tyroksyna
globulina wiążąca tyroksynę
całkowita pula neutrofilów krwi
prealbumina wiążąca tyroksynę
całkowita objętość krwi
całkowita woda organizmu = objętość całkowitej
przestrzeni wodnej organizmu
płyn transkomórkowy
czynnik wzrostowy transformujący
czynnik wzrostowy transformujący alpha
pojemność płuc całkowita
transport maksymalny w kanalikach nerkowych
czynnik martwicy nowotworów
aktywator tkankowy plazminogenu
nukleotyd trifosfopirydynowy (forma utleniona)
nukleotyd trifosfopirydynowy (forma zredukowana)
trombopoetyna
całkowity obwodowy opór naczyniowy
całkowity przepływ krwi przez nerki
Skróty i symbole używane w fizjologii
hormon uwalniający hormon tyreotropowy = tyreoliberyna
transportujący kwas rybonukleinowy czynnik
pobudzający trombopoezę = trombopoetyna hormon
tyreotropowy = tyreotropina immunoglobuliny
pobudzające gruczoł tarczowy całkowity czas snu
nocnego o wolnych ruchach gałek ocznych
całkowity czas snu nocnego o szybkich ruchach gałek ocznych objętość oddechowa tromboksan
TRH
tRNA
TSF
TSH
TSI
TSN
TSR
TV
TxA
U
(z następującymi
symbolami)
UL uPA
UR
URF
us
US G
V, • V 2 • itd.
VA
vc
VCAM-1
VD
VEGF VF
VEP
VIP
VL
VLDL
VMA
VOR
VPRC
VR
VT
525
stężenie w moczu np. U kr = stężenie kreatyniny
w moczu
nieruchoma warstwa (płynu)
aktywator plazminogenu urokinazowy
odruch bezwarunkowy
czynnik rozkurczający macicę = relaksyna
bodziec bezwarunkowy
ultrasonografia
jednobiegunowe odprowadzenia przedsercowe EKG
= odprowadzenia Wilsona wentylacja minutowa
pęcherzyków płucnych pojemność życiowa płuc
cząsteczki adhezyjne komórek naczyniowych
objętość przestrzeni martwej fizjologicznej czynnik
naczyniowy wzrostu śródbłonka jednobiegunowe
odprowadzenie EKG z lewej kończyny dolnej
wzrokowe potencjały wywołane wazoaktywny
peptyd jelitowy jednobiegunowe odprowadzenie
EKG z lewej kończyny górnej
lipoproteidy osocza o bardzo małej gęstości kwas
wanilino-migdałowy = kwas 3-metoksy-4-hydroksymigdałowy
maksymalna objętość tlenu pobranego na minutę
odruch przedsionkowo-oczny wskaźnik
hematokrytu
jednobiegunowe odprowadzenie EKG z prawej kończyny górnej wentylacja minutowa płuc
526
Skróty i symbole używane w fizjologii
v\VF
— czynnik von Willebranda
W
— czuwanie
— leukocyty = krwinki białe
X chromosom
Y chromosom
— chromosom X płciowy u ludzi
— chromosom Y płciowy u ludzi
Skróty aminokwasów występujących w białkach
527
SKRÓTY AMINOKWASÓW WYSTĘPUJĄCYCH W BIAŁKACH
Skrót
trzyliterowy
Symbol
jednoliterowy
Ala
Arg
Asn
Asp
Asx
Cys
Gin
Glu
Glx
Gly
His
Ile
Leu
Lys
Met
Phe
Pro
Ser
Thr
Trp
Tyr
Val
A
R
N
D
B
C
Q
E
Z
G
H
I
L
K
M
F
P
S
T
W
Y
V
Aminokwas
alanina
arginina
asparagina
kwas asparaginowy
asparagina lub kwas asparaginowy
cysteina
glutamina
kwas glutaminowy
glutamina lub kwas glutaminowy
glicyna
histydyna
izoleucyna
leucyna
lizyna
metionina
fenyloalanina
prolina
seryna
treonina
tryptofan
tyrozyna
walina
528
Mianownictwo steroidów
MIANOWNICTWO STEROIDÓW
Nazwa medyczna
Nazwa systematyczna — chemiczna
Cholesterol
Pregnenolon
17<x-hydroksypregnenolon
Dehydroepiandrosteron (DHEA)
S-DHA
A5-cholesten-3 p-ol
As-pregnen-3 P-ol, 21-on
A5-pregnen-3p, 17a-diol, 20-on
A5-androsten-3 P-ol, 17-on
A5-androsten-3 a-ol, 17-on
Progesteron
17-a-hydroksyprogesteron
Deoksykortykosteron (DOC)
Kortykosteron
11-dehydrokortykosteron
11 - deoksykortyzon
Kortyzol
Kortyzon
Androstendiol
Testosteron
Androsteron
A4-pregnen-3, 20-dion
A4-pregnen-17 a-ol, 3,20-dion
A4-pregnen-21-ol, 3,20-dion
A4-pregnen-lip, 21-diol, 3,20-dion
A4-pregnen-21-ol, 3,11,20-trion
A4-pregnen-17 a-ol, 3,20-dion
A4-pregnen-lip,17a, 21-triol, 3,20-dion
A4-pregnen-17a, 21-diol, 3,11,20-trion
A5-androsten-3a, 17p-diol
A4-androsten-17 P-ol, 3-on
androsten-3 a-ol, 17-on
Estron (E,)
Estradiol (E2)
Estriol (E3)
Ąl,3,5:10.estratrien.3.ol;
Androstendion
11-P-hydroksyandrostendion
Aldosteron
A4-androsten-3, 17-dion
A4-androsteron-ll P-ol, 3,17-dion
A4-pregnen-18 formylo, lip, 21-diol, 3,20-dion
17.on
A'^'O-estratrien-3-ol, 17p-ol
Ai. 3,5:1 o.estratrien-3-ol, 16a, 17P-diol
Skorowidz
A-receptory 127
Acetylocholina 76, 108, 142, 216, 221
— znaczenie dla przepływu wieńcowego 359
Adaptacja do zapachu 132
— siatkówki 144
— — do ciemności 144
----- do silnego światła 144
Adenozynotrifosfataza aktywowana przez sod i potas 26
Adenozynotrifosforan 44, 98, 415
Adenylocyklaza 218 Adrenalina,
działanie 249
— gromadzenie i wydzielanie z pęcherzyków rdze
nia nadnerczy 246(r)
Akomodacja oka 134
Akson(y) 64(r)
— powstawanie impulsów 70
Aktywa 90, 91(r)
Aktywacja enzymów proteolitycznych 162
— plazminogenu 330, 332
Aktywiny 259
Aktywność somatyczna, mechanizmy kierujące
201
----- ośrodki wyzwalające 201
Albuminy 323
Aldosteron 244
Aminokwasy pobudzające błonę postsynaptyczną
78
— w osoczu 325
— wchłanianie w przewodzie pokarmowym 460
Aminy katecholowe, wydzielane przez rdzeń nad
nerczy 246
Amoniak w osoczu 325
Amylina ludzka 262
Anabolizm 18 Androgeny
245 Angiogenina 271
Angiotensyna 385
Angiotensynazy 285
Angiotensynogen 266, 385
