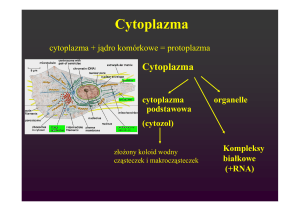
5. Cytoszkielet
Kształt
Transportowanie i rozmieszczenie organelli i makromolekuł
Wewnętrzna organizacja
Ruch
Skurcz
Podział
Fagocytoza
Zjawisko adhezji komórkowej
Mikrotubule (25nm)
Polaryzacja mikrotubul związana jest z hydrolizą GTP i przyłączaniem α i β tubuliny. Koniec
+ ma większe powinowactwo do GTP, niż koniec -, dlatego polaryzacja MT jest wyraźnie
uprzywilejowana na końcu +.
MT zakotwiczone są w Microtubule Organisation Center – głównym jego składnikiem jest γ
tubulina; występuje w kinetochorach, ciałkach podstawowych rzęsek i wici.
Centrosom zwierzęcy – 2 centriole ustawione pod kątem 90o
Interfaza
Preprofaza
Podział
Cytokineza
Komórka zwierzęca
Komórka roślinna
1 MTOC
Sieć kortykalna (MTOC b. liczne
– przy jądrze, plastydach, błonie...)
Pierścień PPB
Budują wrzeciono
Budują wrzeciono
Fragmoplast
Białka Microtubule Associated Proteins:
MAP stabilizujące - łączące sie z bocznymi powierzchniami mikrotubul w miejscu
acetylacji lub fosforylacji tubuliny α
Białka tau – w większości komórek, w aksonach łączy mikrotubule w
pęczki. Jego brak zaburza tworzenie aksonów.
MAP2 – utrzymują uporządkowaną strukturę MT w dendrytach i
aksonach.
Kateniny – łączą MT z innymi elementami cytoszkieletu
MAP łączące koniec + z innymi strukturami
Białka oczapkowujące – białka kompleksu Tip Attachement Complex
(TAP); przyczepiają MT do ER.
MAP umożliwiające ruch na MT; białka motoryczne
Kinezyna – zbudowane z 2 łańcuchów ciężkich (zdolność hydrolizy
ATP) i lekkich (zdolność przyłączania ładunku). Transport
od – do +; sekrecja. Razem z dyneiną bierze udział w
segregacji chromosomów.
Dyneina – zbudowana z łańcuchów ciężkich, pośrednich i lekkich.
Transport od + do -; wchłanianie, zakotwiczają Ap.
Golgiego; motor molekularny rzęsek i wici.
Dynamina – Kieruje aktywnym ruchem ślizgowym jednej MT po
drugiej; zaangażowana w endocytozę.
KAR3 i NCD
1
W komórce roślinnej:
MAP65 – łączy 2 równolegle nachodzące na siebie MT
MAP190 – występują we wrzecionie kariokinetycznym
Związki zaburzające organizacje MT:
Kolchicyna – wiąże się z wolnymi heterodimerami α tubuliny i uniemożliwia
powstanie MT; przeważają procesy depolimeryzacji.
Winkrystyna i Winblastyna – strącają MT
Taksol – stabilizuje MT; wiąże MT uniemożliwiając ich rozpad, ale nie przeszkadza w
polimeryzacji.
D2O – Stabilizuje MT
Mikrofilamenty (filamenty Aktynowe 5-8 nm)
Filamenty aktynowe, zbudowane z monomerów aktyny G, stanowią do 20% wszystkich
białek w komórce eukariotycznej. Są zlokalizowane jedynie pod błoną plazmatyczną i
stanowią część korową cytoplazmy określając jej kształt i mechaniczne właściwości.
Polimeryzacja aktyny wymaga:
ATP, które przy każdorazowym dołączeniu podjednostki do filamentu jest
hydrolizowane do ADP
Jonów K+ i Mg2+
Koniec + (lotkowy) filamentu polimeryzuje 10 razy szybciej niż koniec – (grotowy)
Filamenty aktynowe tworzą różne formy organizacji:
Mikrokosmki
Włókna naprężeniowe – Jeden koniec tych włókien umieszczony w płytce
przylegania, w której komórka jest przytwierdzona do
macierzy zewnątrzkomórkowej. Drugi koniec może być
przyłączony do innej płytki przylegania bądź, do filamentów
pośrednich otaczających jądro.
