układ nerwowy.doc
(188 KB) Pobierz
ZALICZENIA I – UKŁAD NERWOWY
BUDOWA I SKŁAD CIAŁA CZŁOWIEKA
Skład elektrolitowy płynu komórkowego (mEq/l):
Na+ 142
K+ 5
Ca2+ 5
Mg2+ 2
Cl- 102
HCO3- 26
PO43- 2
SO42- 1
Pro- 16
Skład elektrolitowy płynu wewnątrzkomórkowego ( mEq/l):
Na+ 10
K+ 155
Ca2+ 0
Mg2+ 30
Cl- 2
HCO3- 8
PO43- 95
SO42- 20
Pro- 55
CENTRALNY I OBWODOWY UKŁAD NERWOWY
Podział układu nerwowego:
1) Podział anatomiczny:
Ośrodkowy układ nerwowy (to mózgowie i rdzeń kręgowy – struktury leżące wewnątrz
czaszki i kanału kręgowego, otoczone i chronione trzema łącznotkankowymi oponami:
miękką, pajęczą i twardą oraz znajdującym się w przestrzeni podpajęczynówkowej
płynem mózgowo-rdzeniowym)
Obwodowy układ nerwowy ( tworzą go skupiska komórek, zwane zwojami nerwowymi,
oraz nerwy czaszkowe i rdzeniowe- skupiska włókien, które docierają do wszystkich
obszarów skóry, mięśni, stawów i narządów wewnętrznych, łącząc receptory i efektory
całego ciała z rdzeniem kręgowym i mózgowiem)
2) Podział funkcjonalny:
Somatyczny układ nerwowy (przetwarza i przewodzi informacje pochodzące z
receptorów całego ciała, a jego efektorami są mięśnie poprzecznie prążkowane
szkieletowe; znajduje się on pod kontrolą kory mózgowej)
Autonomiczny układ nerwowy (reguluje czynności narządów wewnętrznych oraz
metabolizm tkanek i dokonuje tego na drodze odruchowej, automatycznej, niezależnie
od woli, współdziałając z układem dokrewnym; podlega on kontroli ze strony kory
mózgowej)
Funkcje CSN:
1.
Rdzeń kręgowy
(1) informacje czuciowe
(2) wspólna droga końcowa ruchu
(3) ośrodki odruchów i układu autonomicznego
2.
Rdzeń przedłużony
(1) nn. czaszkowe (smak-VII, słuch- VIII, ruchy języka- IX i XII, ruchy klatki piersiowej tj.
brzusznej- X i XI)
(2) skrzyżowanie piramid + integracja z móżdżkiem
(3) jądro olbrzymiokomórkowe tworu siatkowego
(4) regulacja ciśnienia krwi, oddychania i połykania
3.
Most
(1) przekaz do/od móżdżku (konary środkowe)
(2) ruch odwodzenia gałek ocznych + równowaga
(3) czucie z obszaru głowy +żwacze
(4) regulacja oddychania i snu (twór siatkowy)
4.
Śródmózgowie
(1) czynność dowolna mięśni
(2) integracja z móżdżkiem + limbic system+ twór siatkowy
(3) integracja ze słuchem i wzrokiem + ruchy oczy (n. III i IV) + równowaga
5.
Międzymózgowie
Podwzgórze: (1) integracja części autonomicznych
(2) nadzór układu hormonalnego
(3) regulacja rytmów dobowych
(4) regulacja reakcji obronnych
Wzgórze: (1) czucie (+wzrok i słuch) podkorowe
(2) integracja informacji ruchowych
(3) regulacja czujności (nadzór tworu siatkowego)
6.