Angiotensyna II, wytwarzanie 493(r)
Anion(y) wodorowęglanowe 473
Ankiryna 300
Antygen(y) D 304
— głównego układu zgodności tkankowej 293
— grupowe A BO 304
— Jk 305
— kell 305
— ludzki leukocytarny 294, 305
— S305
Antykodon 38
Antyport 26
Aparat Golgiego 38
Apoferrytyna 461
Apoptoza 49
Arginina 238
Astrocyty 89, 394
Atropina 218, 222
Audiogram 154(r)
B-receptory 129
Badanie ultrasonograficzne serca 354
— wzrokowych potencjałów wywołanych 119(r)
Bariera mózgowa 394
Baroreceptory 383 Bazofile
285, 312 Beta-endorfina 165
Białko(a) aktywujące Gs 32
— błonowe G 32
— błony komórkowej 23
— histonowe 36
— integralne 24
— niehistonowe 37
— nośnikowe 24
530
Skorowidz
Białko(a) osocza 321, 322(t)
-----synteza w komórkach wątrobowych 467(r)
— receptorowe 24
— transbłonowe 28(r)
Bilirubina 304, 464
— schemat krężenia 465(r)
— w osoczu 325
Biliwerdyna 304
Biosynteza hormonów gruczołu tarczowego
250(r)
— immunoglobulin 313
Bit 53
Błędnik 169
— błoniasty 151(r)
— budowa 170
— czynność 170
Błona(y) bębenkowa 151
— komórkowa 23, 24(r)
ciała neuronu 83(r)
-----przenikanie cząsteczek 25(r)
-----transport 24
— postsynaptyczna 67
— presynaptyczna 67
Bodziec(e) 157
— odbieranie 115
— progowe dla receptorów ciepła 162
zimna 162
— słuchowe, odbiór 154
Bradykinina 164
Centromer 356
Chemodetektory w rdzeniu przedłużonym 414
Chemokiny 310
Chemoreceptory oddechowe 412
— w kłębuszkach szyjnych i kłębkach aortowych
413(r)
Chemotaksja 307(r)
Chlor 473 Chłonka 333
— krążenie 373
Cholecystokinina 437, 445, 446, 451
Cholekalcyferol, metabolizm 493
Cholesterol 457 Chromosom(y) 34
— duplikacja 34
Chylomikrony 457
Ciałko(a) blaszkowate 159
— buławkowate 129, 169
— dotykowe 159
— Golgiego 169
— Meissnera 159
— nerkowe 474
— Paciniego 159, 169
— Ruffiniego 162
Ciało kolankowate boczne 147
Ciało kolankowate przyśrodkowe 157
— neuronu 63(r)
Ciepło, drogi odprowadzenia z organizmu 272
— sposoby wytwarzania przez organizm 273
Ciśnienie filtracyjne w naczyniach włosowatych 373
— krwi u człowieka leżącego 360(r)
----------- stojącego 37O(r)
---- w aorcie 363(r)
— onkotyczne białek w naczyniach włosowatych
373
— parcjalne O2 CO2 408(t)
---- tlenu i dwutlenku węgla w pęcherzykach
płucnych 404(r)
— resorpcyjne w naczyniach włosowatych 373
— rozkurczowe 361
— skurczowe 361
— śródczaszkowe 392
— tętnicze krwi 360
-------- regulacja 383
---- średnie 361
— w jamach serca i zbiornikach tętniczych 351 (t)
— w zbiorniku żylnym dużym 369
— żylne centralne 369
---- obwodowe 369
Cykl(e) jajnikowy 503
— komórkowy 34(r), 48
— kwasów trikarboksylowych 416(r)
— kwasu cytrynowego, enzymy 46
— miesiączkowy bezowulacyjny 503
— pentozowy 418(r)
— płciowy żeński, hormonalne sprzężenie zwrot
ne 502
— pracy serca 349(t)
Cytokiny 269, 311(t)
Czas ośrodkowego przewodzenia 121
— trwania załamków odcinków i dostępów w
EKG 345(t)
Cząsteczki różnicujące 292
Część depresyjna ośrodka naczynioruchowego w
rdzeniu przedłużonym 381(r)
— presyjna ośrodka naczynioruchowego w rdze
niu przedłużonym 380(r)
Czop płytkowy w uszkodzonej ścianie naczynia
krwionośnego 327(r)
Czopki 139, 140 Czucie
129, 130
— bólu trzewnego 175
powierzchniowego 162
— dotyku i ucisku 159
— drogi nieswoiste dla impulsów 178
— eksteroceptywne 157
— interceptywne 174
— podział 130
— proprioceptywne 167
— smaku 167
— teleceptywne 131
Skorowidz
Czucie trzewne 174
— zimna i ciepła 162
Czuwanie 197
Czynnik(i) luteolityczny 507
— czuwania 197
— działające na komórki tkanki kostnej 472(t)
— krzepnięcia krwi 329(t), 466
— luteolityczny macicy 257
— martwicy nowotworów alpha 271
— naczyniowe wzrostu śródbłonka 271
— pobudzające i hamujące działanie endoteliny,
schemat działania 388(r)
— snu 197
— von Willebranda 328
— wpływające na zagęszczenie moczu 490
— wzrostowe 269(t)
— angiogenetyczne 271
fibroblastów 271
---- hematopoetyczne 270, 280(t)
— wzrostowy granulocytów 285
--------i makrofagów 270, 285
---- insulinopodobny I i II 237
---- pierwszy 285
— wzrostu 268
— — nerwów 64
---- śródbionka pochodzenia płytkowego 271
---- tkankowe 268
Czynność bioelektryczna mózgu samoistna 118
----------- wywoływana 118, 119
— gruczołu tarczowego 250
— mechaniczna serca 348
— nerwowa wyższa 207
— siatkówki 139
Dehalogenaza jodotyrozynowa 251
Dehydroepiandrosteron 245
Depolaryzacja 71(r)
— skokowa 84
Desymilacja 18
Desynchronizacja 117
Diacyloglicerol 33, 218
Diapedeza 3O7(r), 310
Diureza osmotyczna 490
— wodna 490
DNA, podwójna helisa 35(r), 36
Dopamina 78, 143, 219, 239
Droga(i) czuciowe nieswoiste 130
-------swoiste 130
— i ośrodki czuciowe swoiste 176(t)
— impulsów dla ruchu 183
---- słuchowych 155
---- z błędnika 172
---- z siatkówki do kory mózgu 147(r)
---- z układu ruchu 169
531
Droga(i) przewodząca impulsy z receptorów w skórze do kory mózgu 163(r)
— przewodzące impulsację słuchową 156(r)
-------- węchową 132(r)
Dyfuzja ułatwiona 25
Dynorfina 81, 165
Dysocjacja hemoglobiny, wpływ dwutlenku węgla
303(r)
-------- temperatury 302(r)
Działanie pokarmów swoiste dynamiczne 422
Echokardiografia 354