Lamellipodia – wypustki wysuwane z powierzchni komórki będące gęsta siecią
filamentów aktynowych. Filamenty aktynowe mają swoje końce +
skierowane do krawędzi błony komórkowej. Marszczenie błony
komórkowej występuje wtedy, gdy lamellipodium nie może
przymocować się do podłoża.
Filopodia – stożek wzrostowy rozwijającego się aksonu komórki nerwowej.
Pierścień kurczliwy – złożony z filamentów aktyny i miozyny, rozdziela 2 komórki
siostrzane w trakcie cytokinezy (zależny od [Ca2+]).
Actin Binding Proteins – białka wiążące aktynę:
Tropomiozyna – białka przełączające kontrolujące skurcz mięśnia.
Profilina – rodzina białek wiążących monomery aktynowe; często związana z błoną,
wiąże się w stosunku 1:1, reguluje poziom aktyny włókienkowatej.
Tymozyna β4 – potencjalny inhibitor polimeryzacji aktyny. W niektórych komórkach
(np. płytki krwi człowieka) całkowicie blokuje polimeryzacje
monomerycznej aktyny poprzez wiązanie się z nią.
Spektryna – związana z błoną erytrocytów i stanowi grotowy koniec białka wiążącego.
W tych komórkach błona jest podtrzymywana przez sieć tetramerów
spektryny.
Fibryna – łączą filamenty aktynowe w równoległe szeregi (pęczki).
2
α-aktynina – wiąże filamenty, pomaga zakotwiczyć końce włókienek naprężeniowych
w obszarze błony zwanym płytką przylegania, gdzie komórka łączy się z
macierzą zewnątrzkomórkową.
Filamina – białko tworzące żel, które przyczynia się do tworzenia gęstej, lepkiej sieci
przez sieciowanie filamentów aktynowych.
Związki zaburzające polimeryzację filamentów aktynowych:
Cytochalazyny – wydzielane przez śluzowce wiążą się z końcem + filamentu aktynowego,
nowe monomery nie mogą być dołączane – w efekcie depolimeryzacja.
Falloidyna – wydzielana przez muchomor sromotnikowy. Stabilizuje filamenty aktynowe
hamując depolimeryzację. Zjedzenie surowego mięsa neutralizuje.
Aktyna jest zaangażowana w fagocytozę.
Filamenty pośrednie (10nm)
Trwała, włóknista struktura, odporna na rozciąganie – nadaje komórce wytrzymałości.
Brak polaryzacji
Występują w jądrze komórkowym tworząc laminę jądrową otoczki jądrowej.
Pełnią funkcje podporowe
Struktura filamentów pośrednich:
Monomery →α-helikalnie zwinięte dimery →tetramer (2 dimery zwinięte
antyrównolegle)→protofilament (7 lub 8 splecionych kompleksów tetramerycznych).
Wytrzymałość na rozciąganie zależy w dużej mierze od naprzemiennego ułożenia dimerów
każdego tetrameru.
Białka filamentów pośrednich:
Keratyny – u człowieka, co najmniej 20 różnych. Występują w komórkach nabłonka,
skóry. Ciężkie keratyny są swoiste dla włosów i paznokci. Tworzy
heteropolimery.
Wimentyna i białka pokrewne – najczęściej występujące filamenty pośrednie. Obecne
w fibroblastach i komórkach śródbłonka (tkanka
łączna). W odróżnieniu od keratyn mogą tworzyć
polimery z jednego rodzaju białka, homopolimery.
Białka NeuroFilamentów – rozciągają się wzdłuż neuronu. Nadają długim wypustkom
komórek nerwowych odporność na rozciąganie. Odgrywają
istotną rolę w mocowaniu komórek nabłonkowych do:
1. Połączeń komórkowych nazywanych desmosomami,
kotwiczących przylegające komórki.
2. Hemidesmosomów, które wiążą komórki do blaszki
podstawnej.
3
Laminy jądrowe – 3 rodzaje A, B i C. Lamina jądrowa lub lamina włóknista składa się
z sieci filamentów pośrednich na wewnętrznej powierzchni błony
jądrowej i jest związana z porami jądrowymi. W odróżnieniu od
innych filamentów laminy są demontowane (fosforylacja reszt
serynowych umożliwia rozpuszczanie) podczas mitozy.
Typ III – zawierające wimentynę, desminę, białko tk. glejowej,
peryferynę.
Typ IV – w neurofilamentach
Typ V – filamenty jądrowe/laminy
Typ VI filamenty nestyny (w rozwijajacych sieneuronach)
4