Kora
(1) czucie somatyczne, widzenie, słyszenie, węch, smak + polecenie ruchu
(2) myślenie, mowa, uczenie się, pamięć, motywacje + planowanie ruchów (=czynności
ruchów)
układ limbiczny mózgu (w tym „stara” kora)
(1) emocje (lęk, gniew, wstręt, radość, smutek, zaskoczenie)
(2) popędy (pokarmowy, seksualny)
(3) kontrola podwzgórza (układ hormonalny i autonomiczny)
(4) uczenie się i zapamiętywanie
układ siatkowaty mózgu
(1) świadomość podstawowa (rdzenna z tworu siatkowatego)
(2) czuwanie i sen
(3) gotowość funkcjonalna – czujność
(4) świadomość wyższa (korowa) – intelekt, emocje, pamięć
BUDOWA I FUKCJE KOMÓRKI NERWOWEJ
Komórki glejowe i ich funkcje
Komórki glejowe nie uczestniczą bezpośrednio w przetwarzaniu informacji i przewodzeniu sygnałów,
pełnią za to wiele funkcji pomocniczych, a ich liczba wielokrotnie przekracza liczbę neuronów. Dzielimy
je na:
Makroglej
-astrocyty (funkcja podporowa, odżywcza, tworzą barierę krew-mózg)
- oligodendrocyty (wytwarzają osłonki wokół nerwów)
Mikroglej ( funkcja fagocytarna, usuwanie uszkodzonych i obumarłych komórek)
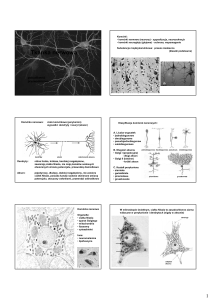
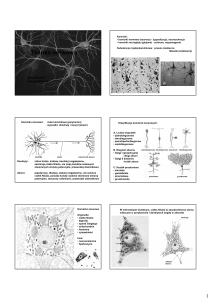
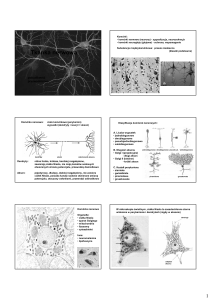
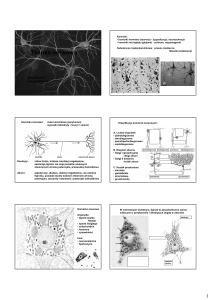
Budowa oraz rola neuronów
Typowy neuron posiada cztery odmienne morfologicznie i funkcjonalnie regiony: ciało komórkowe,
dendryty (w różnej licznie), akson (jeden) wraz z jego zakończeniami presynaptycznymi.
Rodzaje neuronów:
wielobiegunowe (najpowszechniejsze, posiadają wiele rozgałęzionych dendrytów i jeden akson)
pseudobiegunowe (typowe komórki czuciowe, obie wypustki połączone w jeden akson)
dwubiegunowe (dwie wypustki, dendryt odbierający impuls i akson)
jednobiegunowe (bezkręgowce; ma tylko jedną wypustkę pełniącą rolę aksonu i dendrytu)
Podział funkcjonalny neuronów:
aferentne (przewodzą informacje bezpośrednio do receptorów; aksony zmierzają do CSNnerwów czuciowych; ich ciała komórkowe lezą głównie w zwojach rdzeniowych i czaszkowych)
eferentne (ich ciała komórkowe lezą w rdzeniu kręgowym lub pniu mózgu; aksony nerwów
ruchowych biegną do efektorów np. do mięśni poprzecznie prążkowanych)
pośredniczące (najliczniejsza grupa neuronów; przekazują informacje między jednym a drugim
neuronem)
Błonowy potencjał spoczynkowy
Polaryzacją błony komórkowej nazywamy stan, w którym na zewnętrznej powierzchni błony
komórkowej gromadzą się ładunki dodatnie, a po stronie wewnętrznej- ujemne. Polaryzacja powoduje,
że pomiędzy wnętrzem komórki a jej otoczeniem istnieje potencjał elektrycznych, zwana potencjałem
spoczynkowym. Potencjał spoczynkowy neuronu wynosi -70mV. Działające na neuron bodźce mogą
powodować zmniejszenie lub zwiększenie różnicy potencjałów. W pierwszym przypadku będziemy
mówić o depolaryzacji, w drugim o hiperpolaryzacji błony komórkowej. Depolaryzacja zwiększa
pobudliwość neuronu i przyczynia się do powstania impulsu elektrycznego w postaci tzw. potencjału
czynnościowego, natomiast hiperpolaryzacja zmniejsza pobudliwość neuronu (hamuje w danym
momencie jego aktywność). Do utrzymania prze neuron spoczynkowej różnicy potencjałów potrzebne są
mechanizmy, które będą aktywnie reagowały nadmierny wpływ lub ucieczkę jonów z komórki.