Efektory 115
Egzocytoza 43(r), 44
Egzony 37
Eikonazoidy 477
— miejsce powstawania i działania 267(t)
Ejakulacja 500
Eksternalizacja białek receptorowych 27
Elektrody odbierające 116
Elektroencefalogram 116
- człowieka 117(r), 198(r)
Elektroforeza bibułowa białek surowicy krwi
323(r)
Elektrogeneza potencjałów korowych 118
Elektrokardiogram 344
— odprowadzenia kończynowe 339
Elektrokortykogram 116
Elektrolity 471
Elektrookulogram 145
Elektroretynogram 146, 146(r)
Elementy morfotyczne w szpiku kostnym czerwonym i we krwi obwodowej 281(t) Elongacja 38
Endocytoza 42 Endotelina 389
— schemat cząsteczki 388(r)
Energetyka skurczu mięśnia 98
Enkefalina leucynowa 81, 165
— metioninowa 81, 165
Enterocyty 455
Eozynofile 285, 311
Erytroblast(y) ortochromatyczne 282
— polichromatofile 282
— zasadochlonne I i II 282
Erytrocytopoeza 282
— w szpiku kostnym czerwonym 283(r)
Erytrocyty, ciśnienie osmotyczne 300
— cytoszkielet 300
— funkcja 298
— wartości średnie charakteryzujące 299(t)
Erytropoetyna 282, 493
Esteraza cholinowa 105
Estradiol 245, 255, 408
Estriol 255, 508
532
Skorowidz
Estrogeny 255, 503, 508
— działanie 256
— regulacja wydzielania w fazie folikularnej 256(r)
— wolne 255
Estron 255, 508
Fagocytoza 42, 43(r), 3O7(r)
Fala(e) biegnące 153
— ciśnienia w przychlonce 153
— dykrotyczna 365
— perystaltyczna 431
— tętna 363
— EEG 116
Faza(y)Gl 48
— GO 48
— folikularna cyklu miesiączkowego 255, 505
— lutealna cyklu miesiączkowego 255, 505
— połykania 427
— S48
— snu 199
Fenoksybenzamina 218
Fenyloalanina, przemiana w organizmie 217(r)
Ferrytyna 461 Fibrynogen 324, 466
— stabilny 332
Fibrynoliza, schemat 332(r)
Filtr wątrobowy 464
Filtracja i resorpcja w sieci naczyń włosowatych
372(r)
— ktębuszkowa 479
— w naczyniach włosowatych 373
Fonokardiogram 357, 357(r), 364(r)
Fotostymulator 119(r)
Frakcja filtracyjna 482
Gałązki kolateralne włókien aferentnych 178
— oboczne włókien aferentnych 178
Gałka oczna 134(r)
— budowa 134
— rodzaje wykonywanych ruchów 149
Gastryna 437, 451
Glicentyna 454
Glikogen 466
— wątrobowy 263
Glikokaliks 28(r)
Glikokortykoidy 242
Glikoliza46, 416
— beztlenowa 98, 417
Globina 300
Globuliny 323
Glukagon 262, 454
Glukoza 483
— regulacja zawartości we krwi 262(r)
Gonadotropina łożyskowa ludzka 509
Gorączka 275
Granice fizjologiczne 4l>
Granulocytopoeza 285
— w szpiku kostnym czerwonym 286(r)
Granulocyty 307
Grasica, budowa 295(r)
— hormony 264
— komórki zrębu 294 Gromadzenie energii
w komórce 415 Gruczoł(y) błony śluzowej
jelita grubego 447
— dokrewne, czynność 225
— dwunastnicze podśluzówkowe 440
— jelitowe 440
— Lieberkuhna 440
— przytarczyczne, hormony 260
— tarczowy, hormony 249
---- transport hormonów 251
Grudki chłonne 296
Grupa(y) krwi, podział 304, 3O5(t)
— układu Rh 304
Haloperidol 219
Hamowanie czucia bólu 165, 166(r)
— presynaptyczne 71(r) 81, 165
— wewnętrznych odruchów warunkowych 206
Heksametonium 218, 222
Hem 300
Hematokryt 277
Hematopoeza w szpiku kostnym czerwonym 279(r)
Hemofilia 332
Hemoglobina 300
— budowa cząsteczki 301(r)
— rola 303
Hemoliza erytrocytu 300
Hemostaza 326 Heparyna
312, 333 Hepatocyty 463
Hiperpolaryzacja 71(r)
Hipnogram 200(r)
Hipokamp 203 Histamina
266, 312 Homeostaza 22
Hormon(y) 225
— działające na tkanki przez aktywację lub inaktywację cyklazy adenylanowej 31(t)
— folikulotropowy 257, 259, 503
— gonadotropowe 239
— grasicy 264
— gruczołu tarczowego, regulacja wydzielania 252
rola 254
------- transport 251
-------- udział w termoregulacji 274(r)
— i przekaźniki chemiczne w przewodzie pokar
mowym, działanie 454
— jajnikowe, regulacja wydzielania 257
Skorowidz
Hormon(y) kory nadnerczy 242
— lipotropowy 240
— luteinizujący 257, 258, 503
— melanotropowe 81, 241
— peptydowe żolądkowo-jelitowe 265
— pochodne aminokwasów 225
— podwzgórzowe 227, 228(t)
---- działające hamująco 234
--------pobudzająco 233
— polipeptydowe 466
— przedniego piąta przysadki 236(t)
— rdzenia nadnerczy 246
rola 248
— steroidowe pochodne cholesterolu 226
— szyszynki 263
— tkankowe 265
— tropowe 240
— tyreotropowy 252
— wydzielane przez korę gruczołu nadnerczowego 244(t)
— wzrostu 237
— żolądkowo-jelitowe 432, 450
---- działanie 444(r), 452, 453(t)
Hydroliza ATP 416(r)
Immunoglobuliny, biosynteza 313
— budowa 316(r)
Impuls(y) nerwowy 63, 66
— słuchowe, przebieg 155
— sumowanie 67
Impulsacja eferentna 159(r)
— współczulna i przywspółczulna, działanie 223(t)
— z błędnika, drogi przewodzące 173(r)
Inhibina(y) 259, 503
Inozytolotrifosforan 33, 218
Insulina 261
— rola 263
Interleukina(y) 309(t)
Interleukina 3 270
Interleukina 8 271
Interleukina 1 197, 235
Internalizacja receptorów 28
Interoreceptory, odbiór bodźców 174
Introny 37
Izoimmunizacja 304
Jądro(a) brzuszne i grzbietowe ciała czworobocznego 157
— komórkowe, oddziaływanie steroidów na apa
rat genetyczny 41(r)
— ruchowe 125
Jednostka motoryczna 99
Jelito cienkie, motoryka 437
trawienie 440
533
Jelito czcze 455
— ślepe 447
motoryka 448
Jon(y) magnezowe, działanie 473
— wapniowe 33
Kalcytonina 255
Kalikreiny tkankowe 162
Kalmodulina 34, 106
Kamyczki błędnikowe 171
Kanaliki nerkowe 474
----- wchłanianie jonów 484, 485(r)
-------- wody 486(r)
Kanał(y) dla prądów jonów sodowych 61
— informacyjny 53
Karboksyhemoglobina 303
Katabolizm 18
Kationy i aniony w osoczu, płynie tkankowym
i wewnątrzkomórkowym 21(t)
Kininogeny 162 Kininy 164 Klirens
dla inuliny 479
— nerkowy 478
Kłębuszek(i) naczyniowy 474
----- ciałka nerkowego, rozkład ciśnień 480(r)
---- przepuszczalność 480
Kod genetyczny 38—41
Kodon 38
Kolagen 328
Kolba(y) końcowa 162
------- synaptyczna 73
— Krausego 162
Komórka(i) aparatu przykłębuszkowego nerek 385
— bariery szpiku kostnego 279
— Betza 180
— budowa 23(r)
— czynność 22
— dodatkowe gruczołów błony śluzowej 430
— dokrewne G 437
— G wewnątrzwydzielnicze 451
— glejowe 88
— główne gruczołów błony śluzowej 429
— kwasochłonna przysadki 239(r)
— macierzysta linii bazofilów 281, 285
--------eozynofilów 285
--------makrofagów 281
--------neutrofilów 281
— — makrofagów ukierunkowana 287
megakariocytów 287
---- nieukierunkowane szpiku 280
— mezangium 477
— mięśniowa gładka 106
---- poprzecznie prążkowana 89
---- ekstrafuzalne 100, 127
---- intrafuzalne 101, 127
534
Skorowidz
Komórka(i) mięśniowa, pobudliwość 59
— raikrogleju 89
— nerwowa 62(r)
----- piramidalne olbrzymie 180
----- pobudliwość 59
— nerwowo-zmysiowa węchowa 131
— odbiór informacji 29
— okładzinowe gruczołów błony śluzowej 429
— pluripotencjalna hematopoetyczna pnia 280
— płciowe męskie 498
żeńskie 498
— prezentujące antygen 314, 317
— progenitorowe limfoidalne 281
— siateczki 279
— siateczkowo-śródbłonkowe gwiaździste 463
— smakowe 167
— somatyczne 48
— zmysłowa włoskowata 154, 170
— zrębu grasicy 294
Komparator w układzie sterującym 57
Komunikacja autokrynna 54
— endokrynna 54
— międzykomórkowa 54
— parakrynna 54
Konekson 54(r)
Konsolidacja pamięci 204
Kontrola jednokierunkowa 57
— ruchów gałki ocznej 149
— wielkości źrenicy 136
— wzajemnie zwrotna 57
Konwergencja oczu 136
Kora mózgu człowieka, umiejscowienie funkcji 209(t)
----- pole dodatkowe czuciowo-ruchowe 181
------------ tłumiące ruchy ciała 182
-------- pierwszorzędowe tłumiące ruchy 181
Kortykosteron 242
Kortyzol 242 Korzeń
grzbietowy 122 Kosteczki
słuchowe 151 Kowadełko
151 Krążenie 50
— jelitowo-wątrobowe 447, 466
— krwi, ośrodki kontrolujące 375
w sieci naczyń włosowatych 372(r)
— płucne 389, 390(r)
— płynu mózgowo-rdzeniowego 393
— wieńcowe czynniki wpływające 359
— wrotne 394, 395(r)
Kreatynina w osoczu 325
Krew, elementy morfotyczne 298
— krzepnięcie 328
— rola w organizmie 277
— skład 276
Krzepnięcie krwi 328
Krzywa przepływu cewkowego 497
Krzywizna(y) rogówki 134
Krzywizna(y) soczewki 134
Kubki smakowe 167
Kwas(y) 3-metoksy-4-hydroksymigdałowy 217
— deoksyrybonukleinowe 36
— foliowy 284
— gamma-aminomasłowy 79, 197
— glikocholowe 446
— moczowy w osoczu 325
— rybonukleinowe 37
— taurocholowe 446
— tłuszczowe wolne 464
— żółciowe 446, 468
Laktacja, wpływ na wydzielanie prolaktyny 239
Laktogen łożyskowy ludzki 509 Leptyna 424
Leukocyty 306
— we krwi obwodowej 306(t)
Leukotrieny 266
Ligandy 24, 28
Limfocytopoeza 290
— w centralnych tkankach limfoidalnych 291 (r)
Limfocyty 312
— B-szpikozależne 292, 313
— dojrzewanie w centralnych i obwodowych na
rządach limfoidalnych 292(r)
— naturalni niszczyciele 292, 294, 313
— pre-B 281
— pre-T281
— recyrkulujące 292
— T-grasiczozależne 312
Limfokiny 269
Lipaza lipoproteinowa 458
Lipoproteiny osocza 326
----- miejsce powstawania i skład 460(t)
Lizosomy 42
Łańcuch oddechowy, enzymy 44
Łożysko, czynność 507
— krwionośne 276
Łuk odruchowy 121, 122
----- autonomiczny 214
-------- część ośrodkowa 215(r)
----- odruchu monosynaptycznego 123
-------- polisynaptycznego 123
-------- rdzeniowego 122(r)
-------- warunkowego 207
----- somatyczny, monosynaptyczny 124(r)
----- trzewny 175
-------- przełączanie się impulsacji 175(r)
Magnez 471
Makrofagi 287
Markery 293
Skorowidz
Mechanizm(y) fizjologiczne 17
— krzepnięcia krwi, zewnątrzpochodny 330
wewnątrzpochodny 328
— molekularny działania adrenaliny na hepatocyty 248(r)
-------- hormonów o budowie peptydowej 226(r)
-------- hormonów steroidowych 227(r)
----- skurczu komórki mięśniowej poprzecznie
prążkowanej 93(r)
------- wydzielania hormonów rdzenia gruczołu
nadnerczowego 247(r)
— ślizgowy skurczu komórki mięśniowej 94(r)
Mechanoreceptory deflacyjne 414
— inflacyjne 413
Melatonina 197, 263, 502
Metabolizm 18
— komórkowy, samoregulacja 48
Metarodopsyna 139
Methemoglobina 303
Metoda pomiaru przepływu cewkowego 497
Metylotransferaza katecholowa tlenowa 216
Miazga biała śledziony 297
— czerwona śledziony 297
Mięsień(e) gładki, pobudzenie 106(r)
trzewne 106
----- wielojednostkowe 106
— napinacz błony bębenkowej 155
— rozwieracz źrenicy 136
— rzęskowy 135
— sercowy, budowa 110
— strzemiączkowy 155
----- czynności bioelektryczne i mechaniczne
HO(r)
----- odżywianie 360
----- wpływ czynników nerwowych i humoralnych 354