Najważniejszym z nich jest pompa sodowo-potasowa, która wbrew gradientowi stężeń usuwa z
neuronu nadmiar jonów Na+ i wprowadza do komórki jony K+.
Potencjał czynnościowy
Potencjał czynnościowy jest świadectwem pobudzenia neuronu. Bodziec (lub suma bodźców) działający
na neuron powoduje ruch jonów przez błonę komórkową, którego efektem jest miejscowa
depolaryzacja. Jeżeli bodziec jest słaby (bodziec podprogowy), to po krótkim czasie następuje powrót do
potencjału spoczynkowego i nie dochodzi do wyzwolenia potencjału iglicowego. Jeżeli zaś bodziec jest
wystarczająco silny (bodziec nadprogowy), to depolaryzacja osiąga potencjał progowy (zwykle 10-2- mV
powyżej potencjału spoczynkowego, czuli ok. -55mV), przy którym dochodzi do szybkiego otwierania się
bramkowych napięciem kanałów sodowych. Powoduje to gwałtowny napływ jonów Na+ do wnętrza i
dalszą depolaryzację. Cechą charakterystyczną potencjału czynnościowego jest fakt, że zawsze pojawia
się zgodnie z prawem „wszystko albo nic”. Bodziec podprogowy jest nieefektywny, natomiast
przekroczenie progu depolaryzacji wyzwala zawsze w danym neuronie taki sam potencjał czynnościowy,
o maksymalnej amplitudzie. Zmiany pobudliwości pojawiają się od momentu przekroczenia potencjału
progowego. W czasie narastania iglicy potencjału i w początkowym okresie repolaryzacji neuron jest
zupełnie niepobudliwy, niewrażliwy na żadne, nawet bardzo silne, bodźce. Jest to okres refrakcji
bezwzględnej. Po nim następuje okres refrakcji względnej, kiedy neuron można pobudzić tylko bodźcem
silniejszym niż w zwykłych warunkach.
WŁÓKNA NERWOWE
Aksony w obwodowym układzie nerwowym łączą się w wiązki, które tworzą nerwy docierające do
wszystkich narządów i części ciała. Dlatego zamiennie stosuje się w stosunku do nich nazwę włókna
nerwowe. W obrębie jednego nerwu przebiegają zarówno włókna przewodzące informacje do
efektorów, jak i przekazujące sygnały pochodzące z receptorów, zwykle towarzyszą im też włókna układu
autonomicznego. Są to aksony o różnej średnicy, nagie (tzn. bez osłonki mielinowej) lub posiadające
osłonki mielinowe. Zaproponowano wiele podziałów włókien nerwowych, z których najczęściej stosuje
się dwa: ze względu na średnicę i szybkość przewodzenia impulsów (3 główne typy włókien- A, B, C) oraz
ze względu na rodzaj unerwionej struktury (dotyczy układu somatycznego i wyróżnia włókna eferentneruchowe- oraz 4 typy włókien aferentnych –czuciowych).
Przewodnictwo ortodromowe i antydromowe
Sygnał w postaci potencjału czynnościowego generowany jest najczęściej we wzgórzu aksonu, a
następnie przewodzony aż do zakończeń presynaptycznych i poprzez synapsy przekazywany na kolejne
neurony lub efektory. W warunkach fizjologicznych przewodzenie odbywa się zawsze w kierunku od
wzgórza aksonu do zakończeń presynaptycznych. Jest to przewodzenie ortodromowe. W warunkach
eksperymentalnych istnieje możliwość ponudzenia aksonu w dowolnym miejscu, co powoduje
generowanie potencjału czynnościowego, który jest następnie przewodzony w obu kierunkach od
miejsca powstania – zarówno w kierunku zakończenia presynaptycznego (ortodromowo), jak i w
kierunku przeciwnym, do ciała komórkowego. Mówimy wtedy o przewodzeniu antydromowym.