— zwieracz źrenicy 136
Mikcja 497
— ośrodki nerwowe kontrolujące 495(r)
Mineralokortykoidy 244, 490
Miofibryla 89, 92(r)
Miozyna 90, 91(r)
Mitochondria 415
— budowa 45(r)
Mitoza 48
Młoteczek 151
Mocz, tworzenie 490
— wydalanie 494
Mocznik, biosynteza w komórkach wątrobowych
476(r)
— w osoczu 325
Modulacja aktywności ośrodka wdechu 412
Modulator(y) 74
— synaptyczne 79, 143, 195
— miejsce występowania w ośrodkowym i ob
wodowym układzie nerwowym 80(t)
535
Modyfikacja prekursora pre-pro-opiomelanokortyny w komórkach przysadki 241(t)
Monoaminooksydaza 44
Monocytopoeza 287
— w szpiku kostnym czerwonym 286(r)
Monocyty 287, 316
Monosacharydy 464
Motoryka jelita cienkiego, kontrola 439
------- grubego 448
— pustego żołądka 432(r)
— żołądka, kontrola 433
Motylina 454
Mózg, czynność bioelektryczna 116
— synchronizacja czynności elektrycznej 117
Móżdżek, czynność 187
Naczynie(a) chłonne 333
— oporowe, czynność 366(r)
rola 365
— włosowate, ciśnienie filtracyjne 373
filtracja 371
Nadstrzał 69 Narząd
Cortiego 150
— spiralny 150
Nekroza 48
Neoendorfina 165
Nerka(i), czynność 474
— wchłanianie wody 487
Nerw błędny 444
— rdzeniowy 122
— trzewne 468
----- miedniczne 449
Neuron(y) amakrynowe 141
— cholinergiczne 192
— dopaminergiczne 194
— dwubiegunowe 141
— funkcja 63
— GABA-ergiczne 195
— glicynoergiczne 195
— gruszkowate 189
— istoty szarej środkowej śródmózgowia, rola w
hamowaniu czucia bólu 165
— międzysplotowe 141
— neurosekrecyjne podwzgórza 229(r)
— noradrenergiczne 192
— poziome 141
— ruchowe, funkcje 125
— serotoninergiczne 194
— wzrokowe czopkonośne 139, 141
pręcikonośne 139
— wzrokowo-zwojowy 141
Neuropeptyd(y) opioidowe 196
— Y424
Neuroplazma 63
Neutrofile 285, 308
536
Skorowidz
Nici chromatydowe 34, 36
Niezborność oka 137 Nitki
kurczliwe 106 Noradrenalina 78,
108, 216, 264
— gromadzenie w zakończeniach adrenergicznych
75(r)
— wpływ na komórkę mięśnia gładkiego 109(r)
Objętość krwi całkowita 276
— oddechowa płuc 398
— wyrzutowa serca 351
— zalegająca płuc 400
— zapasowa wdechowa płuc 398
----- wydechowa płuc 399
Obwódka rzęskowa 135
Oczopląs fizjologiczny 150
— kaloryczny 174
— optokinetyczny 173
— podczasobrotowy 173
— poobrotowy 173
Odbiór bodźców słuchowych 154
— konturów przedmiotów w polach wzrokowych
kory mózgu 149
Odczyny poprzetoczeniowe 304
Oddawanie kału, łuki odruchowe 450(r)
----- mimowolne 449
Oddychanie 49
— ośrodki nerwowe sterujące 396
— regulacja 410
— wewnętrzne 45, 415
— wybuchowe neutrofilów 310
— zewnętrzne 396
Odpowiedź immunologiczna pierwotna 313, 315(r)
----- wtórna 315
Odprowadzenia jednobiegunowe przedsercowe
Wilsona, schemat 342(r)
— kończynowe dwubiegunowe w elektrokardio
grafii 340(r)
Odruch(y) 121
— Babińskiego 129
— Bainbridge'a 378
— Bezolda-Jarischa 378
— bezwarunkowy 121
— bębenkowe 155
— Heringa-Breuera 414
— masowe 129
— monosynaptyczny 123
— neurohormonalny, wpływ na ciśnienie tętnicze
krwi 384
----- wydzielania wazopresyny 230(r)
— polisynaptyczny 123
— trzewno-mięśniowe 178
— warunkowe 121, 206
— własne rdzenia kręgowego 127
— żołądkowo-jelitowy 437
Odruch(y) żołądkowo-krętniczy 438
— żołądkowo-okrężny 448
Odżywianie 49, 423
Okluzja 126
Oko, akomodacja 134
— konwergencja 136
— miarowe 136, 138(r)
— niemiarowe 136
----- krótkowzroczne 136
----- nadwzroczne 136
— siła rozdzielcza 143
Okres(y) bezwzględnej niewrażliwości 87, 95
------------ mięśnia sercowego 112
— porodu 510
— refrakcji bezwzględnej 70
Oksydaza monoaminowa 217
Oksyhemoglobina 301, 405
Oksytocyna 231, 512
— cząsteczka 232(r)
— powstawanie genu 40(r)
Oksytocynaza 231
Oligodendrocyty 89
Opony mózgowo-rdzeniowe, budowa 393(r)
Opór naczyniowy 367
----- obwodowy całkowity 352, 367
----- płucny całkowity 368
Opóźnienie synaptyczne 126
Opsyna 139
Organizacja neuronalna somatotopowa 161
Osłonka mielinowa 65
Osmolarność 20
Osocze krwi 276
----- skład 319(t)
— lipidy 325
— składniki organiczne pozabiałkowe 325
Ostrość widzenia 140
— wzroku 143
Oś elektryczna serca 347
— patrzenia 133
— układu optycznego oka 133
Ośrodek(i) agresji i ucieczki 202
— głodu 424
— kierujące zachowaniem 201—203
— mikcji w moście boczny 496
przyśrodkowy 494
— mowy 208
nadrzędny 208
----- w korze mózgu człowieka 208(r)
— naczynioruchowy 378
----- część presyjna 379
-------- depresyjna 382
— nerwowe 124
— oddechowy 410
— pneumotaksyczny 412
— pokarmowe 201, 424
— pragnienia 202
Skorowidz
Ośrodek(i) przyspieszający pracę serca 375, 376(r)
— rozrodczy 203
----- żeński w podwzgórzu 502
— ruchowy mowy Broca 208
— ruchów pisarskich ręki 208
— sercowy 375
— słuchowy mowy 208
— sytości 424
— termoregulacji 273
— wdechu 410
— wzrokowy korowy pierwszorzędowy 149
mowy 208
— zwalniający pracę serca 376(r), 377
Otolity 171
Pamięć natychmiastowa 204
— proceduralna 205
— świeża 204
— trwała 204, 205
----- rodzaje 205(r)
Parathormon 260
— regulacja wydzielania 260(r)
Pepsynogen 429
Peptyd hamujący czynność żołądka 454
— jelitowy wazoaktywny 454
— kortykotropowopodobny 81
— natriuretyczny, cząsteczka 491(r)
— opioidowe 81, 165
— przedsionkowy natriuretyczny 214
— wazoaktywny jelitowy 222
Percepcja 130
— wrażeń wzrokowych 148
Pęcherzyk(i) jajnikowe pierwotne 505
dojrzały 505
----- wzrastające 505
— synaptyczne 44, 73
Pęczek noradrenergiczny brzuszny 192
------- grzebietowy 192
— przedsionkowo-komorowy 113, 337
Pętla mięśniowa 102
— sprzężenia zwrotnego 57
-------- zewnętrznego 502
Pinocytoza 42, 43(r)
Plamka przedsionka z komórkami receptorowymi, schemat 171(r)
— ślepa 144
Plemniki 499
Pleć genetyczna 498
— gonadalna 498
Płuca, dyfuzja gazów 403
Płyn(y) hipotoniczny 470
— hipertoniczny 470
— izotoniczne 470
— mózgowo-rdzeniowy, krążenie 393
— ustrojowe 276
— w przestrzeni transkomórkowej 469
537
Płyn(y) w przestrzeni wewnątrzkomórkowej 469
----------- zewnątrzkomórkowej 469
— wewnątrzkomórkowy 19
— zewnątrzkomórkowy 19, 470
Płytki krwi 318
Pneumocyt typu II, budowa 402(r)
Pobudliwość 59
— ośrodków rdzenia kręgowego 125
Pobudzenie komórek 59
Pochodne kwasu arachidonowego 266
Podwzgórze 233(r)
— schemat ośrodkowej kontroli środowiska we
wnętrznego 234(r)
— termodetektory 274
Pojemność krwi krążącej w określonych stanach
fizjologicznych 336
— minutowa serca 352
— płuc całkowita 398, 399(r)
wdechowa 398
----- zalegająca czynnościowa 398
----- życiowa 399
-------- wdechowa 399
— wiązania, odbiór informacji przez komórkę 29
Pokarm, trawienie 425
Pole(a) korowe 179(r)
— ruchowe i czuciowe w korze mózgu człowieka
182(r)
— widzenia 144
----- oka 154(r)
Połączenia szczelinowe 54
Połączenie między korą móżdżku mózgu i rdzeniem kręgowym 188(r) Połóg 512 Połykanie, fazy
427
— kęsa pokarmu 427(r)
Pompa odźwiernikowa 432
działanie 434(r)
— sodowo-potasowa 61
Poród 509
Postawa ciała, ośrodki kontrolujące 180
Potas 471
Potencjał(y) czuciowy wywoływany 120
— czynnościowy 66
----- mięśnia sercowego 110
----- neuronu ruchowego 70(r)
----- wnętrza neuronu 68(r)
— iglicowy 68
— mikrofonowy 155
— następczy 70
----- hiperpolaryzacyjny 70
----- podepolaryzacyjny 70
— postsynaptyczny hamujący 71, 127
pobudzający 67
— progowy 69
— słuchowy wywoływany 120
— spoczynkowy błony komórkowej 60
538
Skorowidz
Potencjal(y) spoczynkowy komórki mięśnia sercowego 110
— wewnątrzkomórkowy komórki mięśnia komór
serca 112(r)
— wzrokowy wywołany 119,120(r)
— zakończenia synaptycznego nerwowo-mięśniowego 103
Powiązania czynnościowe między korą prawej i lewej półkuli 187(r)
Powstawanie impulsów w aksonach 70
Półkula mózgu prawa, przekrój 184(r)
Prawo Dale'a 72
— energii nerwów Johannesa-Mullera 158
— fizjologii 17
Prążek węchowy boczny 132
----- pośredni 132
----- przyśrodkowy 131
Prążki anizotropowe 89
— izotropowe 90
Pre-limfocyty B 290
— T292
Pre-pro-enkefalina 246 Pre-proopiomelanokortyna 241 Pręciki
139
Prędkość przenoszona depolaryzacji w mięśniu
sercowym i układzie przewodzącym 339(t)
— przepływu krwi średnia 362
Prężność cząsteczek tlenu 45
Procesy fizjologiczne organizmu 18
Proerytroblasty 282
Progesteron 257, 507
Projekcja czuciowa 158 Prolaktyna
238, 512 Promegakarioblasty 290
Promieniowanie trzewno-skórne 178
Propranolol 218 Proprioreceptory
167 Prostacykliny 266, 327
Prostaglandyna 257, 266 Próg
nerkowy 483
— pobudliwość komórek smakowych 168
— pobudzenia dla cząsteczek lotnych związków
132
— słyszenia u człowieka 154
Przeczulica skóry 178
Przekaźnik(i) chemiczne, rola w utrzymaniu homoeostazy 213
— wewnątrzkomórkowe 30
— chemiczne 27
— humoralne 318
Przekwitanie 501
Przemiana materii, czynniki wpływające 420
----- podstawowa 419
Przenoszenie informacji 51
— potencjału wywołanego drażnieniem skóry na
palcach ręki 160(r)
Przepływ antydromowy aksoplazmy 64
— burzliwy i laminarny krwi przez naczynie
krwionośne 362(r)
— krwi 362
----- autoregulacja 389
----- i zużycie tlenu przez narządy i tkanki
409(r)
----- przez mózgowie 392
----------schemat 392(r)
-------- nerki całkowity 479
------------efektywny 479
poszczególne narządy w spoczynku wy
rażony w procentach 353(r)
-------- wątrobę 395
------------obliczony 468
----- regulacja miejscowa 386
----- w aorcie 363(r)
----- w naczyniach włosowatych 371
----- w pniu płucnym 363(r)
----- w zbiorniku żylnym 371
— ortodromowy 64
— osocza przez nerki, efektywny 479
Przestrzeń martwa anatomiczna płuc 400
fizjologiczna 401
— płynów ustrojowych 19
Przetwarzanie informacji w organizmie 52(r)
Przewężenie Ranviera 65
Przewodzenie impulsów dla ruchu 183
droga korowo-rdzeniowa boczna
183
---------------- droga korowo-rdzeniowa przednia
183
----- we włóknach bezrdzennych 82
------------ rdzennych 84
----- z siatkówki 146
— impulsu w obrębie synapsy nerwowo-mięśniowej 103(r)
— w zwojach współczulnych 218
Przewód(y) pokarmowy, wchłanianie węglowoda
nów 456
-------- białek i aminokwasów 460
-------- tłuszczów 456
-------- witamin, soli mineralnych i wody 461
— słuchowe zewnętrzne 151
— ślimakowy 153
Przyjmowanie pokarmów 424
Punkt(y) widzenia najbliższy 135
— zamarzania osocza ludzkiego 21
roztworu 20
Rdzeń kręgowy, odruchy własne 127
----- ośrodki 124