Ponieważ impuls nie dociera wtedy do zakończeń aksonu, nie jest przenoszony na inne komórki i ulega
wygaszeniu na ciele komórki lub dendrytach.
Przewodnictwo ciągłe i skokowe
Bardzo ważna funkcjonalnie jest szybkość przewodzenia potencjałów czynnościowych. Zależy ona od
dwóch podstawowych czynników - od grubości aksonu i obecności osłonki mielinowej wokół niego.
Najczęściej istnieje korelacja pomiędzy obiema cechami: aksony cienkie na ogół pozbawione są osłonki
mielinowej, mają ją natomiast aksony grube. W aksonach nagich, niezmielinizowanych, występuje
przewodnictwo ciągłe, które polega na stopniowym przesuwaniu się wzdłuż aksonu fali depolaryzacji.
Przewodnictwo ciągłe u wszystkich kręgowców jest stosunkowo wolne, zwykle w granicach 0,5-2 m/s, a
prędkość przewodzenia rośnie proporcjonalnie do średnicy aksonu. Obecność osłonki mielinowej
zwiększa prędkość przewodzenia od kilkunastu do kilkudziesięciu razy - w najgrubszych włóknach
mielinowych wynosi nawet 120 m/s. Osłonka mielinowa izoluje akson od płynu pozakomórkowego i
zwiększa dystans pomiędzy kolejnymi punktami, które osiągają próg depolaryzacji. Mamy tu do
czynienia z przewodnictwem skokowym, gdyż dochodzi do pozornego skoku potencjału wzdłuż błony
komórkowej pokrytej osłonką, na odcinku pomiędzy kolejnymi przewężeniami, w których występuje
przewodnictwo ciągłe.
TRANSMISJA SYNAPTYCZNA
Budowa i rodzaje synaps
1.
Synapsy chemiczne
Komunikacja pomiędzy neuronami lub pomiędzy neuronem a efektorem odbywa się tylko w
wyspecjalizowanych miejscach, które nazywamy synapsami. Najpowszechniejsze w układzie
nerwowym ssaków są synapsy chemiczne, w których nie dochodzi do bezpośredniego kontaktu
pomiędzy błonami komórkowymi neuronów, lecz oddziela je wąska szczelina. Pozwala to
wyróżnić trzy części takiego połączenia: część presynaptyczną, szczelinę synaptyczną i część
postsynaptyczną. Sygnał przekazywany jest z jednej komórki nerwowej na drugą za
pośrednictwem cząsteczek chemicznych (neurotransmiterów). Zakończenie presynaptyczne
aksonu ma zazwyczaj kształt kolbki, w której znajdują się liczne pęcherzyki synaptyczne,
wypełnione cząsteczkami neurotransmitera (zwykle jeden pęcherzyk zawiera 5-10 tysięcy
cząsteczek przekaźnika). Błona komórkowa pokrywająca kolbkę od strony szczeliny synaptycznej
nazywa się błoną presynaptyczną. Wyróżnia się w niej strefy aktywne, w których rozmieszczone
są kanały jonowe dla Ca2 + , bramkowane napięciem.
2.
Synapsy elektryczne
Synapsy elektryczne nazywa się także połączeniami szczelinowymi. Neurony są od siebie
oddalone zaledwie o ok. 3,5 nm i nie ma pomiędzy nimi właściwej szczeliny synaptycznej, gdyż
są ze sobą połączone specjalnymi strukturami (kanałami) białkowymi, które umożliwiają
bezpośredni przepływ prądu jonowego pomiędzy komórką presynaptyczną a postsynaptyczną.