----- pobudliwość ośrodków 125
Reagowanie adekwatne 115
Reakcja(e) adrenergiczne 217
— anafilaktyczne 312
Skorowidz
Reakcja(e) degranulacji w neutrofilach 310
— humoralna 317
Receptor(y) AMPA 78
— błonowe 27
— błony komórkowej, gęstość 57(r)
— cholinergiczne 221, 222
— dopaminergiczny 219
— J414
— kainianowy 78
— muskarynowe 109, 218, 221
— nikotynowy 218, 221
— NMDA 78
— opioidowe 81
— rola w organizmie 210
— skórne czucia eksteroceptywnego 158(r)
— transferrynowe 284
— w błonie śluzowej języka, rozmieszczenie
168(r)
— właściwości 157
— układu ruchu 169
Regulacja oddychania 414(r)
— stężenia wolnych glikokortykoidów krążących
we krwi 243(r)
— w dół 58
----- górę 58
Relaksyna 268
Renina 385, 492
Replikacja łańcuchów DNA 46
Reprezentacje ruchu w korze mózgu 181
Resorpcja kanalikowa 483
Restryktaza 37
Resynteza ATP 416(r)
-----w czasie glikolizy beztlenowej 417(r)
----------- glikozy tlenowej 417(r)
Retikulocyty 282
Rezonans fal akustycznych 152(r), 153
RNA matrycowy 37
— transportujący 37
Rodopsyna 139
— rozpad pod wpływem światła 140(r)
Rozrusznik 113
Rozwieranie szyjki macicy 510(r)
Równanie Nernsta 60 Równowaga
energetyczna 18, 423
— homeostatyczna 213
regulacja 213
— kwasowo-zasadowa osocza 320
Równoważnik tlenu energetyczny 418
Ruch(y) ciała, ośrodki kontrolujące 180
— jelita cienkiego 438(r)
grubego 449(r)
— masowe 448
— perystaltyczne błyskawiczne 438
— prawego oka 150(r)
Rybosomy 37
Rytm bioelektryczny żołądka podstawowy 432
539
Samoregulacja napięcia mięśniowego 100
Sarkomer 90, 91(r)
Schemat blokowy zespołów czynnościowych ośrodkowego układu nerwowego 211(r)
— kaskadowy wewnątrzpochodnego mechaniz
mu aktywacji protrombiny 331(r)
zewnątrzpochodnego mechanizmu aktywa
cji protrombiny 331(r)
— ośrodkowej kontroli pracy serca 377(r)
Schody bębenka 151
— przedsionka 151
— kanalikowa 482, 483
Sekretyna 437, 445, 454, 468
Sen 197
Serce, budowa 334
— czynność bioelektryczna 337
— zjawiska akustyczne 356
Serotonina 78, 143, 266
Sfigmogram 364(r), 365
Siateczka śródplazmatyczna gładka 42
Siatkówka, czynność 139
— połączenia między komórkami nerwowymi
142(r)
— przewodzenie impulsów 148
Siła rozdzielcza oka 143
— załamywania układu optycznego oka 134
Składniki odżywcze pokarmu 425
Skurcz auksotoniczny 97
— głodowe żołądka 430
— izometryczny 95
— izotoniczny 95
— izowulometryczny 348
— maksymalny 97
— mechanizm molekularny 93
— perystaltyczne 431
jelita 437
— pojedynczy mięśnia poprzecznie prążkowane
go 95(r)
— tężcowy mięśnia poprzecznie prążkowanego
96(r)
---- niezupełny 96
-----zupełny 96
Soczewka w czasie akomodacji oka, zmiany krzywizny 135(r) Sok jelitowy 440
— trzustkowy 441
-----kontrola wydzielania 444
-----skład 442(t)
— żołądkowy, kontrola wydzielania 434
w fazie głowowej i żołądkowej
435(r)
- skład 428, 431(t)
Somatoksynina 238
Somatomammotropina 238
Somatostatyna 238, 261, 454
Spektryna 300
540
Skorowidz
Spermatogeneza 259, 499
Spermatogonie 499 Spermatydy
499 Spirogram 400(r) Splot
podśluzowy jelita 439
— warstwy mięśniowej jelita 439
Sprzężenie elektrowydzielnicze 103
— proste 56
— zwrotne między korą mózgu i ośrodkami podkorowymi 186(r)
----- ujemne 56
Stężenie jonów 19
Stopień zużycia tlenu przez tkanki 409 Strefa
podtorebkowa (szpikozależna) węzłów chłonnych
296
— pobudliwości zwiększonej podprogowo 125
— pobudzenia 126
Struktury mózgowia uczestniczące w tworzeniu
pamięci świeżej 204(r)
Strzemiączko 151
Substancja P 194
Sumowanie impulsów 67
----- przestrzenne 67
----- w czasie 68
— skurczów pojedynczych 96
Surfaktant 403
Symport 26
Synapsa(y) 67
— chemiczne 55, 142
— elektryczne 56, 142
— hamujące 72
— nerwowo-mięśniowa 104(r)
— pobudzające 67
Synchronizacja czynności elektrycznej mózgu
117 Synteza acetylocholiny
74(r)
— białka 38—41
— polipeptydów na rybosomach siateczki śródplazmatycznej 39(r)
System 53
Szczelina synaptyczna 73, 103
Szmery patologiczne serca 356
Szpik kostny czerwony 278
----- komórki bariery 279
----- zrąb 279
----- komórki siateczki 279
Szyszynka, hormony 263
Śledziona 297
— budowa 298(r)
Ślina 426
— skład 426(t)
Środowisko biologiczne 17
organizmu wewnętrzne 18
Temperament, klasyfikacja wg Pawiowa 207
Terminacja translacji 38 Termogeneza
drżeniowa 273 Termoregulacja 272
Testosteron 245, 257
— regulacja stężenia we krwi 258(r)
— rola 259
Tkanka(i) centralne hematopoetyczne 277
— dyfuzja gazów 409
— krwiotwórcze, podział 277
— limfoidalne 290
— obwodowe hematopoetyczne 277
Tłuszcze obojętne, biosynteza w komórkach wątrobowych 467(r) Tony serca 356 Torowanie 125
Transferryna 284, 461 Transkortyna 242
Transkrypcja 36
— mRNA, opóźnienie 81
Translacja 38
Transmitter(y) 74
— chemiczne depolaryzujące błonę postsynaptyczną75
— cholinergiczny 221
pobudzające 75
— hamujące 79
— i modulatory, koegzystencja w komórkach ner
wowych i o.u.n. 196(t)
— noradrenergiczny 216
— pobudzające 75
— synaptyczne i ich działanie 77(t)
Transport aktywny 25
— dwutlenku węgla we krwi 407
— gazów między powietrzem pęcherzykowym
i tkankami 406(r)
----- we krwi 405
— przez błonę komórkową 24
— tlenu we krwi 405
Triacyloglicerole 457
Trijodotyronina 249
Triplet 36
Trombocytopoeza 287
— w szpiku kostnym czerwonym 289(r)
Trombocyty 290, 318
Tromboksan 266, 327
Tromboplastyna tkankowa 330
Troponina 90
Trójkąt Einthovena 341(r)
Trypsyna 442
Trypsynogen, aktywacja w dwunastnicy 443(r)
Tworzenie się moczu, regulacja 490
— tlenku azotu w komórkach śródbłonka naczy
niowego 387(r)
Tymocyty 293, 295
Tymozyna 294
Skorowidz
Typ(y) układów nerwowych wg Pawtowa 207
Tyreoglobulina 251 Tyroksyna 249
Ucho środkowe 151
— wewnętrzne 150
Uderzenie koniuszkowe serca 354
Układ(y) 53
— autonomiczny, część przywspółczulna 219
wspólczulna 215
— limbiczny 203
— naczyń krwionośnych 276
— nerwowy autonomiczny 214, 220(r)
— nieswoiste 115
— noradrenergiczny mózgowia 193(r)
— optyczny oka 133
— ośrodkowy nerwowy, zespoły czynnościowe 210
— pozapiramidowy, czynność 185
— przewodzący serca 337(r)
— rąbkowy 203
— rekrutujący tworu siatkowatego 192
— reninowo-angiotensynowy 385
— rozszerzający naczynia współczulny 382
— ruchu, droga impulsów 169
— sarkotubularny 91
— sercowo-naczyniowy 334
---- i oddechowy, schemat 419(r)
---- schemat 335(r)
— siateczkowo-śródbłonkowy 317, 298
— siatkowaty pnia mózgowia 190
----------- wstępujący 191
----------- wstępujący pobudzający 192, 211
----------- zstępujący 191
— swoiste 115
— w organizmie człowieka 49(r)
— zgodności tkankowej główny 294
Unerwienie wzajemnie zwrotne 127
Uniport 26
Urobilinogen w osoczu 325
Wada(y) refrakcji 136, 138(r)
----- oczu, korekcja 136
Wapń 471
— łatwo wymienialny 472
— niewymienialny 472
— zjonizowany, stężenie prawidłowe 471
— zmagazynowany w tkance kostnej 471
Warstwa barwnikowa 143
Wazopresyna 228, 253, 384, 468, 490
— argininowa 230(r)
— znaczenie dla przepływu wieńcowego 359
Wątroba, czynność wewnątrzwydzielnicza 446
zewnątrzwydzielnicza 466
— funkcje 464
541
Wątroba, magazynowanie 467
— regulacja czynności 468
Wchłanianie składników pokarmowych w dwunastnicy 462(r)
— tłuszczów przez komórki nabłonka błony ślu
zowej jelita cienkiego 458(r)
— z dwunastnicy i jelita cienkiego 459(r)
Wdech, zmiany ciśnienia w płucach 397(r)
Wentylacja pęcherzyków płucnych 402
— płuc 397
----- maksymalna dowolna 401
----- minutowa 401
Węzeł(y) chłonne 296
— budowa 296(r)
----- strefa podtorebkowa (szpikozależna) 296
— przedsionkowo-komorowy 113, 337
— zatokowo-przedsionkowy 113, 336
----- zmiany potencjału wewnątrzkomórkowego
Więzadełka Zinna 135
Widzenie barwne 149
— obuoczne 144
— stereoskopowe 148(r), 149
Właściwości receptorów 157
Włókienko mięśniowe 89
Włókno(a) aferentne 86, 158
— eferentne 86
— kiciaste 188(r)
— nerwowe, podział 86(t), 87
węchowe 131
— pnące 189
Woda 469
— całkowita organizmu 19
— i elektrolity, drogi utraty 473
— organizmu całkowita 469
Wodorowęglany 473
Wrzecionko nerwowo-mięśniowe 101(r)
Wskaźnik sercowy 352 Współczynnik
oddechowy 418 Wstępna boczna 157
Wstrząs rdzeniowy 129 Wydalanie 50
— łożyska 511(r)
Wydech, zmiany ciśnienia w płucach 397(r)
Wydzielanie gastryny 436(r)
— hormonów w cyklu miesiączkowym 503(r)
— soku żołądkowego, fazy 436
— śliny 426
Wymiennik przeciwprądowy 487
Wyspy Langerhansa 261
— trzustkowe, hormony 261
Wytrysk nasienia 499
Wytwarzanie energii w komórce 46(r)
— przeciwciał pod wpływem antygenu 314(r)
Wyznaczanie osi elektrycznej serca na podstawie
odprowadzeń dwubiegunowych 347(r)
542
Skorowidz
Wzgórek dolny blaszki pokrywy 157
Wzmacniacze przeciwprądowe 487 Wzór
Arnetha 3O8(r) Wzwód prącia 499
^.Zagęszczenie moczu, czynniki wpływające 489
Zakończenie synaptyczne nerwowo-mięśniowe, budowa 103(r)
Zakręt skroniowy górny 157
— zaśrodkowy kory mózgu 161
Zależności między ośrodkiem głodu, sytości i pragnienia, schemat 202(r)
Zapamiętywanie 115, 204
Zapłodnienie 507
Zapotrzebowanie dzienne na energię 421(t)
Zasada uzyskania echokardiogramu 355(r)
Zastawka krętniczo-kątnicza 438
Zero fizjologiczne 162
Zespół(y) czynnościowe ośrodkowego układu nerwowego 210
Zespół(y) komparatora 211
— kontroli ruchów ciała 211
— koordynacji ruchów ciała 211
— rozrusznika aktywności somatycznej 211
Ziarna wydzielnicze 44
Zmysł wzroku 133
Zwoje spiralne ślimaka 156
Źrenica(e), kontrola wielkości 136 — schemat
ośrodków kontrolujących szerokość 137(r)
Żołądek, gromadzenie pokarmów 428
— motoryka 430
— trawienie 428
— wypełnianie kolejnymi porcjami pokarmu 433(r)
Żółć, skład 446(t)
— udział w trawieniu 446
— wydzielanie 445
Kolejne wydanie podręcznika cieszącego się
dotychczas wielkim powodzeniem zarówno
wśród studentów akademii medycznych
oraz innych uczelni wyższych, jak i uczniów
szkół średnich o tym profilu.
Książka ułatwia przyswojenie i powtórzenie
przed egzaminem podstawowych praw
i procesów fizjologicznych odnoszących się
do czynności organizmu człowieka.
Została napisana jako kompendium
i w tej formie spełnia należycie swoją funkcję.
Książka nagrodzona przez Ministra Zdrowia i
Opieki Społecznej.
ISBN 83-200-2694-6
www.pzwl.pl
9 788320 026948