Nie ma pośrednictwa przekaźnika chemicznego. Dzięki łączności pomiędzy cytoplazmą jednej i
drugiej komórki nawet podprogowa depolaryzacja błony presynaptycznej powoduje przepływ
prądu i depolaryzację błony postsynaptycznej, a po osiągnięciu potencjału progowego wyzwala,
zgodnie z prawem „wszystko albo nic", potencjał czynnościowy. Impulsy elektryczne pomiędzy
neuronami przekazywane są w tym przypadku praktycznie bez opóźnienia synaptycznego.
Ponadto, potencjał czynnościowy może być wywołany jednocześnie we wszystkich połączonych
synapsami elektrycznymi neuronach, co ułatwia synchroniczne pobudzenie całej grupy komórek.
Trzecia cecha połączeń szczelinowych polega na możliwości dwukierunkowego przekazywania
sygnału, co nigdy nie zachodzi w synapsach chemicznych. Specyfika połączeń, stosunkowo duży
rozmiar i brak selektywności kanałów dają możliwość nie tylko komunikacji elektrycznej, ale i
przekazywania pomiędzy komórkami niektórych jonów oraz małych cząsteczek organicznych.
Zjawiska EPSP i IPSP
Lokalna depolaryzacja nazywa się postsynaptycznym potencjałem pobudzającym - EPSP, natomiast
hiperpolaryzacja określana jest jako postsynaptyczny potencjał hamujący - IPSP. Im więcej cząsteczek
neurotransmitera zostanie uwolnionych z zakończenia aksonu do szczeliny synaptycznej, tym więcej
receptorów postsynaptycznych jest aktywowanych i w efekcie rośnie amplituda EPSP lub IPSP. Każdy
potencjał czynnościowy (który ma zawsze maksymalną amplitudę) powoduje egzocytozę określonej
liczby pęcherzyków synaptycznych. Istnieją jednak mechanizmy, które umożliwiają zwiększenie lub
zmniejszenie ilości uwalnianego neurotransmitera. Jest to torowanie lub hamowanie presynaptyczne, a
kluczową rolę odgrywają synapsy aksono-aksonalne. Przekaźnik chemiczny uwalniany w takiej synapsie
oddziałuje na receptory zakończenia presynaptycznego drugiego aksonu i w rezultacie zwiększa
(torowanie) lub zmniejsza (hamowanie) napływ do jego kolbki synaptycznej jonów Ca2+. Jony
wapniowe, jak wspomniano wyżej, regulują uwalnianie neurotransmitera, co pośrednio doprowadza do
zwiększenia lub zmniejszenia amplitudy powstającego potencjału postsynaptycznego. W ten sposób inny
neuron, poprzez synapsę aksono-aksonalną, w bardzo precyzyjny sposób moduluje czynność nie całej
komórki nerwowej, ale tylko jednej z wielu tworzonych przez nią synaps. Procesy torowania i
hamowania presynaptycznego występują często w zakończeniach aksonów komórek czuciowych w
rdzeniu kręgowym.
Dywergencja i konwergencja
Do komórki nerwowej danego typu dochodzą więc informacje z wielu źródeł. Taki wzorzec połączeń
nazywa się konwergencją. Dzięki konwergencji neuron może integrować dochodzące do niego w tym
samym czasie różne sygnały i po dokonaniu syntezy przekazać do efektora odpowiednią informację w
postaci kodu nerwowego. Akson każdego neuronu rozgałęzia się i tworzy ...
Plik z chomika:
kiwibla
Inne pliki z tego folderu:
Drogi piramidowe i pozapiramidowe.doc (677 KB)
Budowa i sk_ad cia_a cz_owieka.docx (16 KB)
Centralny i obwodowy uk_ad nerwowy.docx (30 KB)
układ nerwowy.doc (188 KB)
Inne foldery tego chomika:
Zgłoś jeśli naruszono regulamin
Strona główna
Aktualności
Kontakt
Dla Mediów
Dział Pomocy
Opinie
Program partnerski
Regulamin serwisu
Polityka prywatności
Ochrona praw autorskich
Platforma wydawców
Copyright © 2012 Chomikuj.pl
3. Układ krwionośny
egzamin
Fizjologia wysiłku
oddechowy